Evaluating Presence Data versus Expert Opinions to Assess Occurrence, Habitat Preferences and Landscape Permeability: A Case Study of Butterflies

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Species Data
2.3. Land-Cover Data
2.4. Data Preparation and Analysis
2.5. Expert Opinions on Suitability and Permeability
2.6. Preparation of Maps
2.7. Comparison of Expert Opinions and Ivlev’s Electivity Indices
2.8. Evaluation of Habitat Preference Results by Experts
3. Results
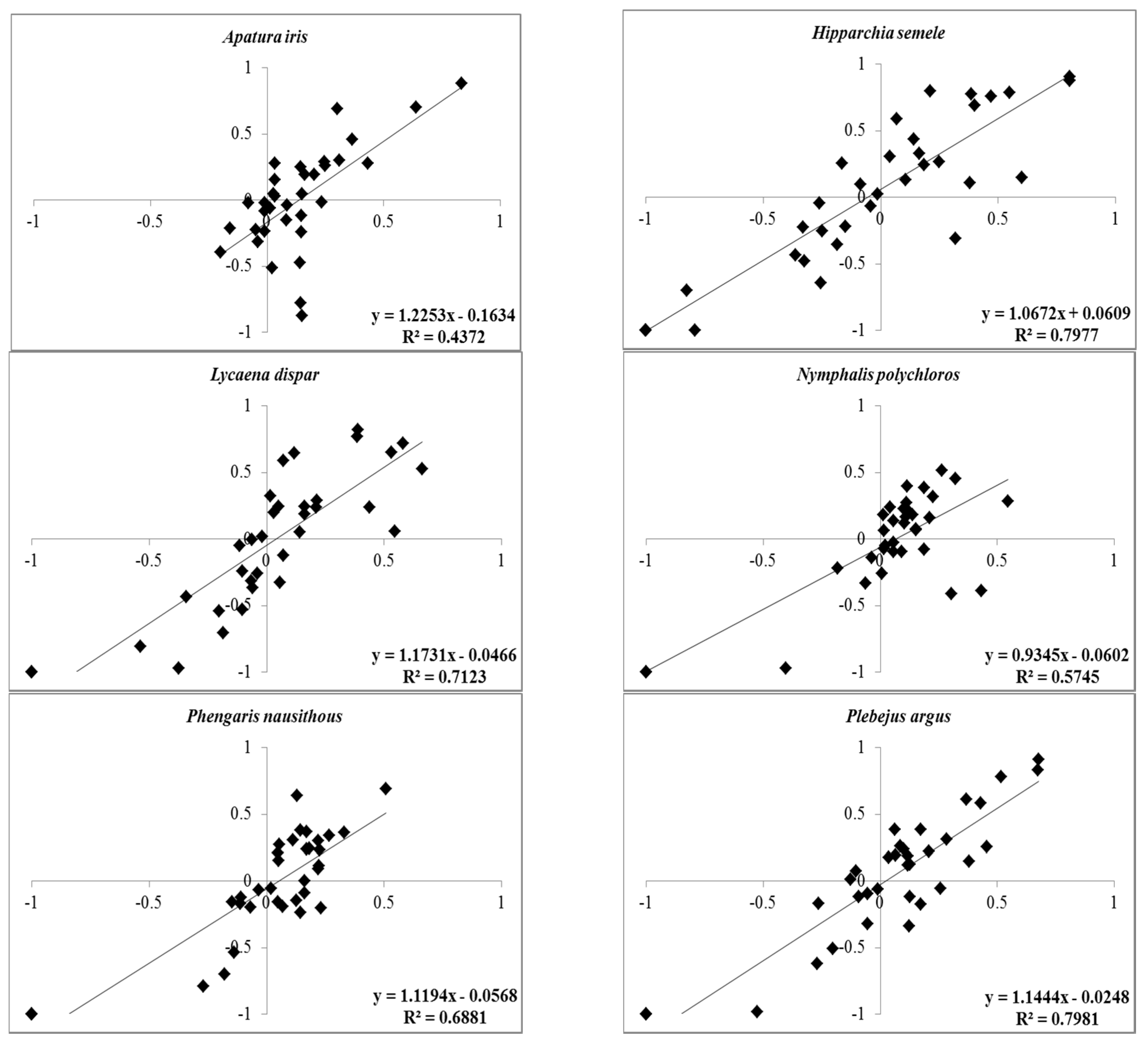
3.1. Relationship between Presence-Based Indices: Two Means for Calculating Ivlev’s Electivity Index
3.2. Effect of Area on Ivlev’s Electivity Indices
3.3. Expert Opinion Habitat Suitability Variance Analysis
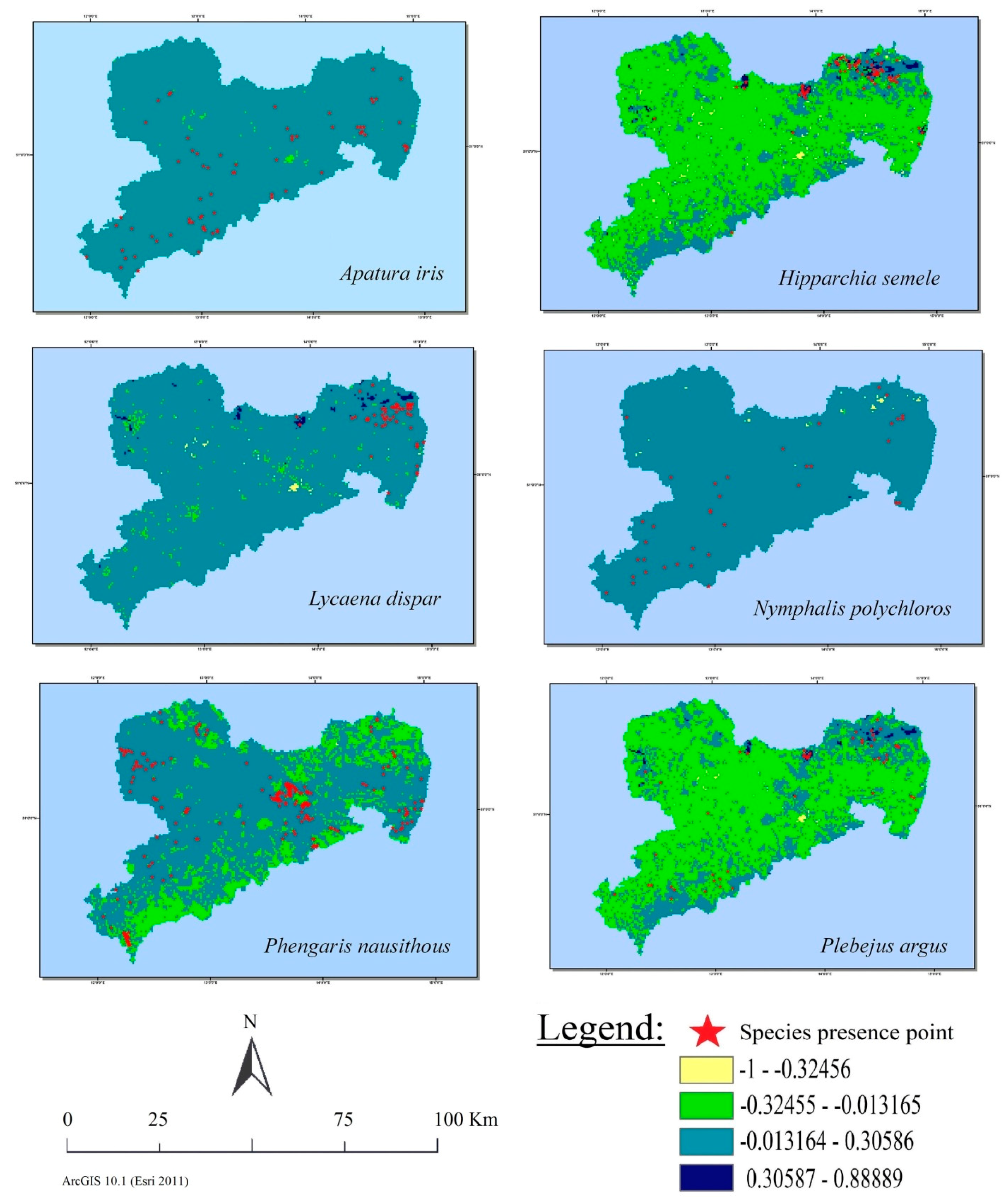
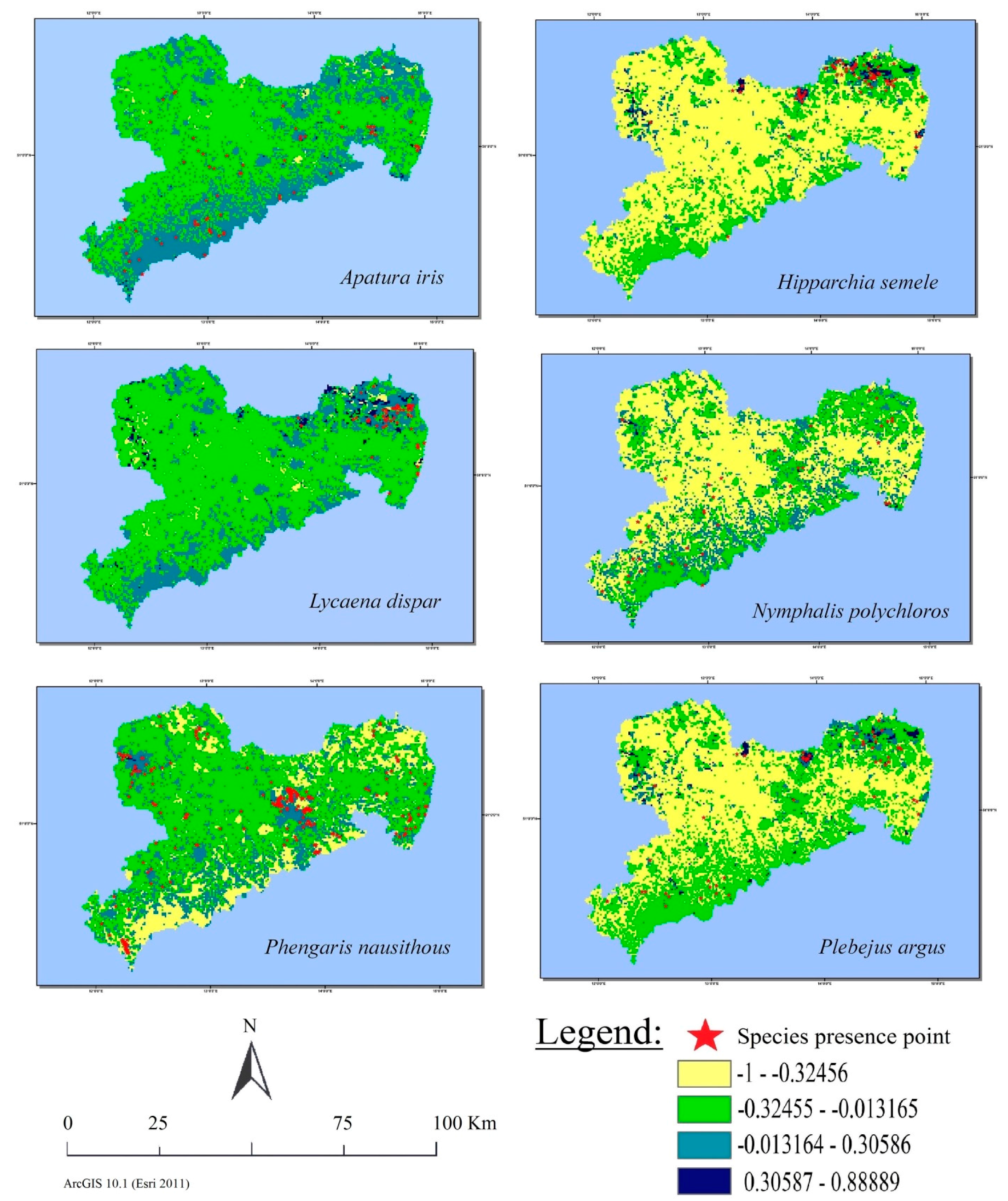
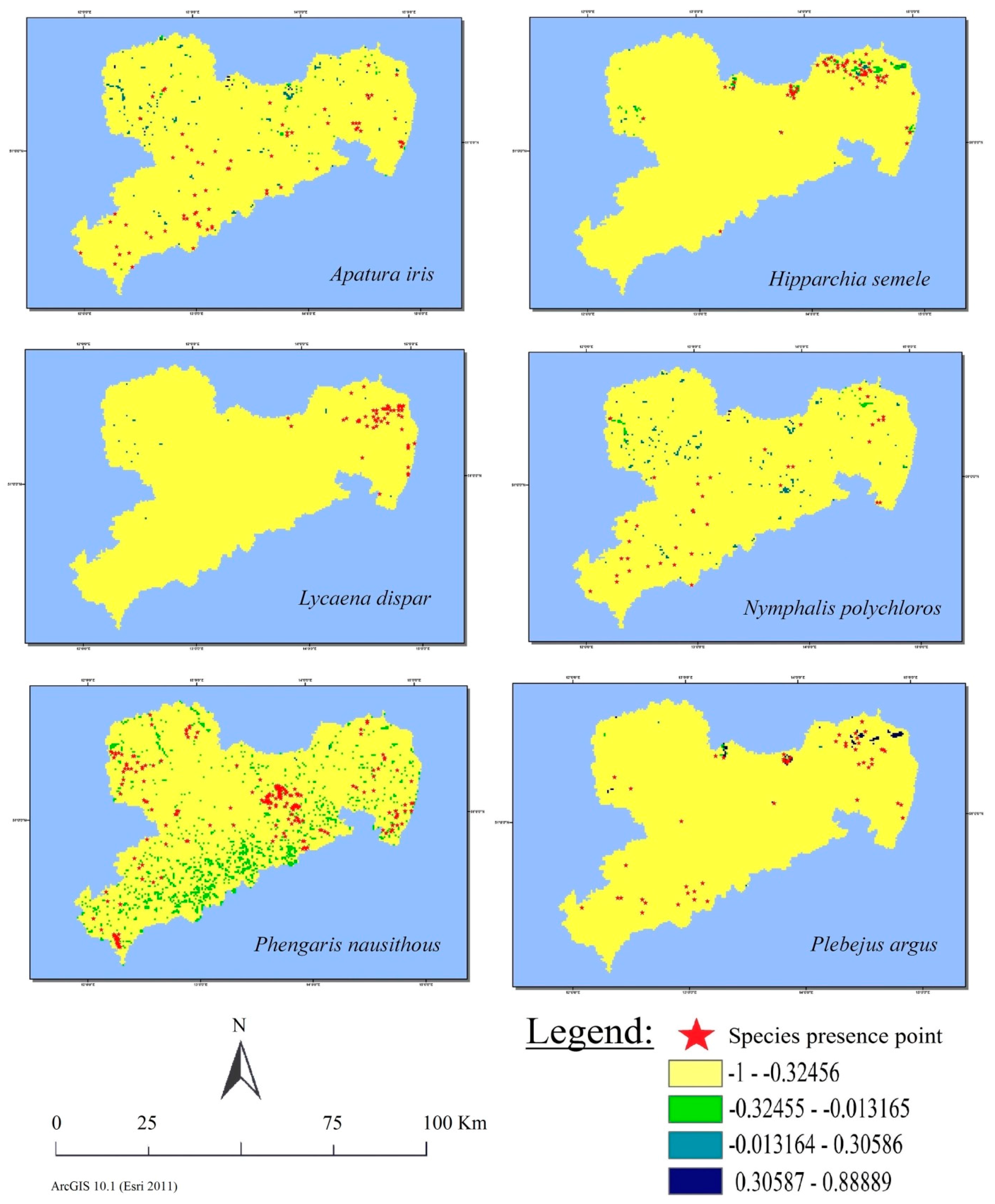
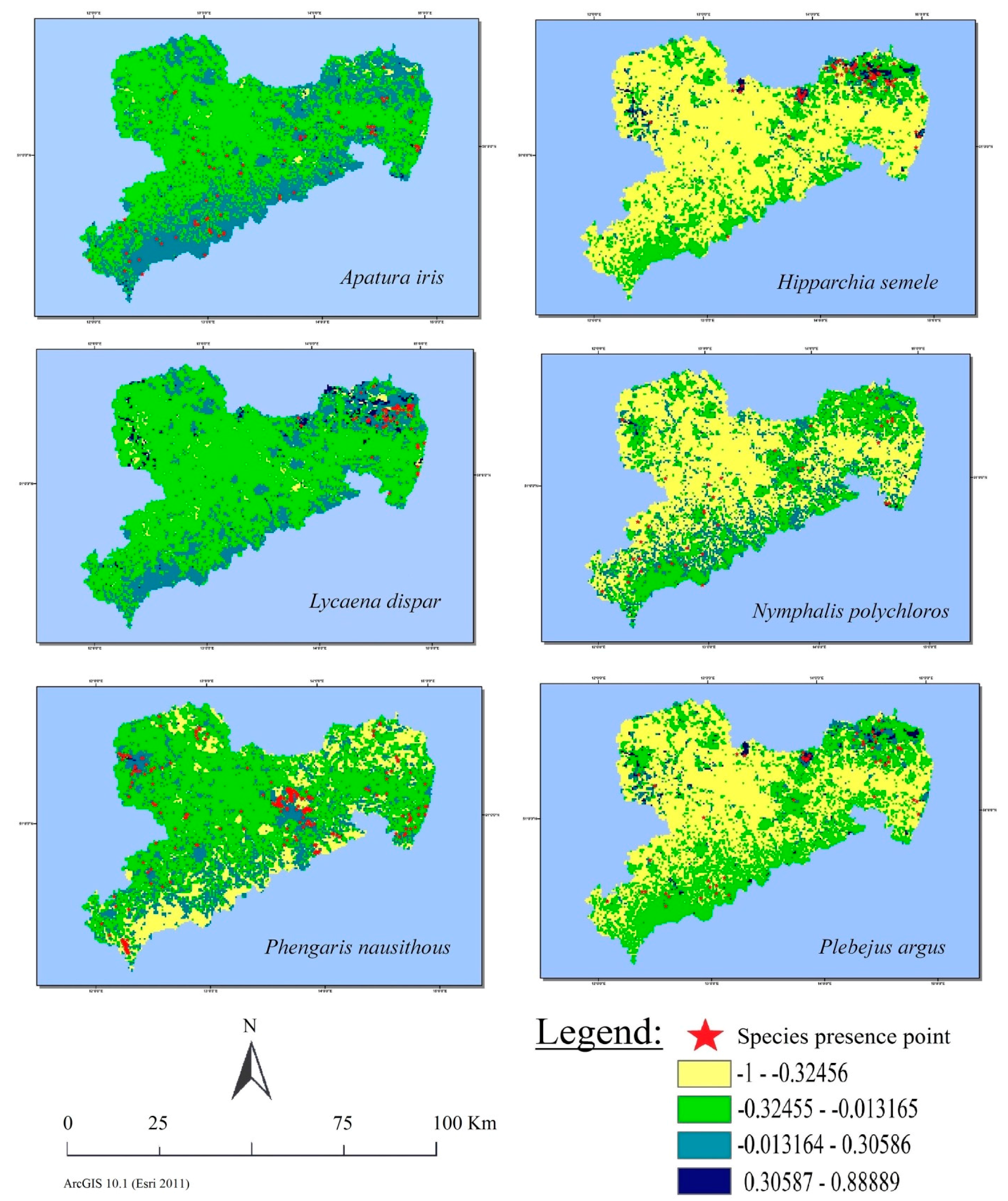
3.4. Visual Analysis of Land Cover Class Suitability Maps
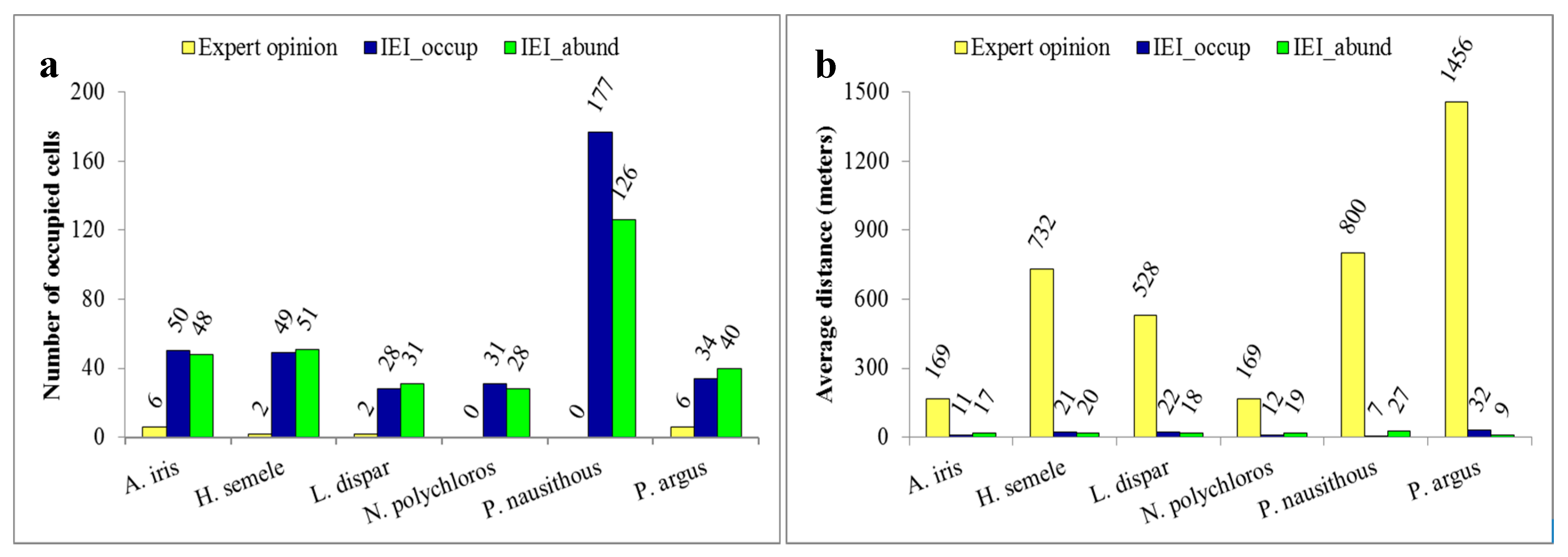
3.5. Spatial Relation between Observations and Suitable Habitats
3.6. Outcomes of Evaluation of the Results by Experts
3.7. Land Covers Class Permeability versus Suitability
4. Discussion
4.1. Empirical Presence Data versus Expert Opinions for Habitat Suitability
4.2. Landscape Permeability
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A. Land-Cover Types and Habitat Types (BTLNK) of Saxony Vector Map (Lfulg 2008) [51]
| BTLNK | Original German Land Cover Description | English Translation of Land Cover Class |
| 21 | Flieβgewässer | streaming water |
| 23 | Stillgewässer | standing waterbody |
| 24 | Gewässerbegleitende Vegetation | riparian vegetation |
