
Monitoring the Status of Mesophotic Biogenic Reefs in the Northern Adriatic Sea: Comparing a Biotic Index and Multivariate Community Patterns

 ,
,  ,
,  ,
,  and
and
Abstract

1. Introduction
2. Materials and Methods
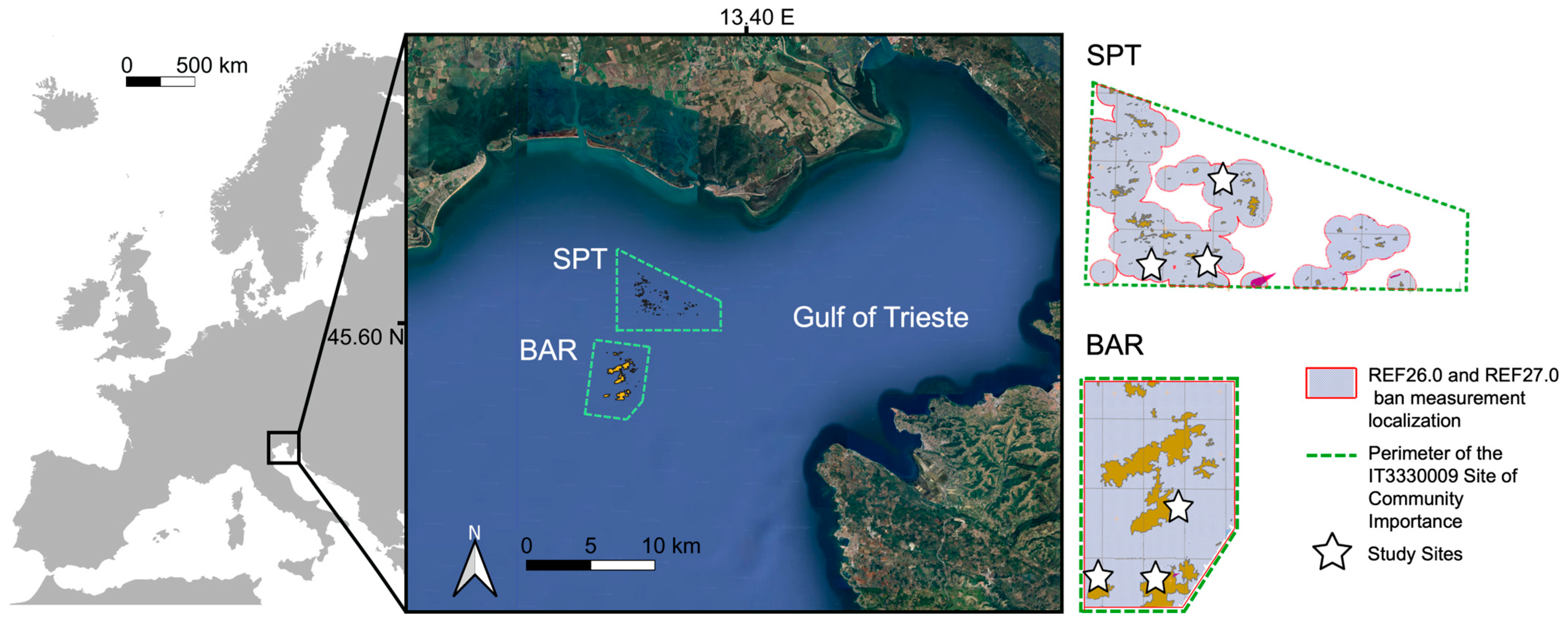
2.1. Study Area and Sampling Design
2.2. Statistical Analysis
2.3. Ecological Status
3. Results
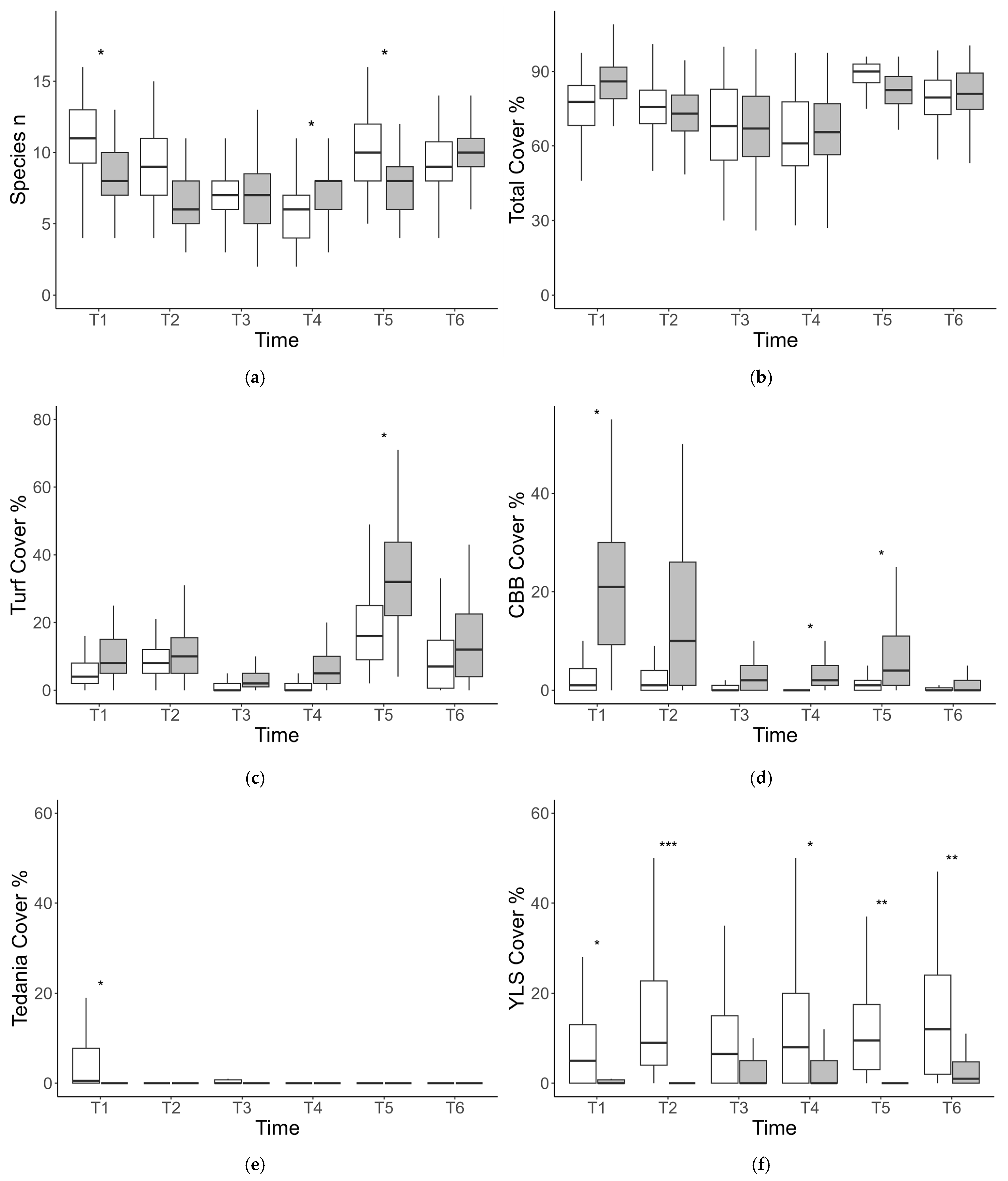
3.1. Structure of Sessile Assemblages
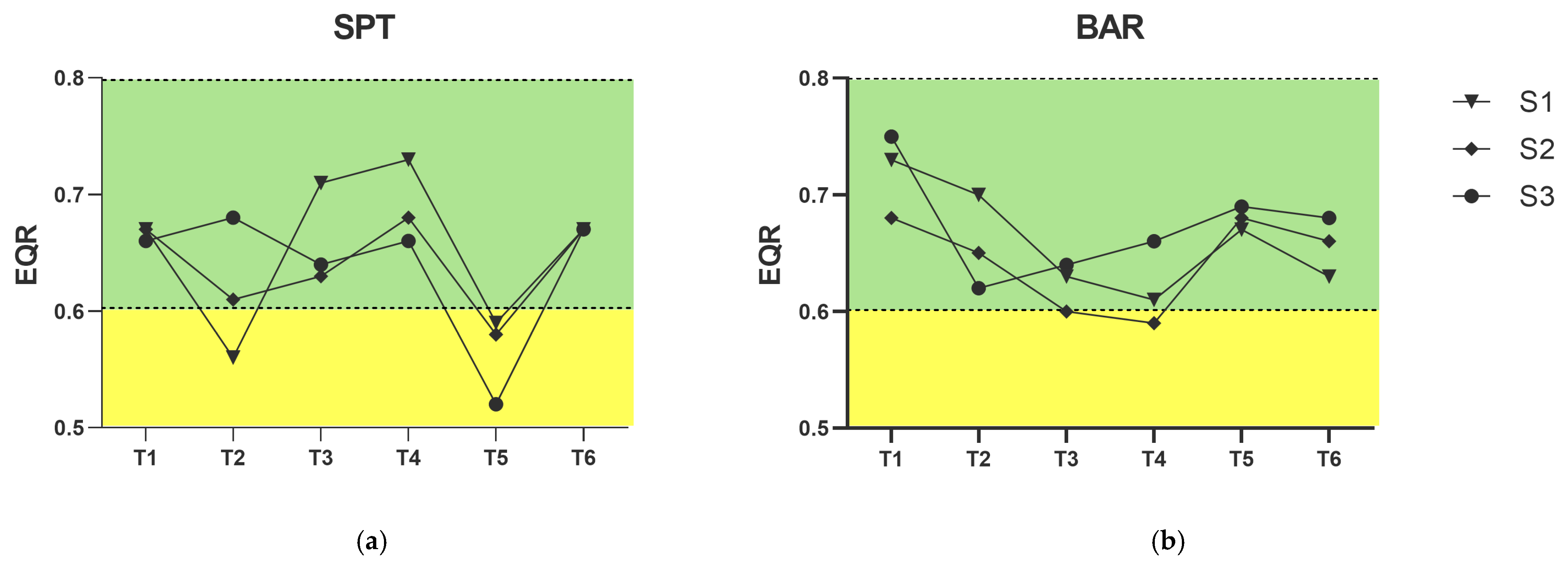
3.2. Ecological Status
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballesteros, E. Mediterranean Coralligenous Assemblages: A Synthesis of Present Knowledge. In Oceanography and Marine Biology: An Annual Review; Gibson, R.N., Atkinson, R.J.A., Gordon, J.D.M., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2006; Volume 44, pp. 123–195. [Google Scholar]
- UNEP/MAP Regional Activity Centre for Specially Protected Areas. Action Plan for the Conservation of the Coralligenous and Other Calcareous Bio-Concretions in the Mediterranean Sea; UNEP MAP RAC-SPA Publication: Tunis, Tunisia, 2008. [Google Scholar]
- Boudouresque, C.F.; Blanfuné, A.; Harmelin-Vivien, M.; Personnic, S.; Ruitton, S.; Thibaut, T.; Verlaque, M. Where Seaweed Forests Meet Animal Forests: The Examples of Macroalgae in Coral Reefs and the Mediterranean Coralligenous Ecosystem. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 369–396. [Google Scholar]
- Garrabou, J.; Ballesteros, E.; Zabala, M. Structure and Dynamics of North-Western Mediterranean Rocky Benthic Communities along a Depth Gradient. Estuar. Coast. Shelf Sci. 2002, 55, 493–508. [Google Scholar] [CrossRef]
- Laborel, J. Marine Biogenic Constructions in the Mediterranean, a Review. Sci. Rep. Port-Cros Natl. Park 1987, 13, 97–126. [Google Scholar]
- Casellato, S.; Stefanon, A. Coralligenous Habitat in the Northern Adriatic Sea: An Overview. Mar. Ecol. 2008, 29, 321–341. [Google Scholar] [CrossRef]
- Guidetti, P.; Terlizzi, A.; Fraschetti, S.; Boero, F. Spatio-Temporal Variability in Fish Assemblages Associated with Coralligenous Formations in South Eastern Apulia (SE Italy). Ital. J. Zool. 2002, 69, 325–331. [Google Scholar] [CrossRef]
- Tonin, S. Economic Value of Marine Biodiversity Improvement in Coralligenous Habitats. Ecol. Indic. 2018, 85, 1121–1132. [Google Scholar] [CrossRef]
