Using Pupillometry to Assess the Atypical Pupillary Light Reflex and LC-NE System in ASD


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Models of Neuropathology-Functional & Anatomical Differences Underlying ASD
3. Underconnectivity & Inefficient Neural Processing
4. Valproic Acid (VPA) Animal Model of ASD and the Visual Neural Pathway
5. Brainstem Circuits Influencing Maladaptive Behavior and Language Acquisition in ASD
6. Eye-Tracking Methodology & Pupillometry to Measure Functional and Anatomical Differences
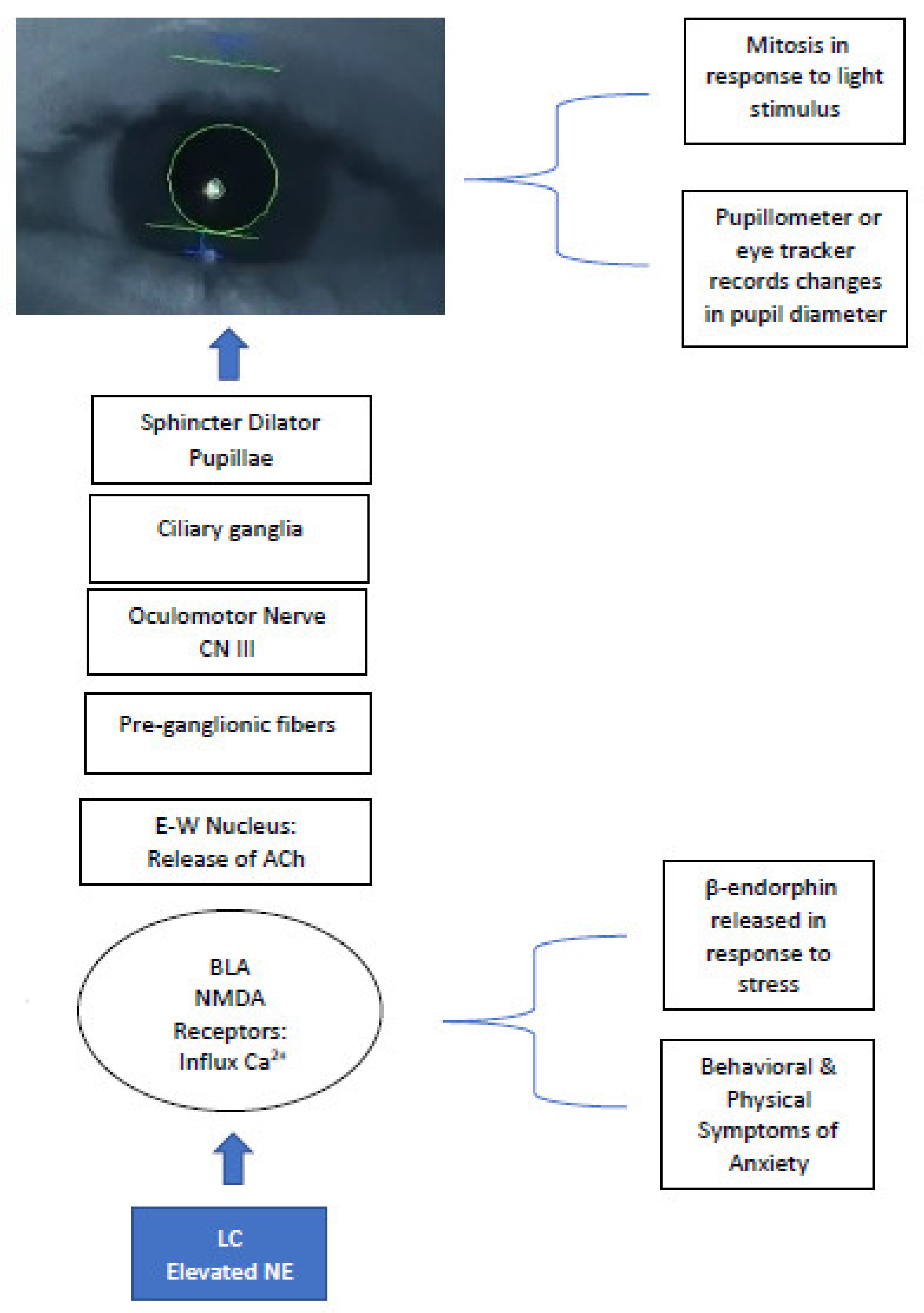
7. Pupillometry to Measure ANS Function in ASD & LC-NE Activity
8. Pupil Response as a Useful Biomarker for Assessing Brain Function in ASD
9. Phasic Hyperarousal and the “ASD Advantage” in Visual Tracking
10. Pupillary Response, Modulation of the LC-NE System, and Elevated Arousal in ASD
11. The Atypical PLR Observed in ASD—Valid Measures of the PLR
12. PLR and Corollaries of ANS Modulation within ASD
13. PLR as an Index of Maturation of the Visual Pathway
14. Discussion and Future Directions
Supplementary Materials
Funding
Conflicts of Interest
References
- Kanner, L. Autistic Disturbances of Affective Contact; 1943; pp. 217–250, [Publisher not identified]. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th ed.; American Psychiatric Association Publishing: Washington, DC, USA, 2013. [Google Scholar]
- Dawson, G.; Jones, E.J.; Merkle, K.; Venema, K.; Lowy, R.; Faja, S.; Webb, S.J. Early behavioral intervention is associated with normalized brain activity in young children with autism. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Huttenlocher, P.R.; Dabholkar, A.S. Regional differences in synaptogenesis in human cerebral cortex. J. Comp. Neurol. 1997, 387, 167–178. [Google Scholar] [CrossRef]
- Banerjee, S.; Riordan, M.; Bhat, M.A. Genetic aspects of autism spectrum disorders: Insights from animal models. Front. Cell. Neurosci. 2014, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Politte, L.C.; Henry, C.A.; McDougle, C.J. Psychopharmacological interventions in autism spectrum disorder. Harvard Rev. Psychiatry 2014, 22, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Turi, M.; Burr, D.C.; Binda, P. Pupillometry reveals perceptual differences that are tightly linked to autistic traits in typical adults. eLife 2018, 7, e32399. [Google Scholar] [CrossRef] [PubMed]
- Ebitz, R.B.; Moore, T. Selective modulation of the pupil light reflex by prefrontal cortex microstimulation. J. Neurosci. 2017, 2433-16. [Google Scholar] [CrossRef] [PubMed]
- Hess, E.H.; Polt, J.M. Pupil size as related to interest value of visual stimuli. Science 1960, 132, 349–350. [Google Scholar] [CrossRef] [PubMed]
- Kahneman, D.; Beatty, J. Pupil diameter and load on memory. Science 1966, 154, 1583–1585. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.F. The neuropathology of autism. Brain Pathol. 2007, 17, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; Pierce, K.; Schumann, C.M.; Redcay, E.; Buckwalter, J.A.; Kennedy, D.P.; Morgan, J. Mapping early brain development in autism. Neuron 2007, 56, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; Pierce, D. Brain overgrowth in autism during a critical time in development: Implications for frontal pyramidal neuron and interneuron development and connectivity. Int. J. Dev. Neurosci. 2005, 23, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Grzadzinski, R.; Huerta, M.; Lord, C. DSM-5 and autism spectrum disorders (ASDs): An opportunity for identifying ASD subtypes. Mol. Autism 2013, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Tager-Flusberg, H.; Joseph, R.M. Identifying neurocognitive phenotypes in autism. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Gerdts, J.; Bernier, R. The broader autism phenotype and its implications on the etiology and treatment of autism spectrum disorder. Autism Res. Treat. 2011, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Piven, J.; Palmer, P.; Jacobi, D.; Childress, D.; Arndt, S. Broader autism phenotype: Evidence from a family history study of multiple-incidence autism families. Am. J. Psychiatry 1997, 154, 185–190. [Google Scholar] [PubMed]
- Lainhart, J.E.; Ozonoff, S.; Coon, H.; Krasny, L.; Dinh, E.; Nice, J.; McMahon, W. Autism, regression, and the broader autism phenotype. Am. J. Med. Genet. 2002, 113, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Lainhart, J.E.; Bigler, E.D.; Bocian, M.; Coon, H.; Dinh, E.; Dawson, G.; Volkmar, F. Head circumference and height in autism: A study by the collaborative program of excellence in autism. Am. J. Med. Genet. Part A 2006, 140, 2257–2274. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; Campbell, K.; Solso, S. Brain growth across the life span in autism: Age-specific changes in anatomical pathology. Brain Res. 2011, 1380, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Toal, F.; Daly, E.M.; Page, L.; Deeley, Q.; Hallahan, B.; Bloemen, O.; Murphy, D.G.M. Clinical and anatomical heterogeneity in autistic spectrum disorder: A structural MRI study. Psychol. Med. 2010, 40, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Amaral, D.; Schumann, C.; Nordahl, C. Neuroanatomy of autism. Trends Neurosci. 2008, 31, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Pickett, J.; London, E. The neuropathology of autism: A review. J. Neuropathol. Exp. Neurol. 2005, 64, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; Karns, C.M.; Davis, H.R.; Ziccardi, R.; Carper, R.A.; Tigue, Z.D.; Lincoln, A.J. Unusual brain growth patterns in early life in patients with autistic disorder an MRI study. Neurology 2001, 57, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Schumann, C.M.; Bloss, C.S.; Barnes, C.C.; Wideman, G.M.; Carper, R.A.; Akshoomoff, N.; Courchesne, E. Longitudinal magnetic resonance imaging study of cortical development through early childhood in autism. J. Neurosci. 2010, 30, 4419–4427. [Google Scholar] [CrossRef] [PubMed]
- Nordahl, C.W.; Lange, N.; Li, D.D.; Barnett, L.A.; Lee, A.; Buonocore, M.H.; Simon, T.; Amaral, D. Brain enlargement is associated with regression in preschool age boys with autism spectrum disorder. Proc. Natl. Acad. Sci. USA 2011, 108, 20195–20200. [Google Scholar] [CrossRef] [PubMed]
- Koshino, H.; Carpenter, P.A.; Minshew, N.J.; Cherkassky, V.L.; Keller, T.A.; Just, M.A. Functional connectivity in an fMRI working memory task in high-functioning autism. Neuroimage 2005, 24, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Herbert, M.R.; Ziegler, D.A.; Deutsch, C.K.; O’brien, L.M.; Lange, N.; Bakardjiev, A.; Caviness, V.J. Dissociations of cerebral cortex, subcortical and cerebral white matter volumes in autistic boys. Brain 2003, 126, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Just, M.A.; Cherkassky, V.L.; Keller, T.A.; Minshew, N.J. Cortical activation and synchronization during sentence comprehension in high-functioning autism: Evidence of underconnectivity. Brain 2004, 127, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Kleinhans, N.M.; Richards, T.; Sterling, L.; Stegbauer, K.C.; Mahurin, R.; Johnson, L.C.; Aylward, E. Abnormal functional connectivity in autism spectrum disorders during face processing. Brain 2008, 131, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Kana, R.K.; Keller, T.A.; Minshew, N.J.; Just, M.A. Inhibitory control in high-functioning autism: Decreased activation and underconnectivity in inhibition networks. Biol. Psychiatry 2007, 62, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Buxhoeveden, D.P.; Casanova, M.F. The minicolumn hypothesis in neuroscience. Brain 2002, 125, 935–951. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.F.; Buxhoeveden, D.P.; Switala, A.E.; Roy, E. Minicolumnar pathology in autism. Neurology 2002, 58, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.F.; van Kooten, I.A.; Switala, A.E.; van Engeland, H.; Heinsen, H.; Steinbusch, H.W.; Schmitz, C. Minicolumnar abnormalities in autism. Acta Neuropathol. 2006, 112, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Adolphs, R.; Sears, L.; Piven, J. Abnormal processing of social information from faces in autism. J. Cogn. Neurosci. 2001, 13, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Baron-Cohen, S.; Ring, H.A.; Bullmore, E.T.; Wheelwright, S.; Ashwin, C.; Williams, S.C.R. The amygdala theory of autism. Neurosci. Biobehav. Rev. 2000, 24, 355–364. [Google Scholar] [CrossRef]
- Dawson, G. Early behavioral intervention, brain plasticity, and the prevention of autism spectrum disorder. Dev. Psychopathol. 2008, 20, 775–803. [Google Scholar] [CrossRef] [PubMed]
- Dawson, G.; Rogers, S.; Munson, J.; Smith, M.; Winter, J.; Greenson, J.; Varley, J. Randomized, controlled trial of an intervention for toddlers with autism: The early start denver model. Pediatrics 2010, 125, e17–e23. [Google Scholar] [CrossRef] [PubMed]
- Marmolejo, N.; Paez, J.; Levitt, J.B.; Jones, L.B. Early postnatal lesion of the medial dorsal nucleus leads to loss of dendrites and spines in adult prefrontal cortex. Dev. Neurosci. 2012, 34, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Gogolla, N.; LeBlanc, J.J.; Quast, K.B.; Südhof, T.C.; Fagiolini, M.; Hensch, T.K. Common circuit defect of excitatory-inhibitory balance in mouse models of autism. J. Neurodev. Dis. 2009, 1, 172. [Google Scholar] [CrossRef] [PubMed]
- Rakic, P. Mode of cell migration to the superficial layers of fetal monkey neocortex. J. Comp. Neurol. 1972, 145, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Bernier, R.; Golzio, C.; Xiong, B.; Stessman, H.A.; Coe, B.P.; Penn, O.; Eichler, E.E. Disruptive CHD8 mutations define a subtype of autism early in development. Cell 2014, 158, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.; Klin, A. Attention to eyes is present but in decline in 2-6-month-old infants later diagnosed with autism. Nature 2013, 504, 427. [Google Scholar] [CrossRef] [PubMed]
- Rodier, P.M.; Ingram, J.L.; Tisdale, B.; Nelson, S.; Romano, J. Embryological origin for autism: Developmental anomalies of the cranial nerve motor nuclei. J. Comp. Neurol. 1996, 370, 247–261. [Google Scholar] [CrossRef]
- Schneider, T.; Przewłocki, R. Behavioral alterations in rats prenatally exposed to valproic acid: Animal model of autism. Neuropsychopharm 2005, 30, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Roullet, F.I.; Lai, J.K.; Foster, J.A. In utero exposure to valproic acid and autism—A current review of clinical and animal studies. Neurotoxicol. Teratol. 2013, 36, 47–56. [Google Scholar] [CrossRef] [PubMed]
- United States Food and Drug Administration. FDA Drug Safety Communication: Children Born to Mothers Who Took Valproate Products While Pregnant May Have Impaired Cognitive Development; FDA: Silver Spring, MD, USA. Available online: https://www.fda.gov/Drugs/DrugSafety/ucm261543.htm (accessed on 11 August 2018).