| 25 | Bauwerke an Gewässern | construction at waterbody |
| 31 | Hochmoor, Zwischenmoor | raised bogs and transitional mires |
| 32 | Niedermoor, Sumpf | fens and swamp land |
| 41 | Wirtschaftsgrünland | grasslands and (managed) meadows |
| 42 | Ruderalflur, Staudenflur | ruderal and herbaceous vegetation |
| 51 | Anstehender Fels | bedrock |
| 52 | Blockschutthalden | scree slopes |
| 53 | größere Lesesteinhaufen und offene Steinrücken | large clearance cairns and open rocks |
| 54 | Offene Flächen | open areas |
| 55 | Zwergstrauchheiden und Borstgrasrasen | dwarf shrub heath and Nardus grassland |
| 56 | Magerrasen trockener Standorte | dry abandoned grasslands |
| 61 | Feldgehölz, Baumgruppe 100 m2–1 ha | group of trees 100 m2–1 ha |
| 66 | Gebüsch | scrubland |
| 67 | neu ab 2005: Streuobstwiese | meadow with fruit trees |
| 70 | Wiederaufforstung | reforestation |
| 71 | Laubbaumart (Reinbestand) | broad leaved forest |
| 72 | Nadelbaumart (Reinbestand) | coniferous forest |
| 73 | Laub-Nadel-Mischwald | mixed forest (broad-leaved-coniferous) |
| 74 | Nadel-Laub-Mischwald | mixed forest (coniferous-broad-leaved) |
| 75 | Laubmischwald | mixed forest (broad-leaved) |
| 76 | Nadelmischwald | mixed forest (coniferous) |
| 77 | Feuchtwälder (Moorwald siehe 31300) | mesic forest |
| 78 | Waldrandbereiche/Vorwälder | forest edges, pioneer forest |
| 79 | Erstaufforstung | afforestation |
| 81 | Acker | arable land |
| 82 | Sonderkulturen | specialised crop |
| 91 | Wohngebiet | residential area |
| 92 | Mischgebiet | mixed area |
| 93 | Gewerbegebiet | industrial area |
| 94 | Grün- und Freiflächen | green spaces |
| 95 | Verkehrsflächen | traffic area |