- Martin, C.S.; Giannoulaki, M.; De Leo, F.; Scardi, M.; Salomidi, M.; Knittweis, L.; Pace, M.L.; Garofalo, G.; Gristina, M.; Ballesteros, E.; et al. Coralligenous and Maërl Habitats: Predictive Modelling to Identify Their Spatial Distributions across the Mediterranean Sea. Sci. Rep. 2014, 4, 5073. [Google Scholar] [CrossRef]
- Martin, S.; Charnoz, A.; Gattuso, J.-P. Photosynthesis, Respiration and Calcification in the Mediterranean Crustose Coralline Alga Lithophyllum cabiochae (Corallinales, Rhodophyta). Eur. J. Phycol. 2013, 48, 163–172. [Google Scholar] [CrossRef]
- Paoli, C.; Montefalcone, M.; Morri, C.; Vassallo, P.; Bianchi, C.N. Ecosystem Functions and Services of the Marine Animal Forests. In Marine Animal Forests; Springer: Cham, Switzerland, 2017; pp. 1271–1312. [Google Scholar]
- Chimienti, G.; Stithou, M.; Dalle Mura, I.; Mastrototaro, F.; D’Onghia, G.; Tursi, A.; Izzi, C.; Fraschetti, S. An Explorative Assessment of the Importance of Mediterranean Coralligenous Habitat to Local Economy: The Case of Recreational Diving. J. Environ. Account. Manag. 2017, 5, 315–325. [Google Scholar] [CrossRef]
- Mangos, A.; Basino, J.P.; Sauzade, D. Valeur Économique Des Bénéfices Soutenables Provenant Des Écosystèmes Marins Méditerranéens; Plan Bleu: Sophia Antipolis, France, 2010; p. 78. [Google Scholar]
- Kružić, P.; Rodić, P. Impact of Climate Changes on Coralligenous Community in the Adriatic Sea. In Proceedings of the 2nd Mediterranean Symposium on the Conservation of Coralligenous and Other Calcareous Bio-Concretions, Portorož, Slovenia, 29–30 October 2014; pp. 100–105. [Google Scholar]
- Piazzi, L.; Gennaro, P.; Balata, D. Threats to Macroalgal Coralligenous Assemblages in the Mediterranean Sea. Mar. Pollut. Bull. 2012, 64, 2623–2629. [Google Scholar] [CrossRef]
- Bevilacqua, S.; Guarnieri, G.; Farella, G.; Terlizzi, A.; Fraschetti, S. A Regional Assessment of Cumulative Impact Mapping on Mediterranean Coralligenous Outcrops. Sci. Rep. 2018, 8, 1757. [Google Scholar] [CrossRef]
- Terrón-Sigler, A.; León-Muez, D.; Peñalver-Duque, P.; Torre, F.E. The Effects of SCUBA Diving on the Endemic Mediterranean Coral Astroides Calycularis. Ocean Coast. Manag. 2016, 122, 1–8. [Google Scholar] [CrossRef]
- Rindi, F.; Braga, J.C.; Martin, S.; Peña, V.; Le Gall, L.; Caragnano, A.; Aguirre, J. Coralline Algae in a Changing Mediterranean Sea: How Can We Predict Their Future, If We Do Not Know Their Present? Front. Mar. Sci. 2019, 6, 723. [Google Scholar] [CrossRef]
