Evaluating Biodiversity Metric Response to Forecasted Land Use Change in the Northern Rio Grande Basin

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Datasets
2.3. Analysis
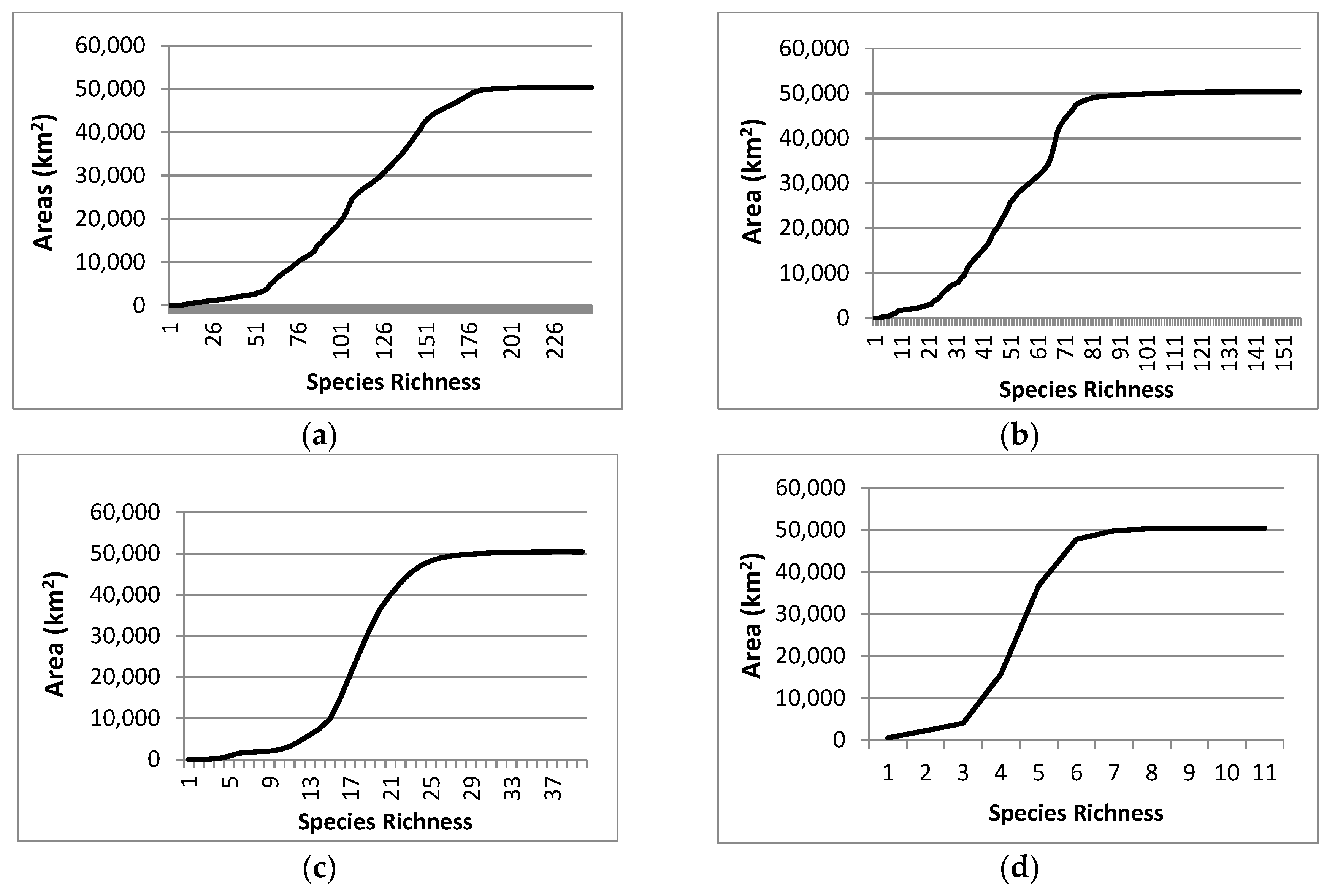
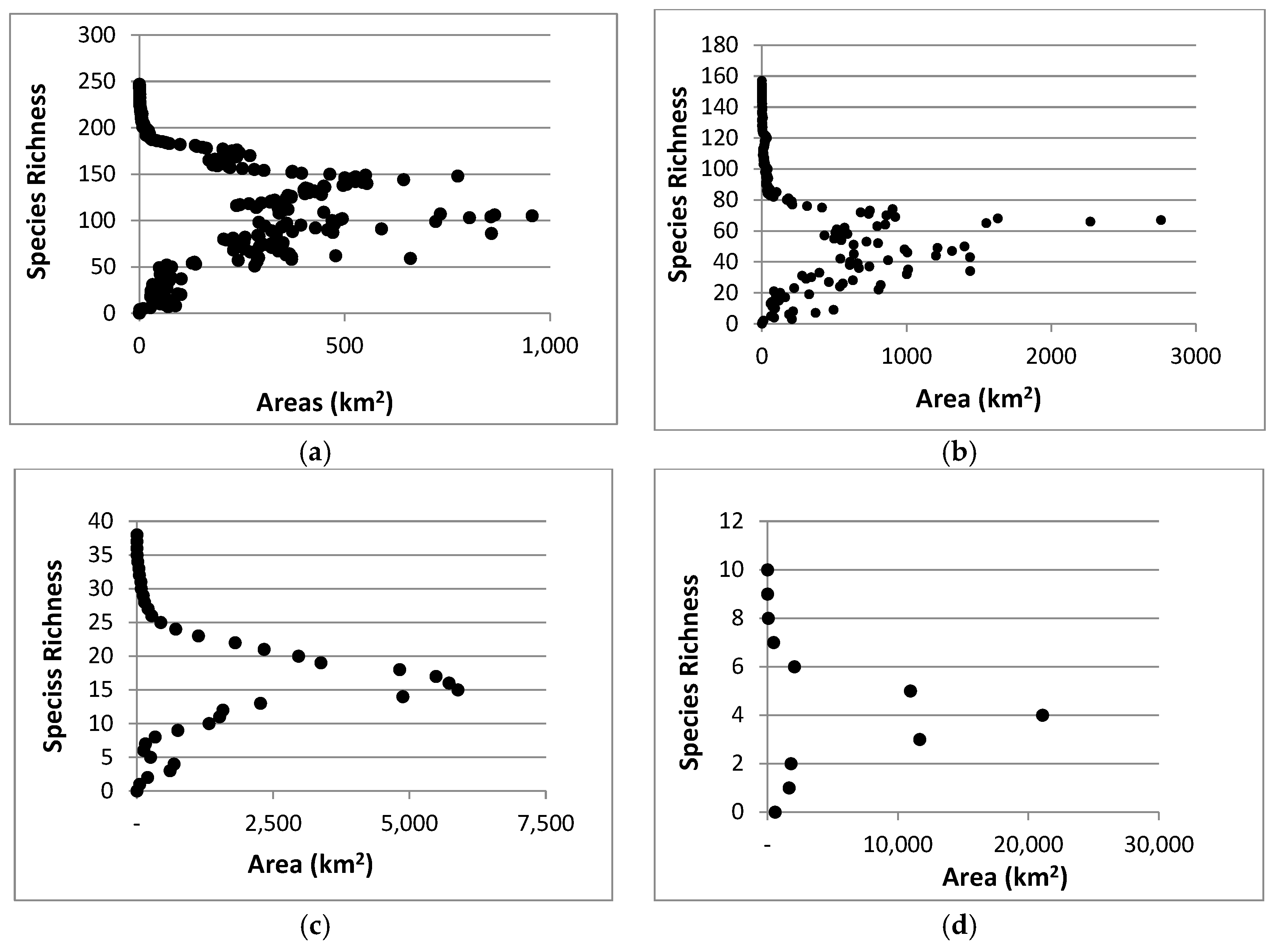
3. Results
3.1. Natural Areas
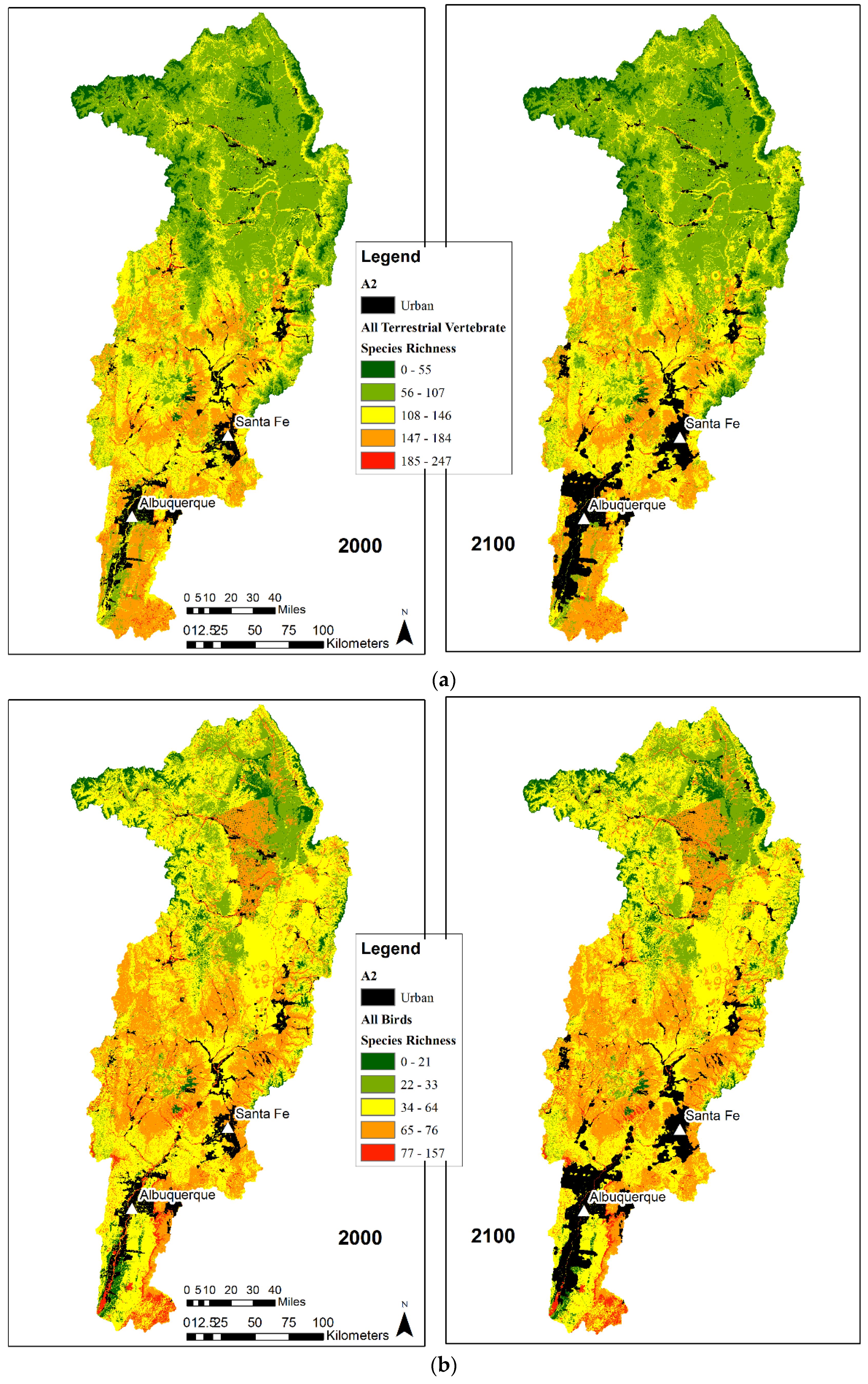
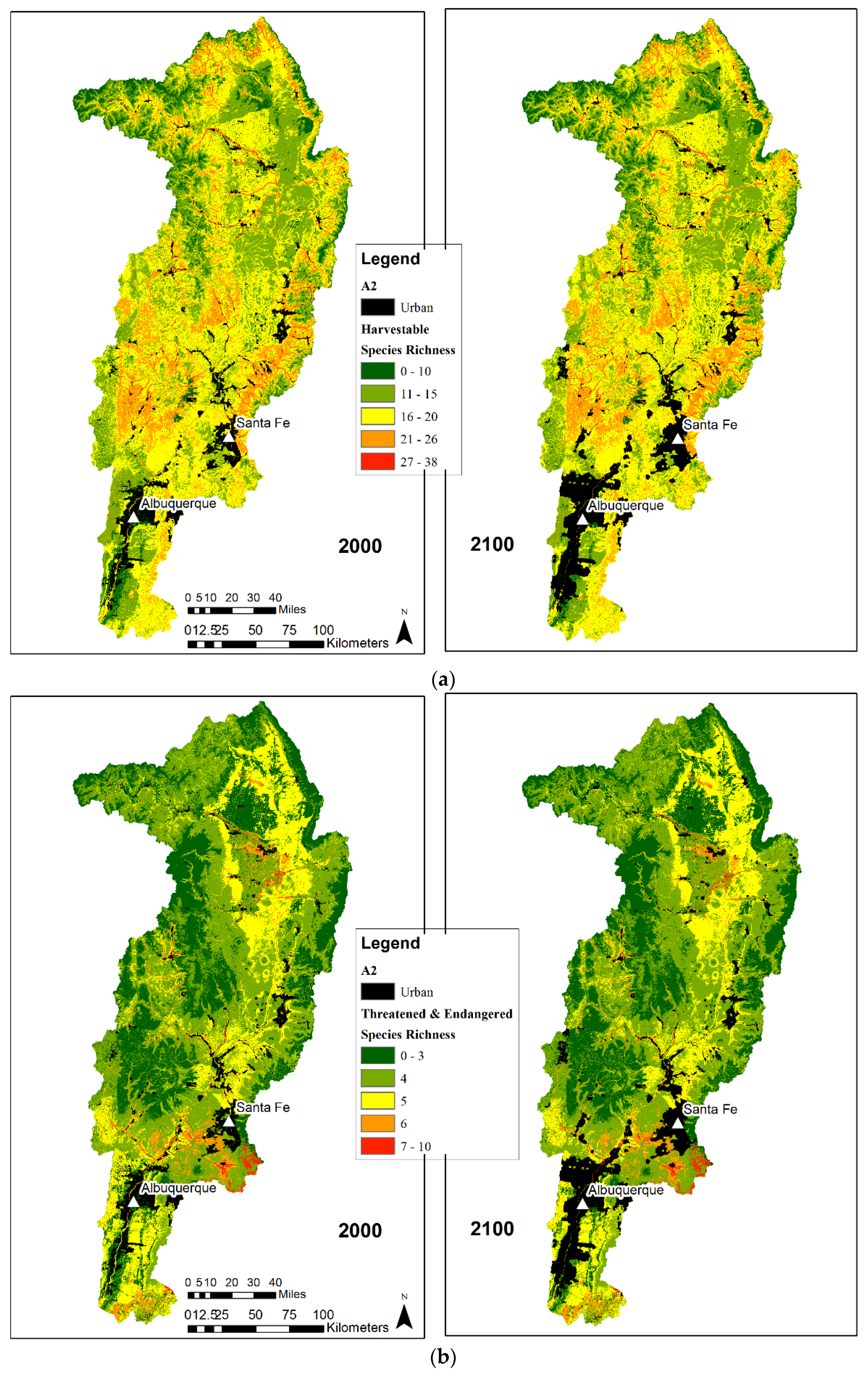
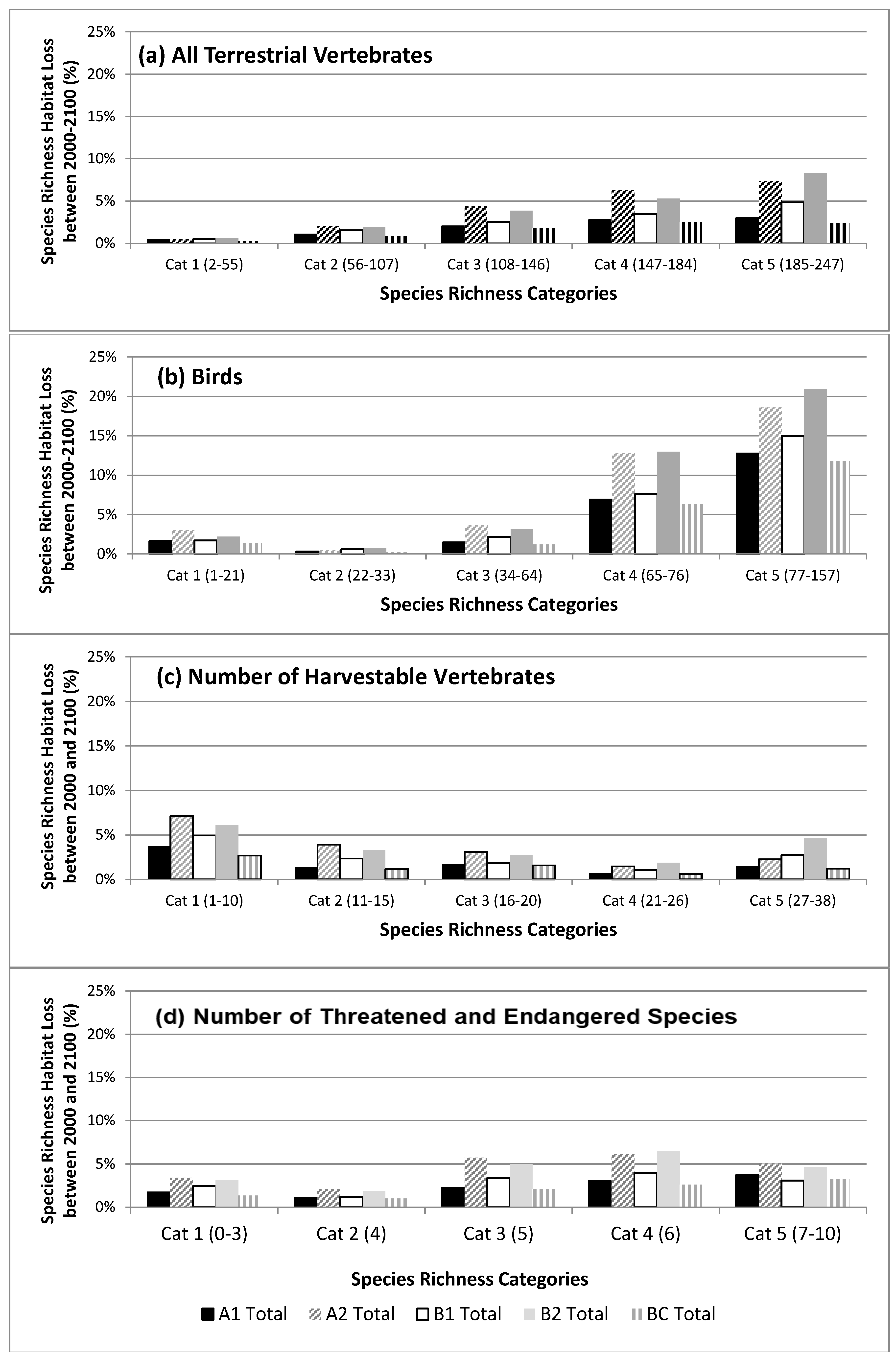
3.2. Species Richness
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mattison, E.H.A.; Norris, K. Bridging the gaps between agricultural policy, land-use and biodiversity. Trends Ecol. Evol. 2005, 20, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Crossman, N.D.; Bryan, B.A.; de Groot, R.S.; Lin, Y.-P.; Minang, P.A. Land science contributions to ecosystem services. Curr. Opin. Environ. Sustain. 2013, 5, 509–514. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment. Ecosystem and Human Well-Being: Biodiversity Synthesis; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Tilman, D. Causes, consequences and ethics of biodiversity. Nature 2000, 405, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Boykin, K.G.; Kepner, W.G.; Bradford, D.F.; Guy, R.K.; Kopp, D.A.; Leimer, A.K.; Samson, E.A.; East, N.F.; Neale, A.C.; Gergely, K.J. A national approach for mapping and quantifying habitat-based biodiversity metrics across multiple spatial scales. Ecol. Indic. 2013, 33, 139–147. [Google Scholar] [CrossRef]
