Ambiguous Results When Using the Ambiguous-Cue Paradigm to Assess Learning and Cognitive Bias in Gorillas and a Black Bear


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experiment 1
2.1. Materials and Methods
2.1.1. Subjects
2.1.2. Materials
2.1.3. Training
2.1.4. Testing
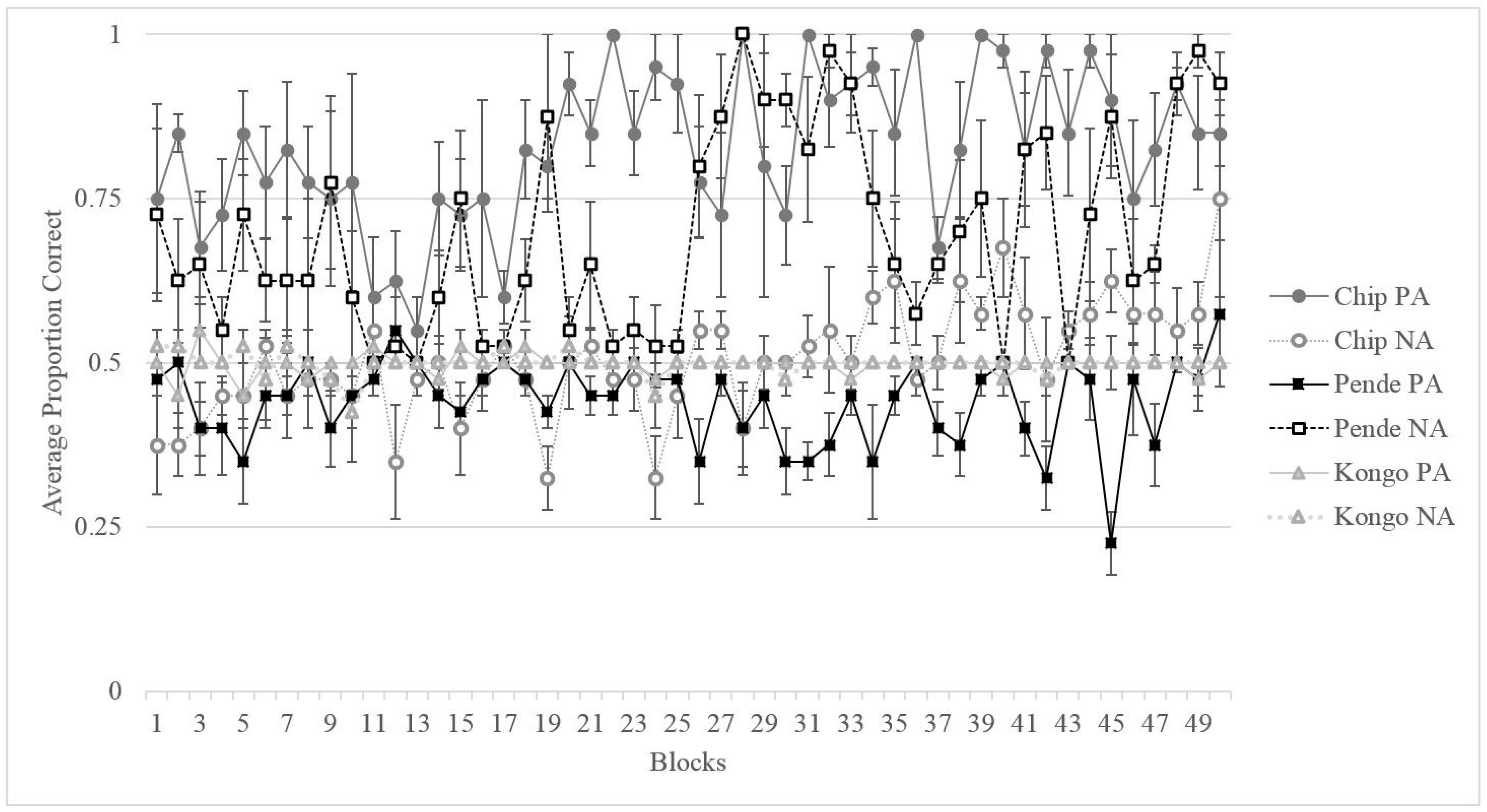
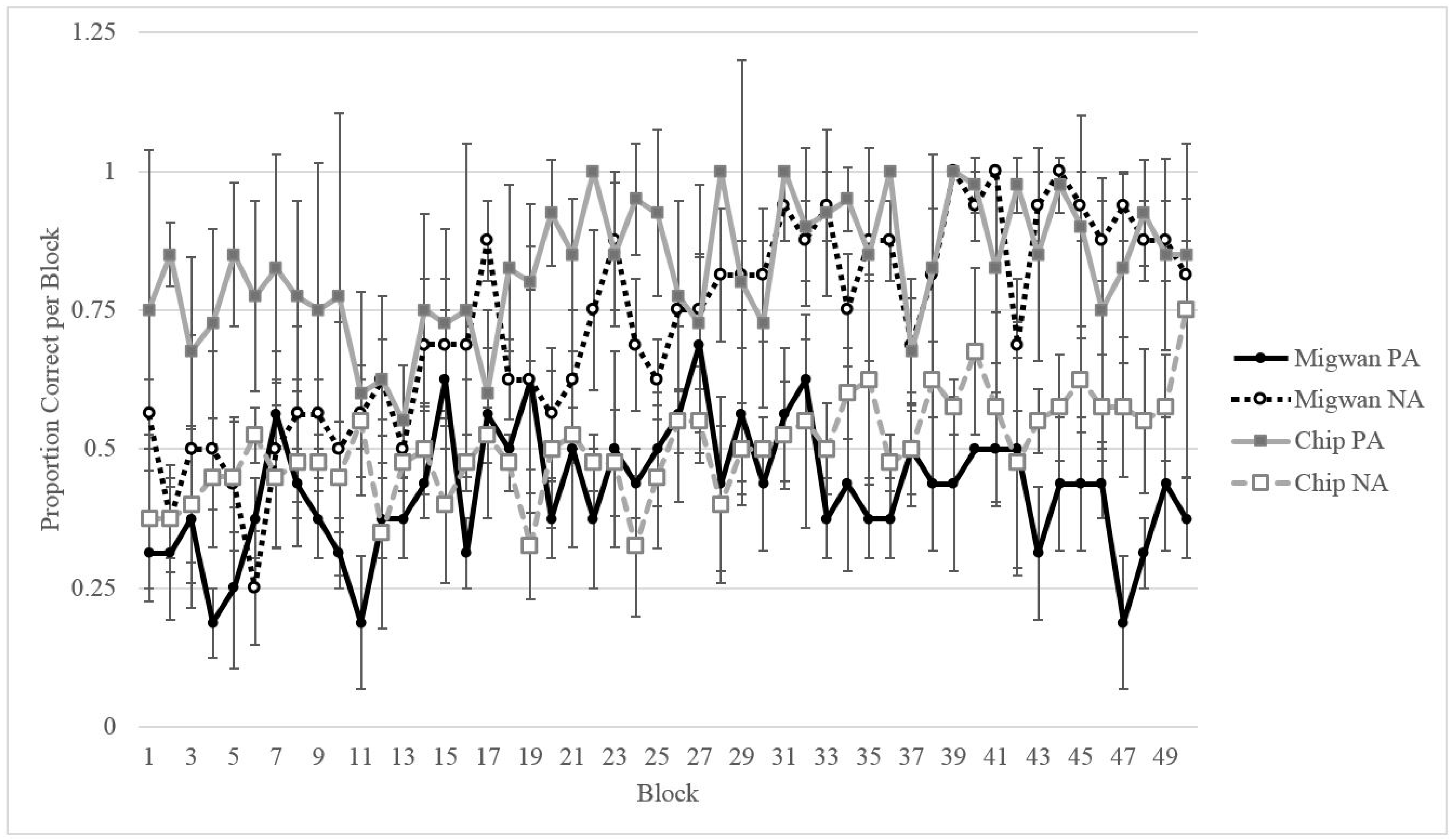