- Lambeck, K.; Antonioli, F.; Purcell, A.; Silenzi, S. Sea-Level Change along the Italian Coast for the Past 10,000 Yr. Quat. Sci. Rev. 2004, 23, 1567–1598. [Google Scholar] [CrossRef]
- Canovas-Molina, A.; Montefalcone, M.; Vassallo, P.; Morri, C.; Bianchi, C.N.; Bavestrello, G. Combining Literature Review, Acoustic Mapping and in Situ Observations: An Overview of Coralligenous Assemblages in Liguria (NW Mediterranean Sea). Sci. Mar. 2016, 80, 7–16. [Google Scholar] [CrossRef]
- Pierdomenico, M.; Bonifazi, A.; Argenti, L.; Ingrassia, M.; Casalbore, D.; Aguzzi, L.; Viaggiu, E.; Le Foche, M.; Chiocci, F. Geomorphological Characterization, Spatial Distribution and Environmental Status Assessment of Coralligenous Reefs along the Latium Continental Shelf. Ecol. Indic. 2021, 131, 108219. [Google Scholar] [CrossRef]
- Falace, A.; Kaleb, S.; Curiel, D.; Miotti, C.; Galli, G.; Querin, S.; Ballesteros, E.; Solidoro, C.; Bandelj, V. Calcareous Bio-Concretions in the Northern Adriatic Sea: Habitat Types, Environmental Factors That Influence Habitat Distributions, and Predictive Modeling. PLoS ONE 2015, 10, e0140931. [Google Scholar] [CrossRef]
- Newton, R.S.; Stefanon, A. The “Tegnue de Ciosa” Area: Patch Reefs in the Northern Adriatic Sea. Mar. Geol. 1975, 19, M27–M33. [Google Scholar] [CrossRef]
- Spagnoli, F.; Dinelli, E.; Giordano, P.; Marcaccio, M.; Zaffagnini, F.; Frascari, F. Sedimentological, Biogeochemical and Mineralogical Facies of Northern and Central Western Adriatic Sea. J. Mar. Syst. 2014, 139, 183–203. [Google Scholar] [CrossRef]
- Curiel, D.; Falace, A.; Bandelj, V.; Kaleb, S.; Solidoro, C.; Ballesteros, E. Species Composition and Spatial Variability of Macroalgal Assemblages on Biogenic Reefs in the Northern Adriatic Sea. Bot. Mar. 2012, 55, 625–638. [Google Scholar] [CrossRef]
- Piazzi, L.; Turicchia, E.; Rindi, F.; Falace, A.; Gennaro, P.; Abbiati, M.; Bandelj, V.; Calcinai, B.; Ciriaco, S.; Costantini, F.; et al. NAMBER: A Biotic Index for Assessing the Ecological Quality of Mesophotic Biogenic Reefs in the Northern Adriatic Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 2023, 33, 298–311. [Google Scholar] [CrossRef]