| 96 | Anthropogen genutzte Sonderflächen | human used special areas |
Appendix B. Habitat Suitability Analysis Based on Expert Opinions versus IEIoccup for Six Butterfly Species
| Species Name | BTLNK | BTLNK-Defination | Status | IEIoccup | Expert |
| Apatura iris | 75 | deciduous mixed forests | Match | high | high |
| 78 | forest edges, pioneer forest | Match | high | high | |
| 67 | meadow with fruit trees | Match | low | low | |
| 82 | specialized crop | Match | low | low | |
| 25 | construction at water body | Mismatch | high | low | |
| 52 | scree slopes | Mismatch | high | low | |
| Hipparchia semele | 56 | dry abandoned grasslands | Match | high | high |
| 54 | open areas | Match | high | high | |
| 67 | meadow with fruit trees | Match | low | low | |
| 82 | specialized crop | Match | low | low | |
| 31 | raised bogs and transitional mires | Mismatch | high | low | |
| 32 | fens and swamp land | Mismatch | high | low | |
| 51 | bedrock | Mismatch | low | high | |
| 52 | scree slopes | Mismatch | low | high | |
| Lycaena dispar | 24 | riparian vegetation | Match | high | high |
| 32 | fens and swamp land | Match | high | high | |
| 52 | scree slopes | Match | low | low | |
| 53 | large clearance cairns and open rocks | Match | low | low | |
| 25 | construction at water body | Mismatch | high | low | |
| 56 | dry abandoned grasslands | Mismatch | high | low | |
| Nymphalis polychloros | 78 | forest edges, pioneer forest | Match | high | high |
| 75 | deciduous mixed forests | Match | high | high | |
| 52 | scree slopes | Match | low | low | |
| 31 | raised bogs and transitional mires | Match | low | low | |
| 25 | construction at water body | Mismatch | high | low | |