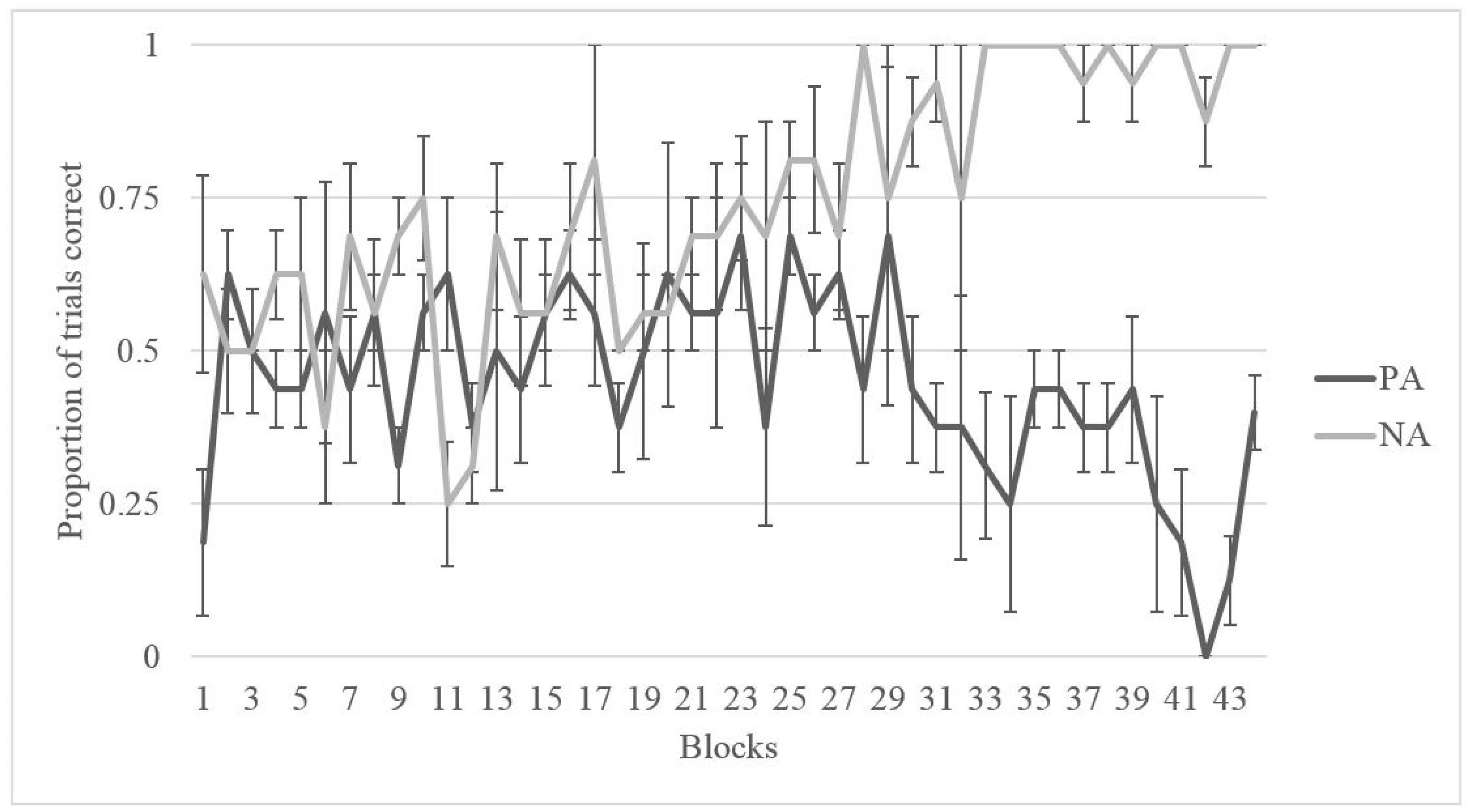
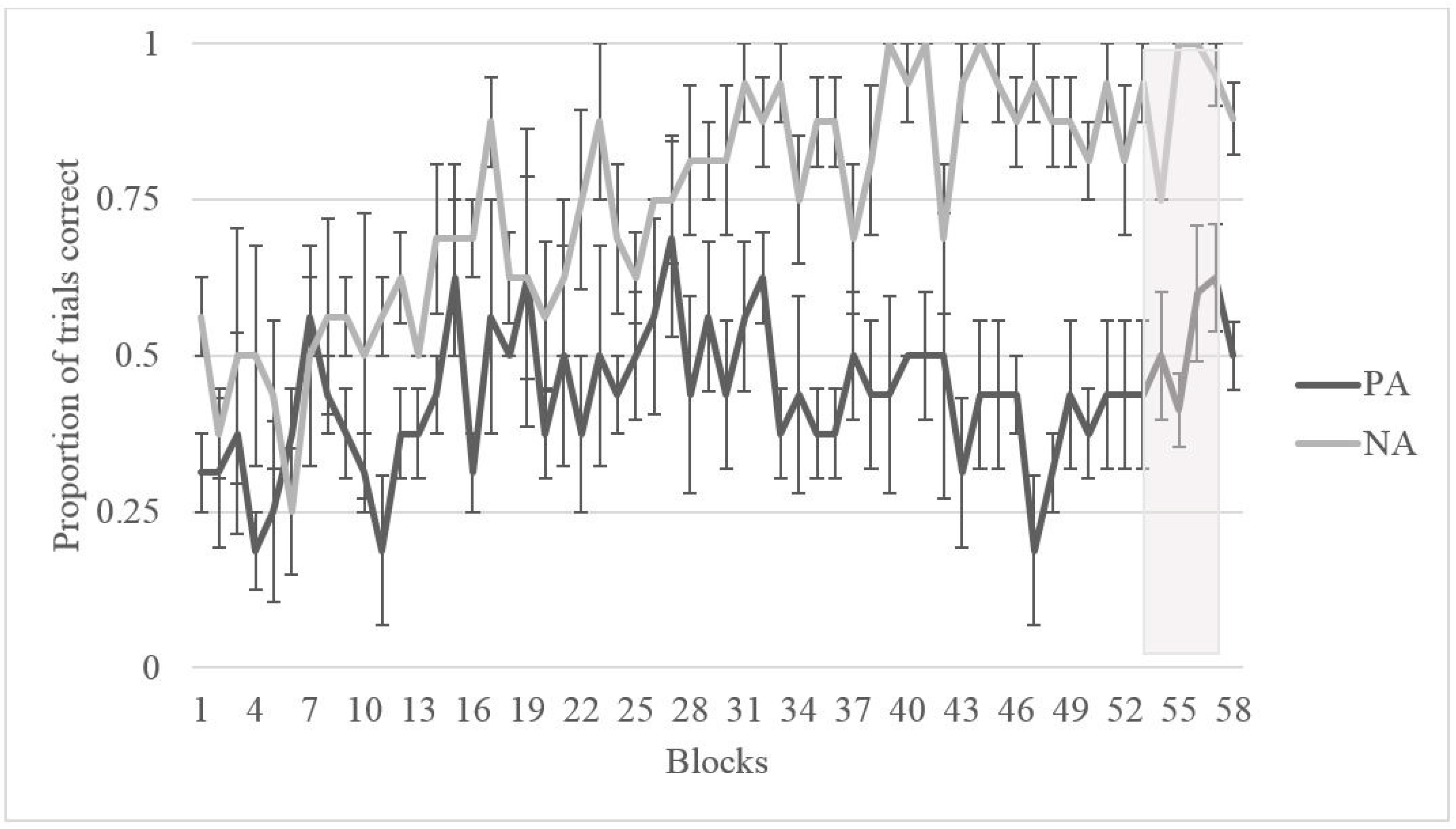
2.2. Results
2.2.1. Training