- Bettoso, N.; Faresi, L.; Pitacco, V.; Orlando-Bonaca, M.; Aleffi, I.F.; Lipej, L. Species Richness of Benthic Macrofauna on Rocky Outcrops in the Adriatic Sea by Using Species-Area Relationship (SAR) Tools. Water 2023, 15, 318. [Google Scholar] [CrossRef]
- Guidetti, P.; Addis, P.; Atzori, F.; Bussotti, S.; Calò, A.; Cau, A.; Culioli, J.-M.; De Lucia, G.; Di Franco, A.; Di Lorenzo, M.; et al. Assessing the Potential of Marine Natura 2000 Sites to Produce Ecosystem-Wide Effects in Rocky Reefs: A Case Study from Sardinia Island (Italy). Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 537–545. [Google Scholar] [CrossRef]
- Meinesz, A.; Blanfuné, A. 1983–2013: Development of Marine Protected Areas along the French Mediterranean Coasts and Perspectives for Achievement of the Aichi Target. Mar. Policy 2015, 54, 10–16. [Google Scholar] [CrossRef]
- Furlan, E.; Torresan, S.; Critto, A.; Lovato, T.; Solidoro, C.; Lazzari, P.; Marcomini, A. Cumulative Impact Index for the Adriatic Sea: Accounting for Interactions among Climate and Anthropogenic Pressures. Sci. Total Environ. 2019, 670, 379–397. [Google Scholar] [CrossRef]
- Giani, M.; Djakovac, T.; Degobbis, D.; Cozzi, S.; Solidoro, C.; Umani, S.F. Recent Changes in the Marine Ecosystems of the Northern Adriatic Sea. Estuar. Coast. Shelf Sci. 2012, 115, 1–13. [Google Scholar] [CrossRef]
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; Nykjaer, L.; Rosenberg, A.A. Cumulative Human Impacts on Mediterranean and Black Sea Marine Ecosystems: Assessing Current Pressures and Opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef] [PubMed]
- Stirn, J.; Avcin, A.; Cencekj, J.; Dorer, M.; Gomiscek, S.; Kvender, S.; Malej, A.; Meischner, D.; Nozina, I.; Paul, J.; et al. Pollution Problems of the Adriatic Sea: An Interdisciplinary Approach. Rev. Intern. Océanogr. Méd. 1974, 35, 21–78. [Google Scholar]
- Acquavita, A.; Covelli, S.; Emili, A.; Berto, D.; Faganeli, J.; Giani, M.; Horvat, M.; Koron, N.; Rampazzo, F. Mercury in the Sediments of the Marano and Grado Lagoon (Northern Adriatic Sea): Sources, Distribution and Speciation. Estuar. Coast. Shelf Sci. 2012, 113, 20–31. [Google Scholar] [CrossRef]
- Carić, H.; Mackelworth, P. Cruise Tourism Environmental Impacts–The Perspective from the Adriatic Sea. Ocean Coast. Manag. 2014, 102, 350–363. [Google Scholar] [CrossRef]