| 51 | bedrock | Mismatch | high | low | |
| Phengaris nausithous | 24 | riparian vegetation | Match | high | high |
| 32 | fens and swamp land | Match | high | high | |
| 31 | raised bogs and transitional mires | Match | low | low | |
| 52 | scree slopes | Match | low | low | |
| 82 | specialized crop | Mismatch | high | low | |
| Plebejus argus | 55 | dwarf shrub heath and Nardus grassland | Match | high | high |
| 56 | dry abandoned grasslands | Match | high | high | |
| 51 | bedrock | Match | low | low | |
| 52 | scree slopes | Match | low | low | |
| 32 | fens and swamp land | Mismatch | high | low | |
| 77 | mesic forest | Mismatch | high | low |
Appendix C. Habitat Suitability Analysis Based on Expert Opinions versus IEIabund for Six Butterfly Species
| Species Name | BTLNK | BTLNK-Defination | Status | IEIabund | Expert |
| Apatura iris | 51 | bedrock | Match | low | low |
| 55 | dwarf shrub heath | Match | low | low | |
| 25 | construction at water body | Mismatch | high | low | |
| 52 | scree slopes | Mismatch | high | low | |
| 77 | mesic forest | Mismatch | low | high | |
| 78 | forest edges, pioneer forest | Mismatch | low | high | |
| Hipparchia semele | 55 | dwarf shrub heath | Match | high | high |
| 56 | dry abandoned grasslands | Match | high | high | |
| 67 | meadow with fruit trees | Match | low | low | |
| 81 | arable land | Match | low | low | |
| 32 | fens and swamp land | Mismatch | high | low | |
| 77 | mesic forest | Mismatch | high | low | |
| 51 | bedrock | Mismatch | low | high | |
| 52 | scree slopes | Mismatch | low | high | |
| Lycaena dispar | 24 | riparian vegetation | Match | high | high |