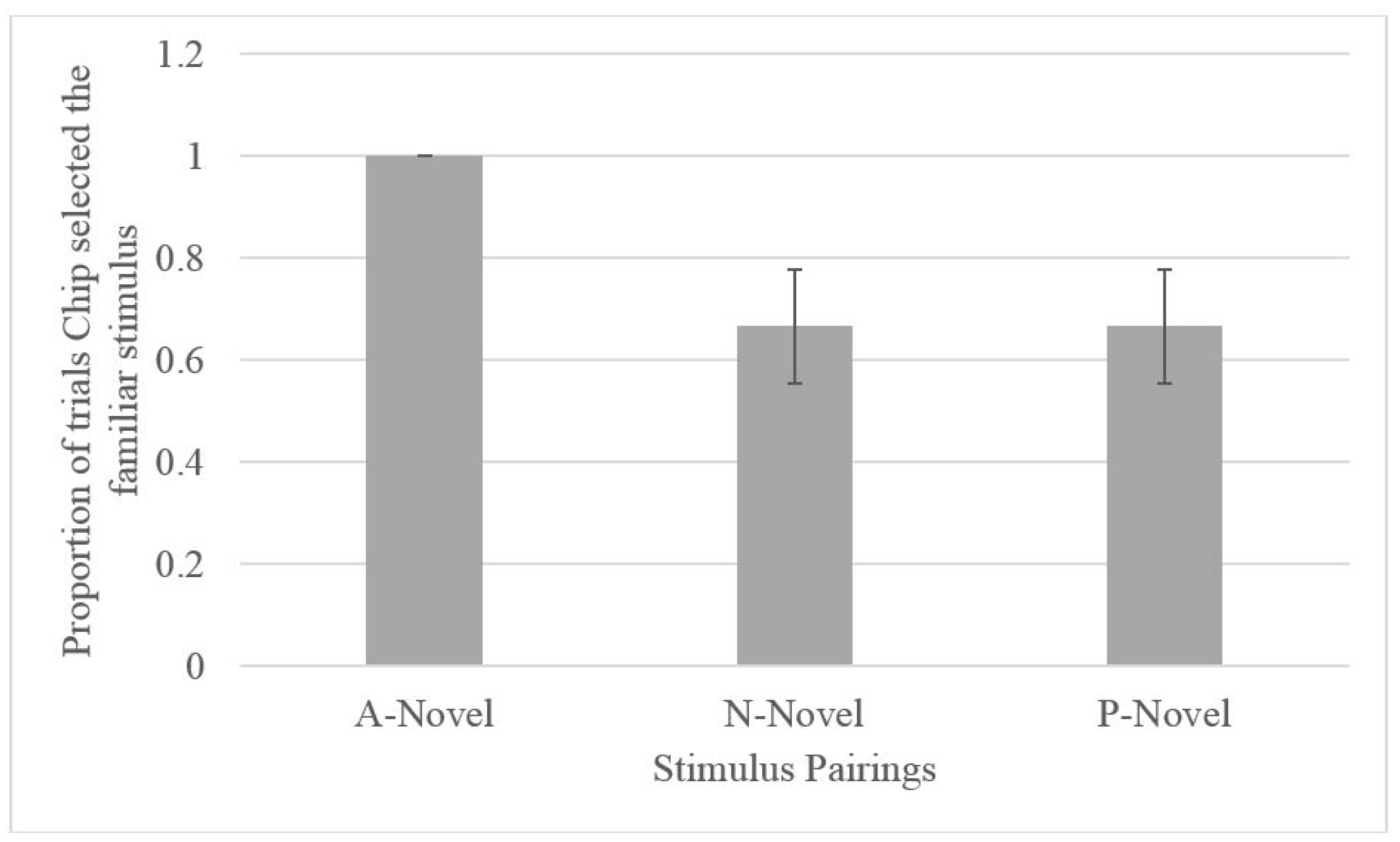
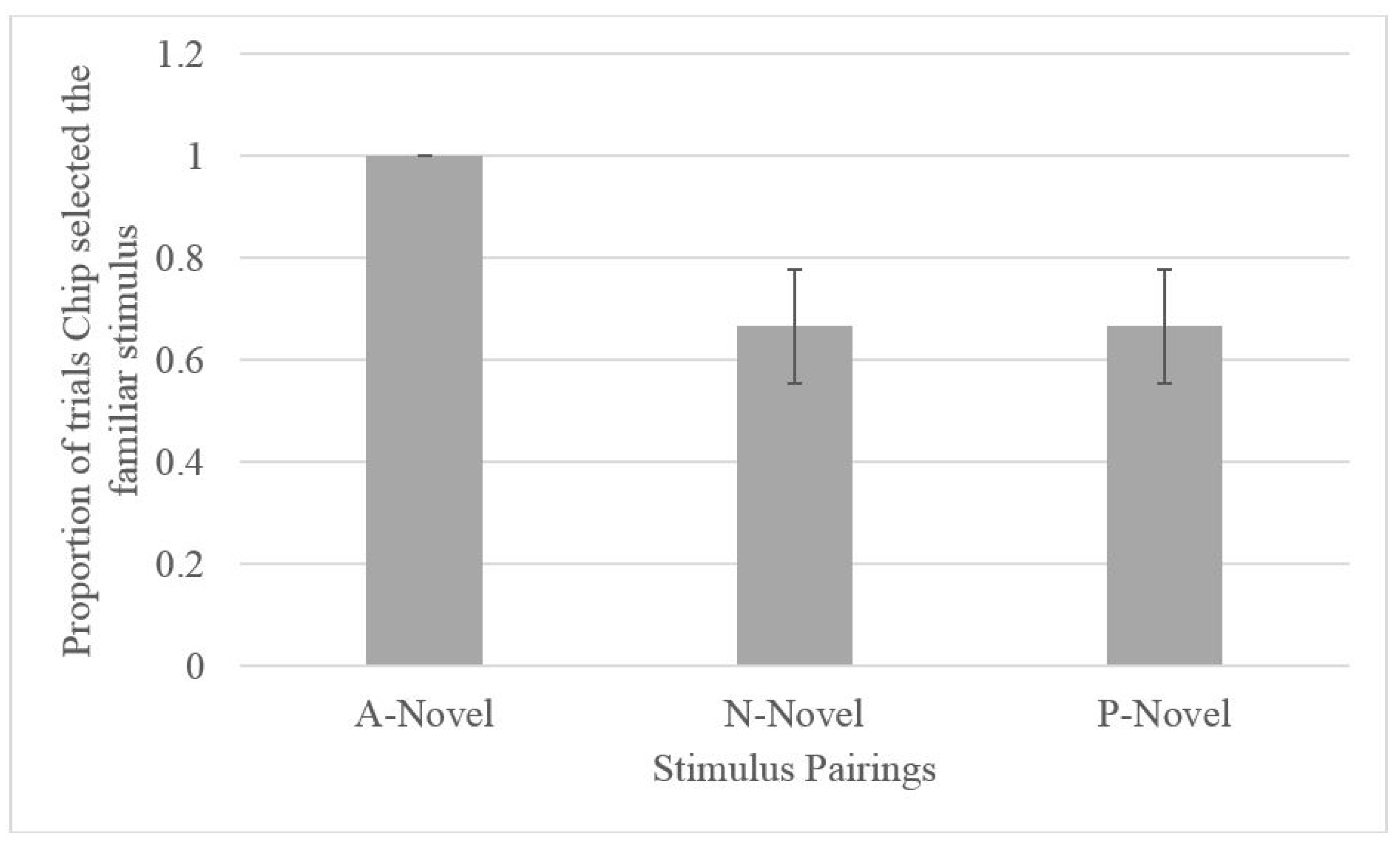
2.2.2. Testing
2.3. Discussion
3. Experiment 2