- Coll, M.; Santojanni, A.; Palomera, I.; Tudela, S.; Arneri, E. An Ecological Model of the Northern and Central Adriatic Sea: Analysis of Ecosystem Structure and Fishing Impacts. J. Mar. Syst. 2007, 67, 119–154. [Google Scholar] [CrossRef]
- Covelli, S.; Faganeli, J.; Horvat, M.; Brambati, A. Mercury Contamination of Coastal Sediments as the Result of Long-Term Cinnabar Mining Activity (Gulf of Trieste, Northern Adriatic Sea). Appl. Geochem. 2001, 16, 541–558. [Google Scholar] [CrossRef]
- Gissi, E.; Menegon, S.; Sarretta, A.; Appiotti, F.; Maragno, D.; Vianello, A.; Depellegrin, D.; Venier, C.; Barbanti, A. Addressing Uncertainty in Modelling Cumulative Impacts within Maritime Spatial Planning in the Adriatic and Ionian Region. PLoS ONE 2017, 12, e0180501. [Google Scholar] [CrossRef] [PubMed]
- Micheli, F.; Levin, N.; Giakoumi, S.; Katsanevakis, S.; Abdulla, A.; Coll, M.; Fraschetti, S.; Kark, S.; Koutsoubas, D.; Mackelworth, P.; et al. Setting Priorities for Regional Conservation Planning in the Mediterranean Sea. PLoS ONE 2013, 8, e59038. [Google Scholar] [CrossRef]
- Torresan, S.; Critto, A.; Rizzi, J.; Marcomini, A. Assessment of Coastal Vulnerability to Climate Change Hazards at the Regional Scale: The Case Study of the North Adriatic Sea. Nat. Hazards Earth Syst. Sci. 2012, 12, 2347–2368. [Google Scholar] [CrossRef]
- Bandelj, V.; Solidoro, C.; Laurent, C.; Querin, S.; Kaleb, S.; Gianni, F.; Falace, A. Cross-Scale Connectivity of Macrobenthic Communities in a Patchy Network of Habitats: The Mesophotic Biogenic Habitats of the Northern Adriatic Sea. Estuar. Coast. Shelf Sci. 2020, 245, 106978. [Google Scholar] [CrossRef]
- Ponti, M.; Fava, F.; Abbiati, M. Spatial–Temporal Variability of Epibenthic Assemblages on Subtidal Biogenic Reefs in the Northern Adriatic Sea. Mar. Biol. 2011, 158, 1447–1459. [Google Scholar] [CrossRef]
- Trobec, A.; Busetti, M.; Zgur, F.; Baradello, L.; Babich, A.; Cova, A.; Gordini, E.; Romeo, R.; Tomini, I.; Poglajen, S.; et al. Thickness of Marine Holocene Sediment in the Gulf of Trieste (Northern Adriatic Sea). Earth Syst. Sci. Data 2018, 10, 1077–1092. [Google Scholar] [CrossRef]
- Regione Autonoma Friuli Venezia Giulia DGR 1701/2019 Del 04 Ottobre 2019—LR 8/2007, Art. 10. Misure Di Conservazione Dei Siti Marini Del Friuli Venezia Giulia. 2019. Available online: https://mtom.regione.fvg.it (accessed on 25 February 2025).