| 32 | fens and swamp land | Match | high | high | |
| 52 | scree slopes | Match | low | low | |
| 53 | large clearance and open rocks | Match | low | low | |
| 55 | dwarf shrub heath | Mismatch | high | low | |
| 77 | mesic forest | Mismatch | high | low | |
| Nymphalis polychloros | 75 | mixed forest (broad-leaved) | Match | high | high |
| 78 | forest edges, pioneer forest | Match | high | high | |
| 31 | raised bogs and transitional mires | Match | low | low | |
| 52 | scree slopes | Match | low | low | |
| 24 | riparian vegetation | Mismatch | high | low | |
| 76 | mixed forest (coniferous) | Mismatch | high | low | |
| 67 | meadow with fruit trees | Mismatch | low | high | |
| 82 | specialized crop | Mismatch | low | high | |
| Phengaris nausithous | 51 | bedrock | Match | low | low |
| 52 | scree slopes | Match | low | low | |
| 82 | specialized crop | Mismatch | high | low | |
| Plebejus argus | 55 | dwarf shrub heath | Match | high | high |
| 56 | dry abandoned grasslands | Match | high | high | |
| 51 | bedrock | Match | low | low | |
| 52 | scree slopes | Match | low | low | |
| 32 | fens and swamp land | Mismatch | high | low | |
| 77 | mesic forest | Mismatch | high | low |
References
- Keller, B.D.; Gleason, D.F.; McLeod, E.; Woodley, C.M.; Airamé, S.; Causey, B.D.; Friedlander, A.M.; Grober-Dunsmore, R.; Johnson, J.E.; Miller, S.L.; et al. Climate change, coral reef ecosystems, and management options for marine protected areas. Environ. Manag. 2009, 44, 1069–1088. [Google Scholar] [CrossRef] [PubMed]
- Convention on Biological Diversity. Available online: http://www.cbd.int/decision/cop/?id=12268 (accessed on 7 December 2016).