3.1. Materials and Methods
3.1.1. Subjects
3.1.2. Testing Environment
3.1.3. Materials
3.1.4. Training
3.1.5. Testing
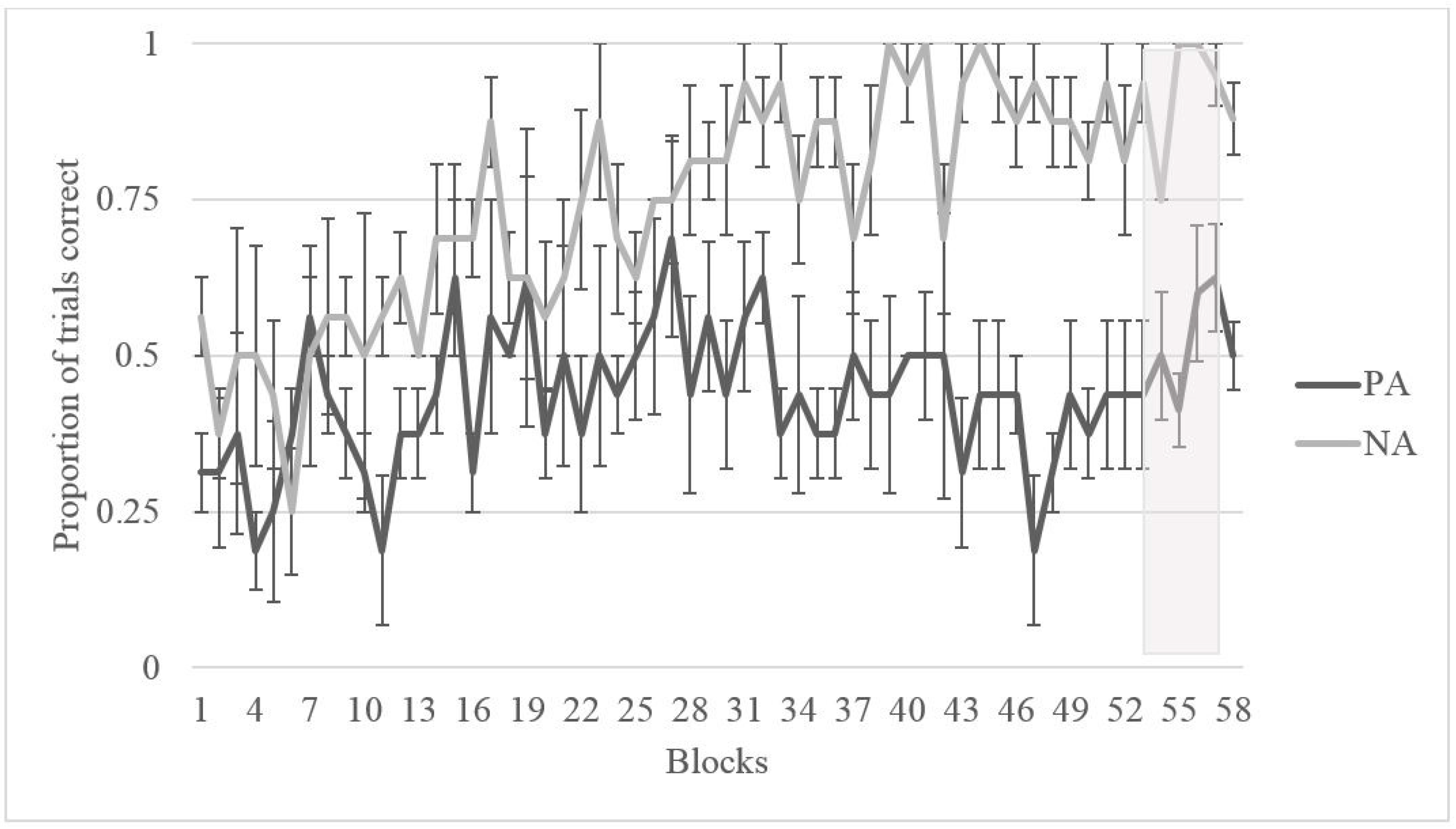
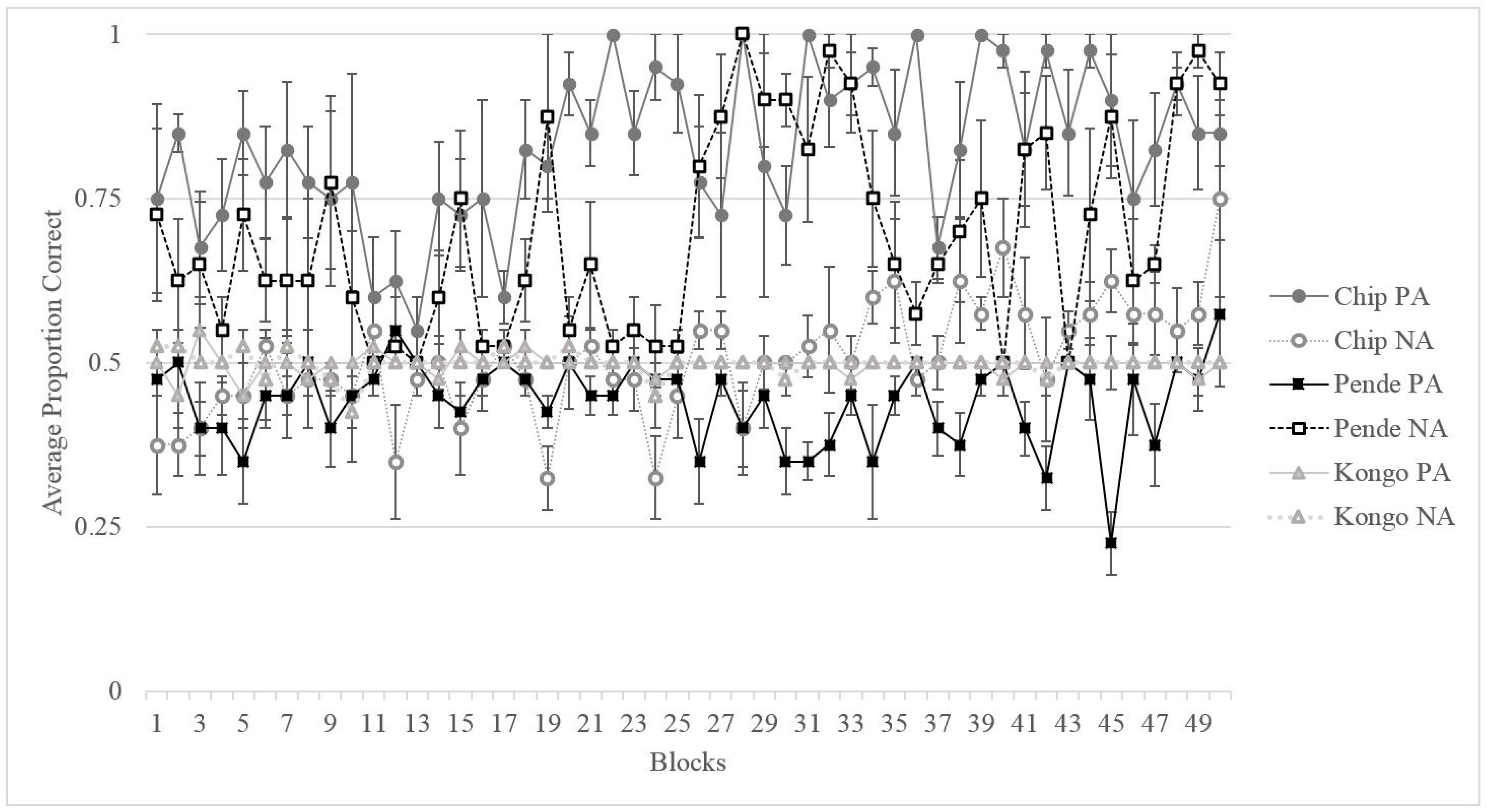
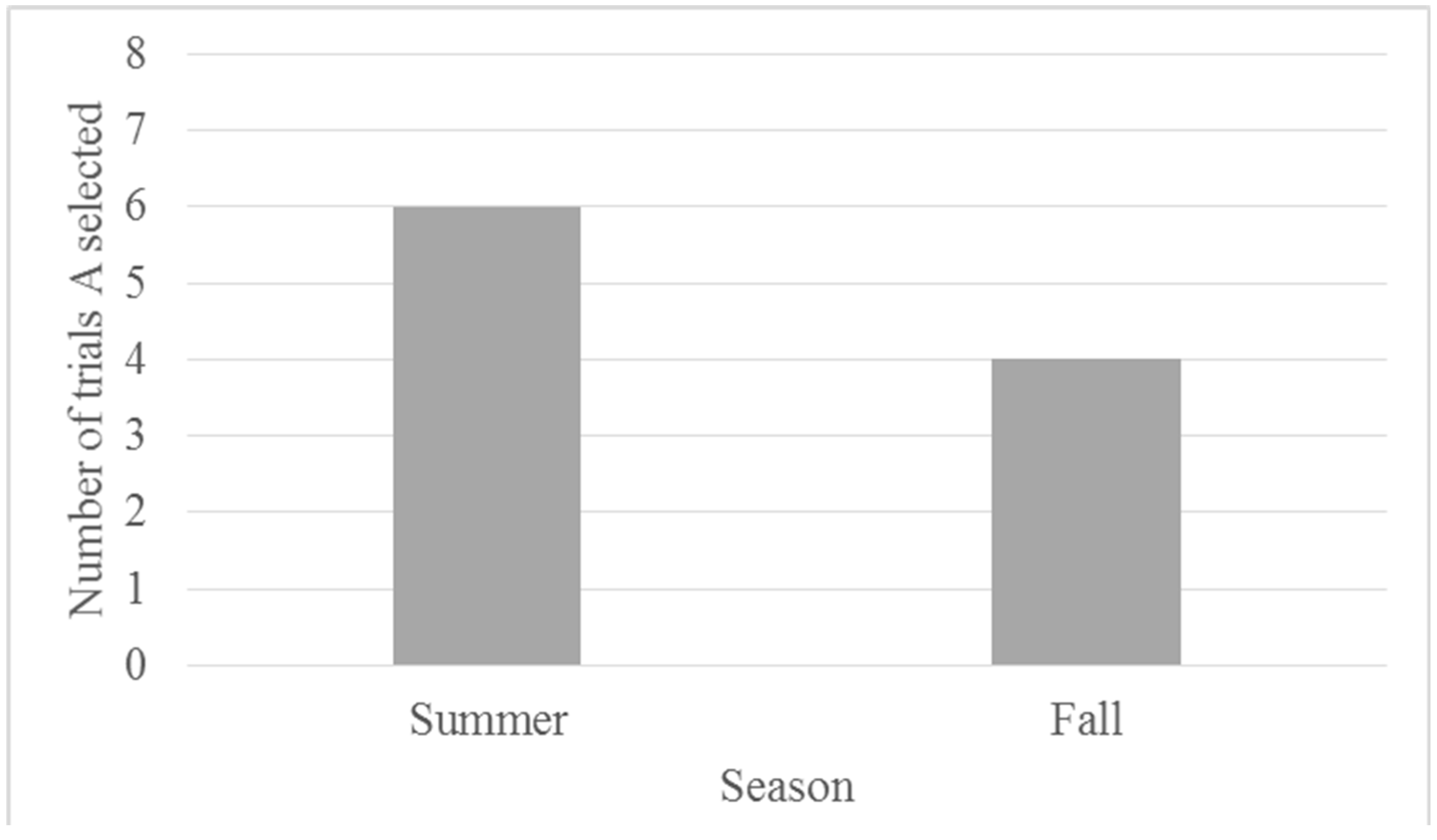