- Tager-Flusberg, H.; Paul, R.; Lord, C. Language and communication in autism. Handb. Autism Pervasive Dev. Disord. 2005, 1, 335–364. [Google Scholar]
- Paul, R. Language Disorders from Infancy through Adolescence: Assessment & Intervention, 3rd ed.; Mosby Elsevier: St. Louis, MO, USA, 2007; ISBN 13 978-0-323-03685-6. [Google Scholar]
- Klin, A.; Jones, W.; Schultz, R.; Volkmar, F.; Cohen, D. Visual fixation patterns during viewing of naturalistic social situations as predictors of social competence in individuals with autism. Arch. Gen. Psychiatry 2002, 59, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.; Carr, K.; Klin, A. Absence of preferential looking to the eyes of approaching adults predicts level of social disability in 2-year-old toddlers with autism spectrum disorder. Arch. Gen. Psychiatry 2008, 65, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Baron-Cohen, S.; Ring, H.A.; Wheelwright, S.; Bullmore, E.T.; Brammer, M.J.; Simmons, A.; Williams, S.C. Social intelligence in the normal and autistic brain: An fMRI study. Eur. J. Neurosci. 1999, 11, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Adolphs, R.; Tranel, D.; Damasio, H.; Damasio, A.R. Fear and the human amygdala. J. Neurosci. 1995, 15, 5879–5891. [Google Scholar] [CrossRef] [PubMed]
- Mobbs, D.; Marchant, J.L.; Hassabis, D.; Seymour, B.; Tan, G.; Gray, M.; Frith, C.D. From threat to fear: The neural organization of defensive fear systems in humans. J. Neurosci. 2009, 29, 12236–12243. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.E.; Iwata, J.; Cicchetti, P.R.D.J.; Reis, D.J. Different projections of the central amygdaloid nucleus mediate autonomic and behavioral correlates of conditioned fear. J. Neurosci. 1988, 8, 2517–2529. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.M.; Nacewicz, B.M.; Johnstone, T.; Schaefer, H.S.; Gernsbacher, M.S.; Goldsmith, H.H.; Davidson, R.J. Gaze fixation and the neural circuitry of face processing in autism. Nat. Neurosci. 2005, 8, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Adolphs, R. Fear, faces, and the human amygdala. Curr. Opin. Neurobiol. 2008, 18, 166–172. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.C.; Webb, S.J.; Keehn, B.; Dawson, G. Patterns of visual attention to faces and objects in autism spectrum disorder. J. Autism Dev. Disord. 2011, 41, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J.; Sacks, D.S.; Crepeau, L.; Abbott, B.B. The psycho- and neuro-biology of fear systems in the brain. In Aversive Events and Behavior; Denny, M.R., Ed.; Lawrence Erlbaum Assocs. Inc. Publisher: New York, NY, USA, 1991; pp. 7–59. [Google Scholar]