- Margules, C.R.; Pressey, R.L. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Kingsland, S.E. Creating a science of nature reserve design: Perspectives from history. Environ. Model. Assess. 2002, 7, 61–69. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, T. Biodiversity informatics: Managing and applying primary biodiversity data. Philos. Trans. R. Soc. Lond. B 2004, 359, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Guralnick, R.P.; Hill, A.W.; Lane, M. Towards a collaborative, global infrastructure for biodiversity assessment. Ecol. Lett. 2007, 10, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Hortal, J.; Lobo, J.M.; Jimenez-Valverde, A. Limitations of biodiversity databases: Case study on seed-plant diversity in Tenerife, Canary Islands. Conserv. Biol. 2007, 21, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Gibson, L.A.; Wilson, B.A.; Cahill, D.M.; Hill, J. Spatial prediction of rufous bristlebird habitat in a coastal heathland: A GIS-based approach. J. Appl. Ecol. 2004, 41, 213–223. [Google Scholar] [CrossRef]
- Posillico, M.; Meriggi, A.; Pagnin, E.; Lovari, S.; Russo, L. A habitat model for brown bear conservation and land use planning in the central Apennines. Biol. Conserv. 2004, 118, 141–150. [Google Scholar] [CrossRef]
- Wintle, B.A.; Elith, J.; Potts, J.M. Fauna habitat modelling and mapping: A review and case study in the Lower Hunter Central Coast region of NSW. Austral Ecol. 2005, 30, 719–738. [Google Scholar] [CrossRef]
- Lobo, J.M.; Lumaret, J.P.; JayRobert, P. Taxonomic databases as tools in spatial biodiversity research. Ann. Soc. Entomol. Fr. 1997, 33, 129–138. [Google Scholar]
- Hanski, I.; Gaggiotti, O.E. Ecology, Genetics and Evolution of Metapopulations; Academic Press: Burlington, VT, USA, 2004; pp. ix–xix. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Pullin, A.S.; Knight, T.M. Effectiveness in conservation practice: Pointers from medicine and public health. Conserv. Biol. 2001, 15, 50–54. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Whigham, P.A. Induction of a marsupial density model using genetic programming and spatial relationships. Ecol. Model. 2000, 131, 299–317. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Guisan, A. Which is the optimal sampling strategy for habitat suitability modelling? Ecol. Model. 2002, 157, 331–341. [Google Scholar] [CrossRef]
- Cawsey, E.M.; Austin, M.P.; Baker, B.L. Regional vegetation mapping in Australia: A case study in the practical use of statistical modeling. Biodivers. Conserv. 2002, 11, 2239–2274. [Google Scholar] [CrossRef]
- Graham, C.H.; Ferrier, S.; Huettman, F.; Moritz, C.; Peterson, A.T. New developments in museum based informatics and applications in biodiversity analysis. Trends Ecol. Evol. 2004, 19, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Huettmann, F. Databases and science-based management in the context of wildlife and habitat: Towards a certified ISO standard for objective decision-making for the global community by using the internet. J. Wildl. Manag. 2005, 69, 466–472. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, A.T. Interpretation of models of fundamental ecological niches and species distributional areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef]
- Noss, R.F.; Cooperrider, A.Y. Saving Nature’s Legacy: Protecting and Restoring Biodiversity; Island Press: Washington, DC, USA, 1994; pp. 1–416. [Google Scholar]
- Prevedello, J.A.; Vieira, M.V. Does the type of matrix matter? A quantitative review of the evidence. Biodivers. Conserv. 2010, 19, 1205–1223. [Google Scholar] [CrossRef]
- Hall, L.S.; Krausman, P.R.; Morrison, M.L. The habitat concept and a plea for standard terminology. Wildl. Soc. B 1997, 25, 173–182. [Google Scholar]
- Dennis, R.L.H.; Shreeve, T.G.; Dyck, H.V. Habitats and resources: The need for a resource-based definition to conserve butterflies. Biodivers. Conserv. 2006, 15, 1943–1966. [Google Scholar] [CrossRef]
- Vanreusel, W.; Dyck, H.V. When functional habitat does not match vegetation types: A resource-based approach to map butterfly habitat. Biol. Conserv. 2007, 135, 202–211. [Google Scholar] [CrossRef]
- Meiklejohn, K.; Ament, R.; Tabor, G. Habitat Corridors & Landscape Connectivity: Clarifying the Terminology; Center for Large Landscape Conservation: Bozeman, MT, USA, 2010; pp. i–ix. [Google Scholar]
- Lindenmayor, D.; Fischer, J. Habitat Fragmentation and Landscape Change: An Ecological and Conservation Synthesis; Island Press: Washington, DC, USA, 2006; pp. 1–352. [Google Scholar]
- Heller, N.E.; Zavaleta, E.S. Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biol. Conserv. 2009, 142, 14–32. [Google Scholar] [CrossRef]
- Krosby, M.; Tewksbury, J.; Haddad, N.M.; Hoekstra, J. Ecological connectivity for a changing climate. Conserv. Biol. 2010, 24, 1686–1689. [Google Scholar] [CrossRef] [PubMed]
- Hovestadt, T.; Binzenhöfer, B.; Nowicki, P.; Settele, J. Do all inter-patch movements represent dispersal? A mixed kernel study of butterfly mobility in fragmented landscapes. J. Anim. Ecol. 2011, 80, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Drew, C.A.; Collazo, J.A. Expert knowledge as a foundation for the management of secretive species and their habitat (Chapter 5). In Expert Knowledge and Its Application in Landscape Ecology; Perera, A.H., Drew, C.A., Johnson, C.J., Eds.; Springer: New York, NY, USA, 2011; pp. 87–107. [Google Scholar]