- Casellato, S.; Sichirollo, E.; Cristofoli, A.; Masiero, L.; Soresi, S. Biodiversity of some rocky outcrops in the gulf of Venice (“Tegnue”). Biol. Mar. Medit. 2005, 12, 69–77. [Google Scholar]
- Bianchi, C.N.; Azzola, A.; Cocito, S.; Morri, C.; Oprandi, A.; Peirano, A.; Sgorbini, S.; Montefalcone, M. Biodiversity Monitoring in Mediterranean Marine Protected Areas: Scientific and Methodological Challenges. Diversity 2022, 14, 43. [Google Scholar] [CrossRef]
- García-Gómez, J.C.; González, A.R.; Maestre, M.J.; Espinosa, F. Detect Coastal Disturbances and Climate Change Effects in Coralligenous Community through Sentinel Stations. PLoS ONE 2020, 15, e0231641. [Google Scholar] [CrossRef]
- Dethier, M.N.; Graham, E.S.; Cohen, S.; Tear, L.M. Visual versus Random-Point Percent Cover Estimations: “objective” Is Not Always Better. Mar. Ecol. Prog. Ser. 1993, 96, 93–100. [Google Scholar] [CrossRef]
- Anderson, M.J. A New Method for Non-Parametric Multivariate Analysis of Variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-Based Tests for Homogeneity of Multivariate Dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; De Caceres, M. Beta Diversity as the Variance of Community Data: Dissimilarity Coefficients and Partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2015. [Google Scholar]
- Anderson, M.; Gorley, R.N.; Clarke, K. PERMANOVA+ for Primer: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008; Available online: https://learninghub.primer-e.com/books (accessed on 25 February 2025).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024. [Google Scholar]
- Sandrini-Neto, L.; Gilbert, E.; Camargo, M. GAD: Analysis of Variance from General Principles. 2024. Available online: https://CRAN.R-project.org/package=GAD (accessed on 25 February 2025).
- Dry, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Classe, S.; Guernard, G.; Jombardt, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. Adespatial: Multivariate Multiscale Spatial Analysis. 2025. Available online: https://CRAN.R-project.org/package=adespatial (accessed on 25 February 2025).
- Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 Establishing a Framework for Community Action in the Field of Marine Environmental Policy (Marine Strategy Framework Directive) (Text with EEA Relevance). Available online: https://www.bsbd.bg/UserFiles/Dir2008-56-EO-EN.pdf (accessed on 25 February 2025).