- McKee, J.K.; Sciulli, P.W.; Fooce, C.D.; Waite, T.A. Forecasting global biodiversity threats associated with human population growth. Biol. Conserv. 2004, 115, 161–164. [Google Scholar] [CrossRef]
- Hutton, J.M.; Leader-Williams, N. Sustainable use and incentive-driven conservation: Realigning human and conservation interests. Oryx 2003, 37. [Google Scholar] [CrossRef]
- Corvalán, C.F.; Hales, S.; McMichael, A.J.; Organization, W.H. Ecosystems and Human Well-Being: Health Synthesis: A Report of the Millennium Ecosystem Assessment; World Health Organization: Geneva, Switzerland, 2005; pp. 12–37. [Google Scholar]
- Franklin, J.F. Preserving biodiversity: Species, ecosystems, or landscapes? Ecol. Appl. 1993, 3, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.A. Ecosystems and the biosphere as complex adaptive systems. Ecosystems 1998, 1, 431–436. [Google Scholar] [CrossRef]
- Braat, L.C.; de Groot, R. The ecosystem services agenda: Bridging the worlds of natural science and economics, conservation and development, and public and private policy. Ecosyst. Serv. 2012, 1, 4–15. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.-S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services: Biodiversity and ecosystem functioning/services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Sala, O.E. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Swetnam, R.D.; Fisher, B.; Mbilinyi, B.P.; Munishi, P.K.T.; Willcock, S.; Ricketts, T.; Mwakalila, S.; Balmford, A.; Burgess, N.D.; Marshall, A.R.; et al. Mapping socio-economic scenarios of land cover change: A GIS method to enable ecosystem service modelling. J. Environ. Manag. 2011, 92, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ouyang, Z.; Taylor, W.W.; Groop, R.; Tan, Y.; Zhang, H. A framework for evaluating the effects of human factors on wildlife habitat: The case of giant pandas. Conserv. Biol. 1999, 13, 1360–1370. [Google Scholar] [CrossRef]
- Lawler, J.J.; Lewis, D.J.; Nelson, E.; Plantinga, A.J.; Polasky, S.; Withey, J.C.; Helmers, D.P.; Martinuzzi, S.; Pennington, D.; Radeloff, V.C. Projected land-use change impacts on ecosystem services in the United States. Proc. Natl. Acad. Sci. USA 2014, 111, 7492–7497. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, M.L.; Norman, L.M.; Boykin, K.G.; Wallace, C.S.A. Biodiversity losses and conservation trade-offs: Assessing future urban growth scenarios for a North American trade corridor. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2013, 9, 90–103. [Google Scholar] [CrossRef]
- Samson, E.A.; Kepner, W.G.; Boykin, K.G.; Bradford, D.F.; Bierwagen, B.G.; Leimer, A.K.K.; Guy, R.K. Evaluating biodiversity response to a forecasted land use change: A case study in the South Platte River Basin, Colorado. In Observing, Studying and Managing for Change—Proceedings for the Fourth Interagency Conference on Research in the Watersheds; USGS Scientific Investigations Report; USGS: Reston, VA, USA, 2011; Volume 5169, pp. 56–62. [Google Scholar]