- Neuhaus, E.; Bernier, R.; Beauchaine, T.P. Brief report: Social skills, internalizing and externalizing symptoms, and respiratory sinus arrhythmia in autism. J. Autism Dev. Disord. 2014, 44, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Boraston, Z.; Blakemore, S.J. The application of eye-tracking technology in the study of autism. J. Physiol. 2007, 581, 893–898. [Google Scholar] [CrossRef] [PubMed]
- Tottenham, N.; Hertzig, M.E.; Gillespie-Lynch, K.; Gilhooly, T.; Millner, A.J.; Casey, B.J. Elevated amygdala response to faces and gaze aversion in autism spectrum disorder. Soc. Cogn. Affect. Neurosci. 2014, 9, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Vivanti, G.; Dissanayake, C. Propensity to imitate in autism is not modulated by the model’s gaze direction: An eye-tracking study. Autism Res. 2014, 7, 392–399. [Google Scholar] [CrossRef] [PubMed]
- de Gelder, B. Towards the neurobiology of emotional body language. Nature 2006, 7, 242–249. [Google Scholar] [CrossRef]
- Steinhauer, S.R.; Siegle, G.J.; Condray, R.; Pless, M. Sympathetic and parasympathetic innervation of pupillary dilation during sustained processing. Int. J. Psychophysiol. 2004, 52, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Laeng, B.; Sirois, S.; Gredebäck, G. Pupillometry a window to the preconscious? Perspect. Psychol. Sci. 2012, 7, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Karatekin, C. Development of attentional allocation in the dual task paradigm. Int. J. Psychophysiol. 2004, 52, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Krieger, D.; Adams, H.P.; Schwarz, S.; Rieke, K.; Aschoff, A.; Hacke, W. Prognostic and clinical relevance of pupillary responses, intracranial pressure monitoring, and brainstem auditory evoked potentials in comatose patients with acute supratentorial mass lesions. Crit. Care Med. 1993, 21, 1944–1950. [Google Scholar] [CrossRef] [PubMed]
- Kerr, F.W.; Hollowell, O.W. Location of pupillomotor and accommodation fibres in the oculomotor nerve: Experimental observations on paralytic mydriasis. J. Neurol. Neurosurg. Psychiatry 1964, 27, 473. [Google Scholar] [CrossRef] [PubMed]
- Lowenstein, O.; Lowenfield, I.E. Role of sympathetic and parasympathetic systems in reflex dilation of the pupil. Arch. Neurol. Psychiatry 1950, 64, 314–340. [Google Scholar] [CrossRef]
- Neuroptics, Inc. Neuroptics 3000© Handheld Pupillometer; Neuroptics, Inc.: Laguna Hills, CA, USA, 2018; Available online: https://neuroptics.com/plr-3000-hand-held/ (accessed on 1 September 2018).