- Drew, C.A.; Perera, A.H. Expert knowledge as a basis for landscape ecological predictive models (Chapter 12). In Predictive Species and Habitat Modeling in Landscape Ecology: Concepts and Applications; Drew, C.A., Wiersma, Y.F., Huettmann, F., Eds.; Springer: New York, NY, USA, 2011; pp. 229–248. [Google Scholar]
- Clevenger, A.P.; Wierzchowski, J.; Chruszcz, B.; Gunson, K. GIS-generated, expert-based models for identifying wildlife habitat linkages and planning mitigation passages. Conserv. Biol. 2002, 16, 503–514. [Google Scholar] [CrossRef]
- Johnson, C.J.; Gillingham, M.P. Mapping uncertainty: Sensitivity of wildlife habitat ratings to Expert opinion. J. Appl. Ecol. 2004, 41, 1032–1041. [Google Scholar] [CrossRef]
- Choy, S.L.; O’Leary, R.; Mengersen, K. Elicitation by design for ecology: Using expert opinion to inform priors for Bayesian statistical models. Ecology 2009, 90, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Schlossberg, S.; King, D.I. Modeling animal habitats based on cover types: A critical review. Environ. Manag. 2009, 43, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.J.; Hurley, M.; Rapaport, E.; Pullinger, M. Using expert knowledge effectively: Lessons from species distribution models for wildlife conservation and management (Chapter 8). In Expert Knowledge and Its Application in Landscape Ecology; Perera, A.H., Drew, C.A., Johnson, C.J., Eds.; Springer: New York, NY, USA, 2012; pp. 153–171. [Google Scholar]
- Thomas, J.A.; Telfer, M.G.; Roy, D.B.; Preston, C.D.; Greenwood, J.J.; Asher, J.; Fox, R.; Clarke, R.T.; Lawton, J.H. Comparative losses of British butterflies, birds and plants and the global extinction crisis. Science 2004, 303, 1879–1881. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.A. Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Philos. Trans. R. Soc. Lond. B 2005, 360, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Pe’er, G.; Settele, J. Butterflies in and for conservation: Trends and Prospects. Isr. J. Ecol. Evol. 2008, 54, 7–17. [Google Scholar] [CrossRef]
- Van Swaay, C.; Van Strien, A.; Aghababyan, K.; Astrom, S.; Botham, M.; Brereton, T.; Chambers, P.; Collins, S.; Domenech Ferre, M.; Escobes, R.; et al. The European Butterfly Indicator for Grassland Species: 1990–2013; De Vlinderstichting: Wageningen, The Netherlands, 2015; pp. 1–39. [Google Scholar]
- Spänhoff, B.; Dimmer, R.; Friese, H.; Harnapp, S.; Herbst, F.; Jenemann, K.; Mickel, A.; Rohde, S.; Schönherr, M.; Ziegler, K.; et al. Ecological status of rivers and streams in Saxony (Germany) according to the water framework directive and prospects of improvement. Water 2012, 4, 887–904. [Google Scholar] [CrossRef]
- Renner, M.; Bernhofer, C. Long term variability of the annual hydrological regime and sensitivity to temperature phase shifts in Saxony/Germany. Hydrol. Earth Syst. Sci. 2011, 15, 1819–1833. [Google Scholar] [CrossRef]
- Bastian, O. Landscape classification in Saxony (Germany)—A tool for holistic regional planning. Landsc. Urban Plan. 2010, 50, 145–155. [Google Scholar] [CrossRef]
- Gimenez-Dixon, M. Lycaena dispar. IUCN 2012. IUCN Red List of Threatened Species; Version 2012.2. Available online: www.iucnredlist.org (accessed on 8 July 2015).
- World Conservation Monitoring Centre. Phengaris nausithous. IUCN 2012. IUCN Red List of Threatened Species; Version 2012.2. 1996. Available online: www.iucnredlist.org (accessed on 13 December 2013).
- Van Swaay, C.; Wynhoff, I.; Verovnik, R.; Wiemers, M.; López Munguira, M.; Maes, D.; Sasic, M.; Verstrael, T.; Warren, M.; Settele, J. Hipparchia semele. IUCN 2012. IUCN Red List of Threatened Species; Version 2012.2. 2010. Available online: www.iucnredlist.org (accessed on 22 December 2013).
- Belgian Species List. Nymphalis polychloros (Linnaeus, 1758). Available online: http://www.species.be/en/3063 (accessed on 15 October 2015).
- LfULG. Biotope and Land Use Map of Saxony, Germany 2005. 2008. Available online: http://www.umwelt.sachsen.de/umwelt/natur/18615.htm (accessed on 11 April 2012).
- Ivlev, V.S. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CT, USA, 1961; pp. 1–302. [Google Scholar]
- Aryal, A.; Raubenheimer, D.; Subedi, S.; Kattel, B. Spatial habitat overlap and habitat preference of Himalayan Musk Deer (Moschus chrysogaster) in Sagarmatha (Mt. Everest) national park, Nepal. J. Biol. Sci. 2010, 2, 217–225. [Google Scholar]
- Storch, I. On spatial resolution in habitat models: Can small-scale forest structure explain Capercaillie numbers? Conserv. Ecol. 2002, 6, 6. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Pearce, J.L.; Cherry, K.; Drielsma, M.; Ferrier, S.; Whish, G. Incorporating expert opinion and fine-scale vegetation mapping into statistical models of faunal distribution. J. Appl. Ecol. 2001, 38, 412–424. [Google Scholar] [CrossRef]
- Brooks, R.P. Improving habitat suitability index models. Wildl. Soc. B 1997, 25, 163–167. [Google Scholar]
- Henle, K.; Potts, S.; Kunin, W.; Matsinos, Y.; Similä, J.; Pantis, J.; Grobelnik, V.; Penev, L.; Settele, J. Scaling in Ecology and Biodiversity Conservation; Pensoft Publishers: Sofia, Bulgaria, 2014; p. e1169. [Google Scholar]
- Edenius, L.; Elmberg, J. Landscape level effects of modern forestry on bird communities in North Swedish boreal forests. Landsc. Ecol. 1996, 11, 325–338. [Google Scholar] [CrossRef]