- Larigauderie, A.; Prieur-Richard, A.-H.; Mace, G.M.; Lonsdale, M.; Mooney, H.A.; Brussaard, L.; Cooper, D.; Cramer, W.; Daszak, P.; Díaz, S.; et al. Biodiversity and ecosystem services science for a sustainable planet: The DIVERSITAS vision for 2012–2020. Curr. Opin. Environ. Sustain. 2012, 4, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. Biodiversity and resilience of ecosystem functions. Trends Ecol. Evolut. 2015, 30, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C. Managing ecosystem services: What do we need to know about their ecology? Ecol. Lett. 2005, 8, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.C.; Matusik-Rowan, P.L.; Boykin, K.G. Prioritizing conservation potential of arid-land montane natural springs and associated riparian areas. J. Arid Environ. 2002, 50, 527–547. [Google Scholar] [CrossRef]
- Skagen, S.K.; Kelly, J.F.; van Riper, C.; Hutto, R.L.; Finch, D.M.; Krueper, D.J.; Melcher, C.P. Geography of spring landbird migration through riparian habitats in Southwestern North America. Condor 2005, 107, 212. [Google Scholar] [CrossRef]
- Szaro, R.C. Riparian Forest and Scrubland Community Types of Arizona and New Mexico. Desert Plants (USA) 1989, 9, 69–138. [Google Scholar]