- Gianni, F.; Turicchia, E.; Abbiati, M.; Calcinai, B.; Caragnano, A.; Ciriaco, S.; Costantini, F.; Kaleb, S.; Piazzi, L.; Puce, S.; et al. Spatial Patterns and Drivers of Benthic Community Structure on the Northern Adriatic Biogenic Reefs. Biodivers. Conserv. 2023, 32, 3283–3306. [Google Scholar] [CrossRef]
- Krauss, J.; Bommarco, R.; Guardiola, M.; Heikkinen, R.K.; Helm, A.; Kuussaari, M.; Lindborg, R.; Ockinger, E.; Partel, M.; Pino, J.; et al. Habitat Fragmentation Causes Immediate and Timedelayed Biodiversity Loss at Different Trophic Levels. Ecol. Lett. 2010, 13, 597–605. [Google Scholar] [CrossRef]
- Gera, A.; Pages, J.F.; Romero, J.; Alcoverro, T. Combined Effects of Fragmentation and Herbivory on Posidonia oceanica Seagrass Ecosystems. J. Ecol. 2013, 101, 1053–1061. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Piccardo, M.; Vellani, V.; Anselmi, S.; Grazioli, E.; Renzi, M.; Terlizzi, A.; Pittura, L.; D’Errico, G.; Regoli, F.; Bevilacqua, S. The Application of the Weight-Of-Evidence Approach for an Integrated Ecological Risk Assessment of Marine Protected Sites. Ecol. Indic. 2024, 159, 111676. [Google Scholar] [CrossRef]
- Russo, E.; Monti, M.A.; Mangano, M.C.; Raffaetà, A.; Sarà, G.; Silvestri, C.; Pranovi, F. Temporal and Spatial Patterns of Trawl Fishing Activities in the Adriatic Sea (Central Mediterranean Sea, GSA17). Ocean Coast. Manag. 2020, 192, 105231. [Google Scholar] [CrossRef]
- Personnic, S.; Boudouresque, C.-F.; Astruch, P.; Ballesteros, E.; Blouet, S.; Bellan-Santini, D.; Bonhomme, P.; Thibault-Botha, D.; Feunteun, E.; Harmelin-Vivien, M.; et al. An Ecosystem-Based Approach to Assess the Status of a Mediterranean Ecosystem, the Posidonia oceanica Seagrass Meadow. PLoS ONE 2014, 9, e98994. [Google Scholar] [CrossRef] [PubMed]
- Astruch, P.; Orts, A.; Schohn, T.; Belloni, B.; Ballesteros, E.; Bănaru, D.; Bianchi, C.N.; Boudouresque, C.-F.; Changeux, T.; Chevaldonné, P.; et al. Ecosystem-based assessment of a widespread Mediterranean marine habitat: The Coastal Detrital Bottoms, with a special focus on epibenthic assemblages. Front. Mar. Sci. 2023, 10, 1130540. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.-F.; Personnic, S.; Ruitton, S.; Ballesteros, E.; Bellan-Santini, D.; Bianchi, C.N.; Bussotti, S.; Cebrian, E.; et al. An ecosystem-based approach to assess the status of Mediterranean algae-dominated shallow rocky reefs. Mar. Pollut. Bull. 2017, 117, 311–329. [Google Scholar] [CrossRef]
- Ingrosso, G.; Abbiati, M.; Badalamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Nike Bianchi, C.; et al. Chapter Three—Mediterranean Bioconstructions Along the Italian Coast. Adv. Mar. Biol. 2018, 79, 61–136. [Google Scholar] [CrossRef] [PubMed]
- Turicchia, E.; Abbiati, M.; Bettuzzi, M.; Calcinai, B.; Morigi, M.P.; Summers, A.P.; Ponti, M. Bioconstruction and Bioerosion in the Northern Adriatic Coralligenous Reefs Quantified by X-Ray Computed Tomography. Front. Mar. Sci. 2022, 8, 790869. [Google Scholar] [CrossRef]
- Vitelletti, M.L.; Manea, E.; Bongiorni, L.; Ricchi, A.; Sangelantoni, L.; Bonaldo, D. Modelling Distribution and Fate of Coralligenous Habitat in the Northern Adriatic Sea under a Severe Climate Change Scenario. Front. Mar. Sci. 2023, 10, 1050293. [Google Scholar] [CrossRef]
- Bertolino, M.; Costa, G.; Carella, M.; Cattaneo-Vietti, R.; Cerrano, C.; Pansini, M.; Quarta, G.; Calcagnile, L.; Bavestrello, G. The Dynamics of a Mediterranean Coralligenous Sponge Assemblage at Decennial and Millennial Temporal Scales. PLoS ONE 2017, 12, e0177945. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | d.f. | SS | MS | Pseudo-F | p (Perm) |