- Saab, V. Importance of spatial scale to habitat use by breeding birds in riparian forests: A hierarchical analysis. Ecol. Appl. 1999, 9, 135–151. [Google Scholar] [CrossRef]
- Graf, R.F.; Bollmann, K.; Suter, W.; Bugmann, H. The importance of spatial scale in habitat models: Capercaillie in the Swiss Alps. Landsc. Ecol. 2005, 20, 703–717. [Google Scholar] [CrossRef]
- Reinhardt, R.; Sbieschne, H.; Settele, J.; Fischer, U.; Fiedler, G. Tagfalter von Sachsen; Entomologische Nachrichten und Berichte: Dresden, Germany, 2007; pp. 1–695. [Google Scholar]
- Settele, J.; Kudrna, O.; Harpke, A.; Kühn, I.; van Swaay, C.; Verovnik, R.; Warren, M.; Wiemers, M.; Hanspach, J.; Hickler, T.; et al. Climatic Risk Atlas of European Butterflies. BioRisk 2008, 1, 1–710. [Google Scholar] [CrossRef]
- Pe’er, G.; Henle, K.; Dislich, C.; Frank, K. Breaking functional connectivity into components: A novel approach using an Individual-Based model, and first outcomes. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Nowicki, P.; Vrabec, V.; Binzenhofer, B.; Feil, J.; Zaksek, B.; Hovestadt, T.; Settele, J. Butterfly dispersal in inhospitable matrix: Rare, risky, but long-distance. Landsc. Ecol. 2014, 29, 401–412. [Google Scholar] [CrossRef]
- Dover, J.; Settele, J. The influences of landscape structure on butterfly distribution and movement: A review. J. Insect Conserv. 2009, 13, 3–27. [Google Scholar] [CrossRef]
- Kühn, E.; Feldmann, R.; Harpke, A.; Hirneisen, N.; Musche, M.; Leopold, P.; Settele, J. Getting the public involved into butterfly conservation—Lessons learned from a new monitoring scheme in Germany. Isr. J. Ecol. Evol. 2008, 54, 89–103. [Google Scholar] [CrossRef]
- Moilanen, A.; Kujala, H.; Leathwick, J. The Zonation framework and software for conservation prioritization. In Spatial Conservation Prioritization: Quantitative Methods and Computational Tools; Moilanen, A., Wilson, K.A., Possingham, H.P., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 196–210. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Indices | R2-Value * | R2-Value ** | Equation *** |
|---|---|---|---|---|
| Apatura iris | IEIoccup | 0.0133 | 0.025 | y = −1E− 08x + 0.1581 |
| IEIabund | 0.0055 | 0.0577 | y = 3E − 08x − 0.0522 | |
| Hipparchia semele | IEIoccup | 0.091 | 0.2166 | y = 1E − 07x − 0.2474 |
| IEIabund | 0.1048 | 0.1863 | y = 1E − 07x − 0.12 | |
| Lycaena dispar | IEIoccup | 0.0078 | 0.0287 | y = 3E − 08x − 0.0123 |
| IEIabund | 0.0191 | 0.0733 | y = 7E − 08x − 0.0935 | |
| Nymphalis polychloros | IEIoccup | 0.0026 | 0.0277 | y = 5E − 08x − 0.0158 |
| IEIabund | 0.0135 | 0.0635 | y = 8E − 08x − 0.0747 | |
| Phengaris nausithous | IEIoccup | 0.0055 | 0.0377 | y = 2E − 08x − 0.0234 |
| IEIabund | 0.0163 | 0.1343 | y = 4E − 08x − 0.1555 | |
| Plebejus argus | IEIoccup | 0.0083 | 0.0557 | y = 5E − 08x − 0.0104 |
| IEIabund | 0.029 | 0.1128 | y = 9E − 08x − 0.0488 |
| Species Name | Habitat Suitability | R2-Value | Equation |
|---|---|---|---|
| Apatura iris | IEIoccup | 0.0057 | y = − 0.0315x + 0.2181 |
| IEIabund | 0.0076 | y = − 0.0676x + 0.1755 | |
| Expert opinions avg | 0.3761 | y = 0.4428x + 0.9587 | |
| Hipparchia semele | IEIoccup | 0.0005 | y = 0.0183x − 0.0871 |
| IEIabund | 0.0015 | y = − 0.0379x + 0.1334 | |
| Expert opinions avg | 0.6745 | y = 0.8973x − 0.3351 | |
| Lycaena dispar | IEIoccup | 0.0018 | y = − 0.0198x + 0.0759 |
| IEIabund | 0.0134 | y = − 0.0749x + 0.1925 | |
| Expert opinions avg | 0.5745 | y = 1.053x − 0.9407 | |
| Nymphalis polychloros | IEIoccup | 0.0082 | y = 0.0646x − 0.1158 |
| IEIabund | 0.0318 | y = 0.1564x − 0.4051 | |
| Expert opinions avg | 0.1229 | y = 0.2138x + 1.7411 | |
| Phengaris nausithous | IEIoccup | 0.0038 | y = − 0.026x + 0.1072 |
| IEIabund | 0.0045 | y = 0.0383x − 0.1423 | |
| Expert opinions avg | 0.5757 | y = 0.9888x − 0.5078 | |
| Plebejus argus | IEIoccup | 0.0007 | y = − 0.0129x + 0.0931 |
| IEIabund | 0.0002 | y = − 0.0088x + 0.0638 | |
| Expert opinions avg | 0.449 | y = 0.7903x + 0.1576 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arfan, M.; Pe’er, G.; Bauch, B.; Settele, J.; Henle, K.; Klenke, R. Evaluating Presence Data versus Expert Opinions to Assess Occurrence, Habitat Preferences and Landscape Permeability: A Case Study of Butterflies. Environments 2018, 5, 36. https://doi.org/10.3390/environments5030036
Arfan M, Pe’er G, Bauch B, Settele J, Henle K, Klenke R. Evaluating Presence Data versus Expert Opinions to Assess Occurrence, Habitat Preferences and Landscape Permeability: A Case Study of Butterflies. Environments. 2018; 5(3):36. https://doi.org/10.3390/environments5030036
Chicago/Turabian StyleArfan, Muhammad, Guy Pe’er, Bianca Bauch, Josef Settele, Klaus Henle, and Reinhard Klenke. 2018. "Evaluating Presence Data versus Expert Opinions to Assess Occurrence, Habitat Preferences and Landscape Permeability: A Case Study of Butterflies" Environments 5, no. 3: 36. https://doi.org/10.3390/environments5030036
APA StyleArfan, M., Pe’er, G., Bauch, B., Settele, J., Henle, K., & Klenke, R. (2018). Evaluating Presence Data versus Expert Opinions to Assess Occurrence, Habitat Preferences and Landscape Permeability: A Case Study of Butterflies. Environments, 5(3), 36. https://doi.org/10.3390/environments5030036