- Naiman, R.J.; Decamps, H.; Pollock, M. The role of riparian corridors in maintaining regional biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.A.; Stromberg, J.C.; Goodrich, D.C.; Dixon, M.D.; Lansey, K.; Kang, D.; Brookshire, D.S.; Cerasale, D.J. Projecting avian response to linked changes in groundwater and riparian floodplain vegetation along a dryland river: A scenario analysis. Ecohydrology 2011, 4, 130–142. [Google Scholar] [CrossRef]
- Brand, L.A.; White, G.C.; Noon, B.R. Factors influencing species richness and community composition of breeding birds in a desert riparian corridor. Condor 2008, 110, 199–210. [Google Scholar] [CrossRef]
- Brand, L.A.; Dixon, M.D.; Fetz, T.; Stromberg, J.C.; Stewart, S.; Garber, G.; Goodrich, D.C.; Brookshire, D.S.; Broadbent, C.D.; Benedict, K. Projecting avian responses to landscape management along the Middle Rio Grande, New Mexico. Southwest. Nat. 2013, 58, 150–162. [Google Scholar] [CrossRef]
- Steinitz, C.; Arias, H.; Bassett, H.; Flaxman, M.; Goode, T.; Maddock, T.; Mouat, D.; Peiser, R.; Shearer, A. Alternative Futures for Changing Landscapes: The Upper San Pedro River Basin in Arizona and Sonora; Island Press: Washington, DC, USA, 2003; ISBN 978-1-55963-224-9. [Google Scholar]
- Fernald, A.G.; Baker, T.T.; Guldan, S.J. Hydrologic, riparian, and agroecosystem functions of traditional acequia irrigation systems. J. Sustain. Agric. 2007, 30, 147–171. [Google Scholar] [CrossRef]
- Fernald, A.; Tidwell, V.; Rivera, J.; Rodríguez, S.; Guldan, S.; Steele, C.; Ochoa, C.; Hurd, B.; Ortiz, M.; Boykin, K.; et al. Modeling sustainability of water, environment, livelihood, and culture in traditional irrigation communities and their linked watersheds. Sustainability 2012, 4, 2998–3022. [Google Scholar] [CrossRef]