|---|---|---|---|---|---|
| Ti | 5 | 2.57 × 105 | 5.15 × 104 | ||
| Ar | 1 | 1.99 × 105 | 1.99 × 105 | ||
| Ti × Ar | 5 | 8.88 × 104 | 1.78 × 104 | 2.912 | 0.0001 |
| Si(Ar) | 4 | 3.01 × 104 | 7.70 × 103 | ||
| St(Si(Ar)) | 12 | 3.85 × 104 | 3.21 × 103 | ||
| Ti × Si(Ar) | 20 | 1.22 × 105 | 6.10 × 103 | 1.8127 | 0.0001 |
| Ti × St(Si(Ar)) | 60 | 2.02 × 105 | 3.37 × 103 | 1.5176 | 0.0001 |
| Res | 1067 | 2.37 × 106 | 2.22 × 103 |
| N. of Taxa | Total Coverage | ||||
|---|---|---|---|---|---|
| Source of Variation | d.f. | MS | F | MS | F |
| Ti | 5 | 348.20 | 1.15 × 104 | 19.143 *** | |
| Ar | 1 | 180.10 | 31.00 | 0.167 NS | |
| Ti × Ar | 12 | 138.60 | 7.657 *** | 1.4 × 103 | 2.330 NS |
| Si(Ar) | 4 | 20.70 | 185.00 | ||
| St(Si(Ar)) | 5 | 7.10 | 1.392 NS | 274.00 | 1.593 NS |
| Ti × Si(Ar) | 20 | 18.10 *** | 3.549 *** | 603.00 | 3.506 *** |
| Ti × St(Si(Ar)) | 60 | 5.10 NS | 1.085 NS | 172.00 | 1.023 NS |
| Res | 972 | 4.70 | 168.00 | ||
| Cochran’s C-test | C = 0.025 NS | C = 0.049 *** | |||
| Transformation | None | None | |||
| Turf | F | CBB | F | Tedania spp. | F | YLS | F | ||
|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | d.f. | MS | MS | MS | MS | ||||
| Ti | 5 | 1.23 × 104 | 10.01 | 20.78 | 18.20 | ||||
| Ar | 1 | 6.12 × 103 | 45.76 | 2.06 | 1.13 × 103 | ||||
| Ti × Ar | 12 | 1.34 × 103 | 5.711 *** | 2.77 | 2.856 * | 12.99 | 5.959 *** | 26.60 | 5.541 *** |
| Si(Ar) | 4 | 245.00 | 1.042 NS | 2.82 | 2.31 | 1.059 NS | 1.70 | 0.354 NS | |
| St(Si(Ar)) | 5 | 95.00 | 0.28 | 1.44 | 5.10 | ||||
| Ti × Si(Ar) | 20 | 235.00 | 1.078 NS | 0.97 | 5.389 *** | 2.18 | 1.246 NS | 4.80 | 1.000 NS |
| Ti × St(Si(Ar)) | 60 | 218.00 | 2.564 *** | 0.18 | 1.500 ** | 1.75 | 1.635 *** | 4.80 | 1.714 *** |
| Res | 972 | 85.00 | 0.12 | 1.07 | 2.8 | ||||
| Cochran’s C-test | C = 0.048 *** | C = 0.029 NS | C = 0.058 *** | C = 0.029 NS | |||||
| Transformation | None | Square root | None | Square root | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motta, G.; Vellani, V.; Piccardo, M.; De Luca, M.; Ciriaco, S.; Segarich, M.; Peratoner, L.; Spoto, M.; Terlizzi, A.; Renzi, M.; et al. Monitoring the Status of Mesophotic Biogenic Reefs in the Northern Adriatic Sea: Comparing a Biotic Index and Multivariate Community Patterns. Environments 2025, 12, 124. https://doi.org/10.3390/environments12040124
Motta G, Vellani V, Piccardo M, De Luca M, Ciriaco S, Segarich M, Peratoner L, Spoto M, Terlizzi A, Renzi M, et al. Monitoring the Status of Mesophotic Biogenic Reefs in the Northern Adriatic Sea: Comparing a Biotic Index and Multivariate Community Patterns. Environments. 2025; 12(4):124. https://doi.org/10.3390/environments12040124
Chicago/Turabian StyleMotta, Gregorio, Verdiana Vellani, Manuela Piccardo, Matteo De Luca, Saul Ciriaco, Marco Segarich, Lisa Peratoner, Maurizio Spoto, Antonio Terlizzi, Monia Renzi, and et al. 2025. "Monitoring the Status of Mesophotic Biogenic Reefs in the Northern Adriatic Sea: Comparing a Biotic Index and Multivariate Community Patterns" Environments 12, no. 4: 124. https://doi.org/10.3390/environments12040124
APA StyleMotta, G., Vellani, V., Piccardo, M., De Luca, M., Ciriaco, S., Segarich, M., Peratoner, L., Spoto, M., Terlizzi, A., Renzi, M., & Bevilacqua, S. (2025). Monitoring the Status of Mesophotic Biogenic Reefs in the Northern Adriatic Sea: Comparing a Biotic Index and Multivariate Community Patterns. Environments, 12(4), 124. https://doi.org/10.3390/environments12040124