- Hernandez, N.; Metzger, A.; Magne, R.; Bonnet-Brilhault, F.; Roux, S.; Barthelemy, C.; Martineau, J. Exploration of core features of a human face by healthy and autistic adults analyzed by visual scanning. Neuropsychologia 2009, 47, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Pelphrey, K.A.; Sasson, N.J.; Reznick, J.S.; Paul, G.; Goldman, B.D.; Piven, J. Visual scanning of faces in autism. J. Autism Dev. Disord. 2002, 32, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Riby, D.M.; Hancock, P.J.B. Do faces capture the attention of individuals with Williams Syndrome or autism? Evidence from tracking eye movements. J. Autism Dev. Disord. 2009, 39, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Pierce, K.; Conant, D.; Hazin, R.; Stoner, R.; Desmond, J. Preference for geometric patterns early in life as a risk factor for autism. Arch. Gen. Psychiatry 2011, 68, 101–109. [Google Scholar] [CrossRef] [PubMed]
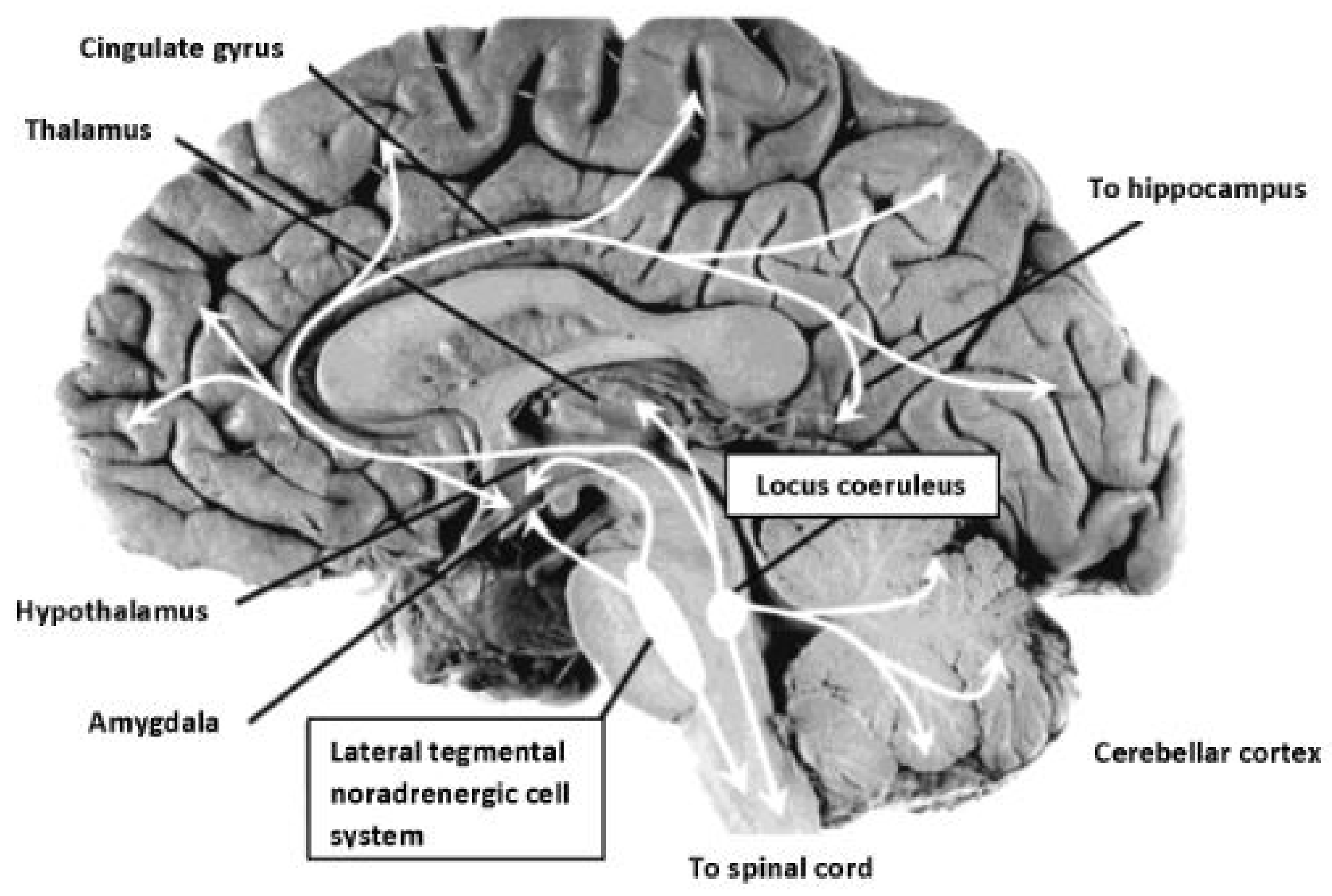
- Mehler, M.F.; Purpura, D.P. Autism, fever, epigenetics and the locus coeruleus. Brain Res. Rev. 2009, 59, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Berridge, C.W.; Waterhouse, B.D. The locus coeruleus–noradrenergic system: Modulation of behavioral state and state-dependent cognitive processes. Brain Res. Rev. 2003, 42, 33–84. [Google Scholar] [CrossRef]
- Kim, M.A.; Lee, H.S.; Lee, B.Y.; Waterhouse, B.D. Reciprocal connections between subdivisions of the dorsal raphe and the nuclear core of the locus coeruleus in the rat. Brain Res. 2004, 1026, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Samuels, E.R.; Szabadi, E. Functional neuroanatomy of the noradrenergic locus coeruleus: Its roles in the regulation of arousal and autonomic function part I: Principles of functional organisation. Curr. Neuropharmacol. 2008, 6, 254–285. [Google Scholar] [CrossRef] [PubMed]
- Southwick, S.M.; Bremner, J.D.; Rasmusson, A.; Morgan, C.A.; Arnsten, A.; Charney, D.S. Role of norepinephrine in the pathophysiology and treatment of posttraumatic stress disorder. Biol. Psychiatry 1999, 46, 1192–1204. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Cohen, J.D. An integrative theory of locus coeruleus-norepinephrine function: Adaptive gain and optimal performance. Annu. Rev. Neurosci. 2005, 28, 403–450. [Google Scholar] [CrossRef] [PubMed]
- Gilzenrat, M.S.; Nieuwenhuis, S.; Jepma, M.; Cohen, J.D. Pupil diameter tracks changes in control state predicted by the adaptive gain theory of locus coeruleus function. Cogn. Affect. Behav. Neurosci. 2010, 10, 252–269. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.R.; Robertson, I.H.; Balsters, J.H.; O’connell, R.G. Pupillometry and P3 index the locus coeruleus–noradrenergic arousal function in humans. Psychophysiology 2011, 48, 1532–1543. [Google Scholar] [CrossRef] [PubMed]
- Siegle, G.J.; Ichikawa, N.; Steinhauer, S. Blink before and after you think: Blinks occur prior to and following cognitive load indexed by pupillary responses. Psychophysiology 2008, 45, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Blaser, E.; Eglington, L.; Carter, A.S.; Kaldy, Z. Pupillometry reveals a mechanism for the Autism Spectrum Disorder (ASD) advantage in visual tasks. Sci. Rep. 2014, 4, 4301. [Google Scholar] [CrossRef] [PubMed]