- Fernald, A.; Guldan, S.; Boykin, K.; Ochoa, C.; Al, E. Linked hydrologic and social systems that support resilience of traditional irrigation communities. Hydrol. Earth Syst. Sci. 2015, 19, 293–307. [Google Scholar] [CrossRef]
- Turner, B.; Tidwell, V.; Fernald, A.; Rivera, J.; Rodriguez, S.; Guldan, S.; Ochoa, C.; Hurd, B.; Boykin, K.; Cibils, A. Modeling acequia irrigation systems using system dynamics: Model development, evaluation, and sensitivity analyses to investigate effects of socio-economic and biophysical feedbacks. Sustainability 2016, 8, 1019. [Google Scholar] [CrossRef]
- Bierwagen, B.G.; Theobald, D.M.; Pyke, C.R.; Choate, A.; Groth, P.; Thomas, J.V.; Morefield, P. National housing and impervious surface scenarios for integrated climate impact assessments. Proc. Natl. Acad. Sci. USA 2010, 107, 20887–20892. [Google Scholar] [CrossRef] [PubMed]
- Boykin, K.G.; Thompson, B.C.; Deitner, R.A.; Schrupp, D.; Bradford, D.; O’Brien, L.; Drost, C.; Propeck-Gray, S.; Rieth, W.; Thomas, K.; et al. Predicted animal habitat distributions and species richness. In Southwest Regional Gap Analysis Final Report; US Geological Survey, Gap Analysis Program: Moscow, ID, USA, 2007. [Google Scholar]
- U.S. Census Bureau. American FactFinder—Results; U.S. Census Bureau: Suitland, MD, USA, 2015.
- U.S. Census Bureau. Interim State Population Projections. 2005. Available online: http://www.census.gov/population/projections/data/state/projectionsagesex.html (accessed on 2 September 2016).
- Nakicenovic, N.; Alcamo, J.; Grubler, A.; Riahi, K.; Roehrl, R.A.; Rogner, H.-H.; Victor, N. Special Report on Emissions Scenarios (SRES), A Special Report of Working Group III of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2000; ISBN 0-521-80493-0. [Google Scholar]
- Prior-Magee, J.S.; Boykin, K.G.; Bradford, D.F.; Kepner, W.G.; Lowry, J.H.; Schrupp, D.L.; Thomas, K.A.; Thompson, B.C. Ecoregional Gap Analysis of the Southwestern United States: The Southwest Regional Gap Analysis Project Final Report; U.S. Geological Survey, Gap Analysis Program: Moscow, ID, USA, 2007.
- US Environmental Protection Agency. Land-Use Scenarios: National-Scale Housing-Density Scenarios Consistent with Climate Change Storylines (Final Report); US Environmental Protection Agency: Washington, DC, USA, 2009.
- Boykin, K.G.; Thompson, B.C.; Propeck-Gray, S. Accuracy of gap analysis habitat models in predicting physical features for wildlife-habitat associations in the southwest US. Ecol. Model. 2010, 221, 2769–2775. [Google Scholar] [CrossRef]
- Wenny, D.G.; Devault, T.L.; Johnson, M.D.; Kelly, D.; Sekercioglu, C.H.; Tomback, D.F.; Whelan, C.J. The need to quantify ecosystem services provided by birds. Auk 2011, 128, 1–14. [Google Scholar] [CrossRef]
- United States Congress. Endangered Species Act; United States Congress: Washington, DC, USA, 1973.
- New Mexico Department of Game and Fish. Comprehensive Wildlife Conservation Strategy for New Mexico; New Mexico Department of Game and Fish: Santa Fe, NM, USA, 2006; p. 526.
- Arizona Game and Fish Department. Arizona’s State Wildlife Action Plan: 2012–2022; Arizona Game and Fish Department: Phoenix, AZ, USA, 2012; p. 233.
- Ochoa, C.G.; Guldan, S.J.; Cibils, A.F.; Lopez, S.C.; Boykin, K.G.; Tidwell, V.C.; Fernald, A.G. Hydrologic connectivity of head waters and floodplains in a semi-arid watershed. J. Contemp. Water Res. Educ. 2013, 152, 69–78. [Google Scholar] [CrossRef]
- Hohensee, P.; Samuel, M.; Drake, D. For what it’s worth: Appreciating the economic value of wildlife. Wildl. Prof. 2017, 11, 40–43. [Google Scholar]
- U.S. Fish and Wildlife Service; U.S. Census Bureau. 2011 National Survey of Fishing, Hunting, and Wildlife-Associated Recreation; U.S. Fish and Wildlife Service: Washington, DC, USA; U.S. Census Bureau: Suitland, MD, USA, 2011; p. 172.