- Sahley, T.L.; Panksepp, J. Brain opioids and autism: An updated analysis of possible linkages. J. Autism Dev. Disord. 1987, 17, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J. Brain opiods: A neurochemical substrate for narcotic and social dependence. In Theory in Psychopharmacology; Cooper, S.J., Ed.; London Academic Press: London, UK, 1981; Volume 1. [Google Scholar]
- Panksepp, J.; Conner, R.; Forster, P.K.; Bishop, P.; Scott, J.P. Opioid effects on social behavior of kennel dogs. Appl. Anim. Ethol. 1983, 10, 63–74. [Google Scholar] [CrossRef]
- Panksepp, J.; Lensing, P.; Leboyer, M.; Bouvard, M.P. Naltrexone and other potential new pharmacological treatments of autism. Brain Dysfunct. 1991, 4, 281–300. [Google Scholar]
- Bouvard, M.P.; Leboyer, M.; Launay, J.M.; Recasens, C.; Plumet, M.H.; Waller-Perotte, D.; Panksepp, J. Low-dose naltrexone effects on plasma chemistries and clinical symptoms in autism: A double-blind, placebo-controlled study. Psychiatry Res. 1995, 58, 191–201. [Google Scholar] [CrossRef]
- Beversdorf, D.; Ferguson, B.; Reznicek, E.; Lewis, M.; Christ, S.; Stichter, J. Effects of propranolol on social functioning in autism spectrum disorder (S18. 002). Neurology 2013, 80 (Suppl. 7), S18-002. [Google Scholar]
- Ji, N.Y.; Findling, R.L. An update on pharmacotherapy for autism spectrum disorder in children and adolescents. Curr. Opin. Psychiatr 2015, 28, 91–101. [Google Scholar] [CrossRef]
- Kirsch, P.; Esslinger, C.; Chen, Q.; Mier, D.; Lis, S.; Siddhanti, S.; Meyer-Lindenberg, A. Oxytocin modulates neural circuitry for social cognition and fear in humans. J. Neurosci. 2005, 25, 11489–11493. [Google Scholar] [CrossRef] [PubMed]
- Blanke, M.L.; VanDongen, A.M.J. Activation mechanisms of the NMDA receptor. In Biology of the NMDA Receptor; Van Dongen, A.M., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2009. Available online: http://www.ncbi.nlm.nih.gov/books/NBK5274/ (accessed on 1 September 2018).
- Vasa, R.A.; Mazurek, M.O.; Mahajan, R.; Bennett, A.E.; Bernal, M.P.; Nozzolillo, A.A.; Coury, D.L. Assessment and treatment of anxiety in youth with autism spectrum disorders. Pediatrics 2016, 137 (Suppl. 2), S115–S123. [Google Scholar] [CrossRef] [PubMed]
- Sagar-Ouriaghli, I.; Lievesley, K.; Santosh, P.J. Propranolol for treating emotional, behavioural, autonomic dysregulation in children and adolescents with autism spectrum disorders. J. Psychopharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, A.; White, C.A.; Saklayen, S.; Scaduto, M.J.; Carpenter, A.L.; Abduljalil, A.; Beversdorf, D.Q. Effect of propranolol on functional connectivity in autism spectrum disorder—A pilot study. Brain Imaging Behav. 2010, 4, 189–197. [Google Scholar] [CrossRef] [PubMed]
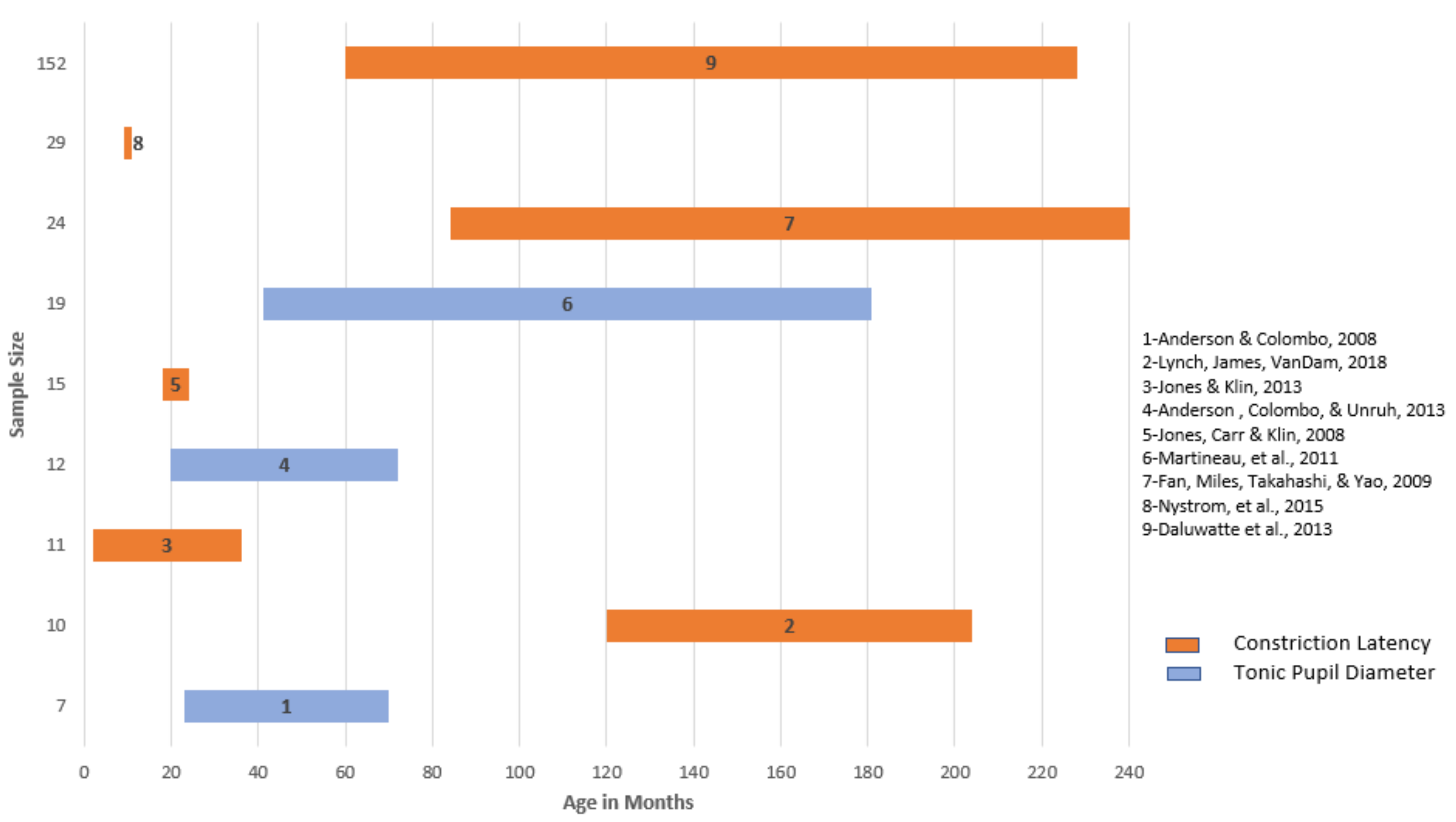
- Fan, X.; Miles, J.H.; Takahashi, N.; Yao, G. Abnormal transient pupillary light reflex in individuals with autism spectrum disorders. J. Autism Dev. Disord. 2013, 39, 1499–1508. [Google Scholar] [CrossRef] [PubMed]
- Nyström, P.; Gredebäck, G.; Bölte, S.; Falck-Ytter, T. Hypersensitive pupillary light reflex in infants at risk for autism. Mol. Autism 2015, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Daluwatte, C.; Miles, J.; Christ, S.; Beversdorf, D.; Takahashi, T.N.; Yao, G. Pupillary light reflex and heart rate variability in children with autism spectrum disorder. J. Autism Dev. Disord. 2013, 43, 1910–1925. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.J.; Colombo, J.; Unruh, K.E. Pupil and salivary indicators of autonomic dysfunction in autism spectrum disorder. Dev. Psychobiol. 2013, 55, 465–482. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.J.; Columbo, J. Larger tonic pupil size in young children with autism spectrum disorder. Dev. Psychobiol. 2008, 51, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Lynch, G.; James, S.; VanDam, M. Pupillary response and phenotype in ASD: Latency to constriction discriminates ASD from typically developing adolescents. Autism Res. 2018, 11, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.A. β-Endorphin inhibition of endogenous norepinephrine release from the A2 noradrenergic nucleus in vitro: Role of Mu opiate receptors and Na+ ion permeability. Brain Res. Bull. 1997, 44, 19–23. [Google Scholar] [CrossRef]
- Martineau, J.; Hernandez, N.; Hiebel, L.; Roche, L.; Metzger, A.; Bonnet-Brilhaut, F. Can pupil size and pupil responses during visual scanning contribute to the diagnosis of autism spectrum disorder in children? J. Psychiatry Res. 2011, 45, 1077–1082. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lynch, G. Using Pupillometry to Assess the Atypical Pupillary Light Reflex and LC-NE System in ASD. Behav. Sci. 2018, 8, 108. https://doi.org/10.3390/bs8110108
Lynch G. Using Pupillometry to Assess the Atypical Pupillary Light Reflex and LC-NE System in ASD. Behavioral Sciences. 2018; 8(11):108. https://doi.org/10.3390/bs8110108
Chicago/Turabian StyleLynch, Georgina. 2018. "Using Pupillometry to Assess the Atypical Pupillary Light Reflex and LC-NE System in ASD" Behavioral Sciences 8, no. 11: 108. https://doi.org/10.3390/bs8110108
APA StyleLynch, G. (2018). Using Pupillometry to Assess the Atypical Pupillary Light Reflex and LC-NE System in ASD. Behavioral Sciences, 8(11), 108. https://doi.org/10.3390/bs8110108