- Cartron, J.L.E.; Stoleson, S.H.; Stoleson, P.L.; Shaw, D.W. Riparian areas. In Livestock Management in the American Southwest: Ecology, Society and Economics; Elsevier: Amsterdam, The Netherlands, 2000; pp. 281–328. [Google Scholar]
- Hubbard, J.P. Importance of riparian ecosystems: Biotic considerations. In Importance, Preservation, and Management of Riparian Habitat: A Symposium (RR Johnson and DA Jones, Tech. Coords.); RM-43; USDA Forest Service: Washington, DC, USA, 1977; pp. 14–18. [Google Scholar]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western U.S. Forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Mccarthy, P.D. Climate change adaptation for people and nature: A case study from the U.S. Southwest. Adv. Clim. Chang. Res. 2012, 3, 22–37. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the Western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Calkins, M.T.; Beever, E.A.; Boykin, K.G.; Frey, J.K.; Andersen, M.C. Not-so-splendid isolation: Modeling climate-mediated range collapse of a montane mammal Ochotona princeps across numerous ecoregions. Ecography 2012, 35, 780–791. [Google Scholar] [CrossRef]
- Lowry, J.; Ramsey, R.D.; Thomas, K.; Schrupp, D.; Sajwaj, T.; Kirby, J.; Waller, E.; Schrader, S.; Falzarano, S.; Langs, L.; et al. Mapping moderate-scale land-cover over very large geographic areas within a collaborative framework: A case study of the Southwest Regional Gap Analysis Project (SWReGAP). Remote Sens. Environ. 2007, 108, 59–73. [Google Scholar] [CrossRef]
- Lowry, J.H., Jr.; Ramsey, R.D.; Thomas, K.A.; Schrupp, D.; Kepner, W.; Sajwaj, T.; Kirby, J.; Waller, E.; Schrader, S.; Falzarano, S.; et al. Land cover classification and mapping. In Southwest Regional Gap Analysis Final Report; Prior-Magee, J.S., Ed.; US Geological Survey, Gap Analysis Program: Moscow, ID, USA, 2007; Chapter 2. Available online: http://swregap. nmsu. edu/ (accessed on 1 May 2018).
- Rogers, B.M.; Jantz, P.; Goetz, S.J. Vulnerability of eastern US tree species to climate change. Glob. Chang. Biol. 2017, 23, 3302–3320. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.M.; McGill, B.J. Land use and life history limit migration capacity of eastern tree species. Glob. Ecol. Biogeogr. 2018, 27, 57–67. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scenario | Description |
|---|---|
| Base Case (BC) | Represents a medium fertility rate, medium domestic migration, and medium international migration (US Census midline scenario). |
| A1 | Represents fast economic growth, low population growth, and high global integration. Fertility is low with high domestic and international migration. |
| B1 | Represents a globally integrated world but with more emphasis on environmentally sustainable economic development. Fertility and domestic migration are low, while international migration is high. |
| A2 | Represents continued economic development with more regional focus and slower economic convergence between regions. Fertility and domestic migration are high and international migration is medium. |
| B2 | Represents a regionally oriented world of moderate population growth and local solutions to environmental and economic issues. The fertility rate is medium with low domestic migration and medium international migration. |
| Land Cover | Scenarios | ||||
|---|---|---|---|---|---|
| A1 | A2 | B1 | B2 | BC | |
| Exurban | 4022 | 1078 | 820 | 878 | 377 |
| Suburban | 5007 | 18,364 | 274 | 700 | 333 |
| Urban | 188 | 95 | 3 | 6 | 3 |
| Natural Areas | −9217 | −19,537 | −1097 | −1583 | −712 |
| Percent Natural Area Loss | −1.65% | −3.49% | −0.20% | −0.28% | −0.13% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samson, E.A.; Boykin, K.G.; Kepner, W.G.; Andersen, M.C.; Fernald, A. Evaluating Biodiversity Metric Response to Forecasted Land Use Change in the Northern Rio Grande Basin. Environments 2018, 5, 91. https://doi.org/10.3390/environments5080091
Samson EA, Boykin KG, Kepner WG, Andersen MC, Fernald A. Evaluating Biodiversity Metric Response to Forecasted Land Use Change in the Northern Rio Grande Basin. Environments. 2018; 5(8):91. https://doi.org/10.3390/environments5080091
Chicago/Turabian StyleSamson, Elizabeth A., Kenneth G. Boykin, William G. Kepner, Mark C. Andersen, and Alexander Fernald. 2018. "Evaluating Biodiversity Metric Response to Forecasted Land Use Change in the Northern Rio Grande Basin" Environments 5, no. 8: 91. https://doi.org/10.3390/environments5080091
APA StyleSamson, E. A., Boykin, K. G., Kepner, W. G., Andersen, M. C., & Fernald, A. (2018). Evaluating Biodiversity Metric Response to Forecasted Land Use Change in the Northern Rio Grande Basin. Environments, 5(8), 91. https://doi.org/10.3390/environments5080091