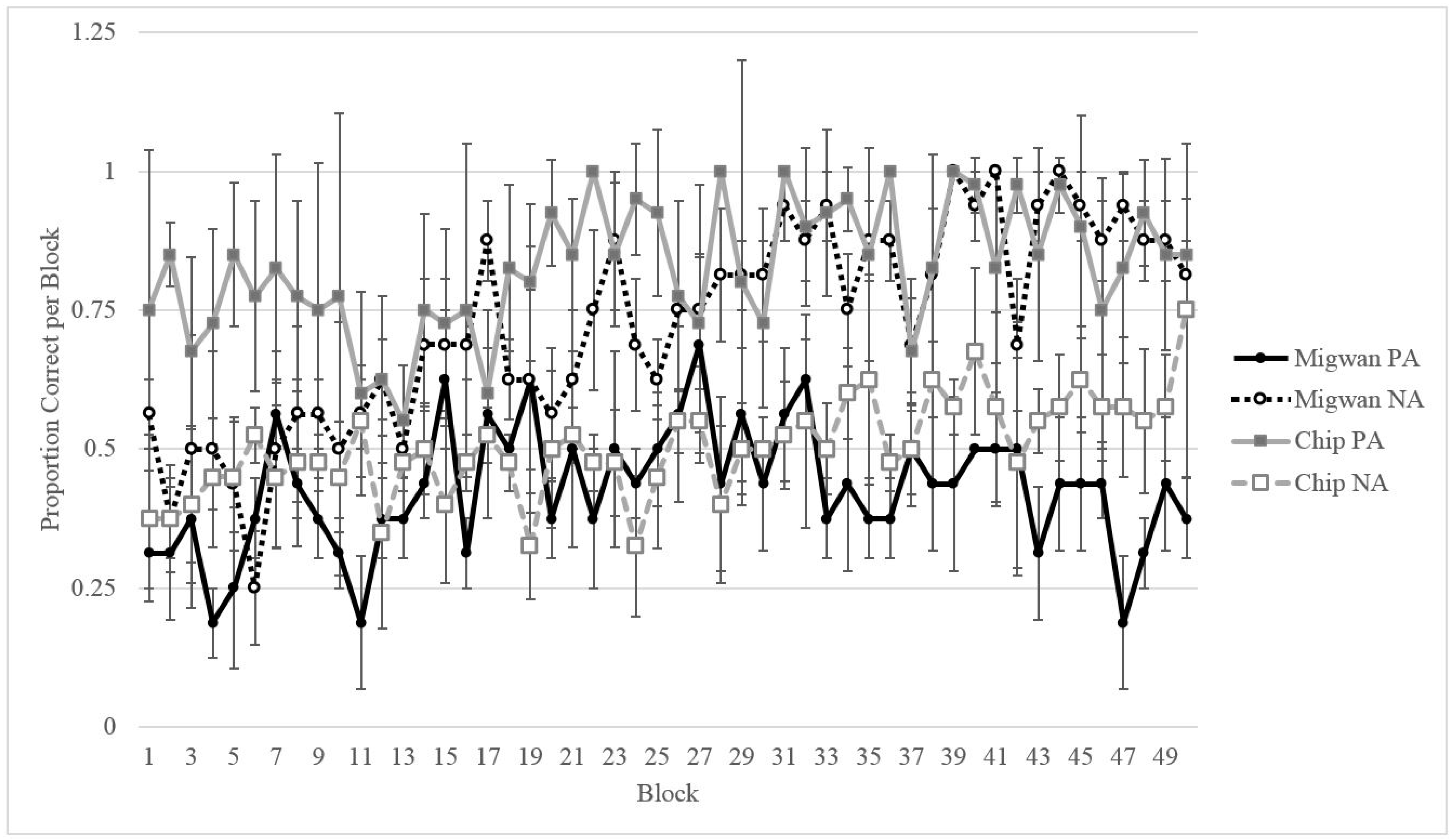
3.2. Results
3.2.1. Training
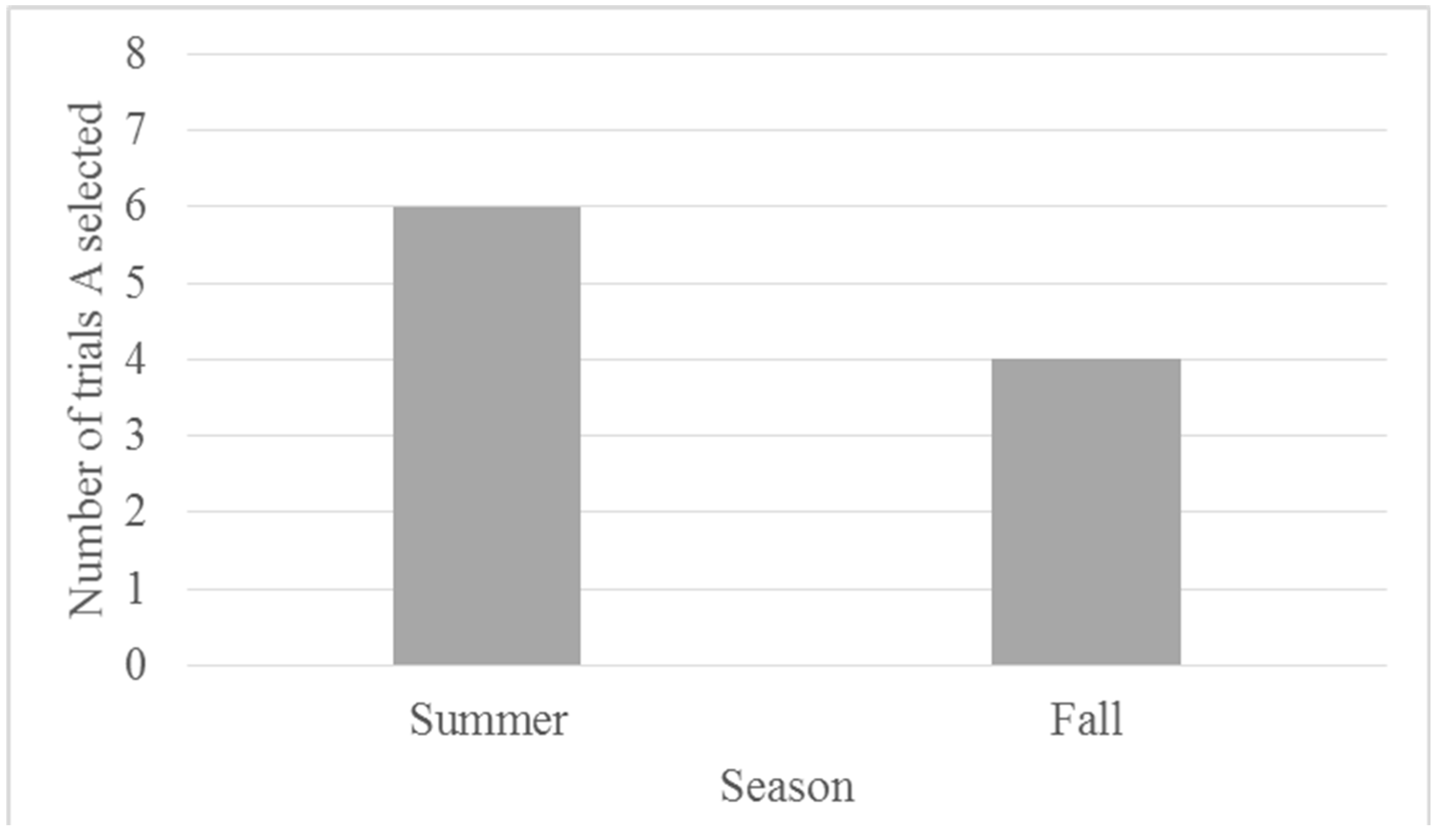
3.2.2. Testing
3.2.3. Comparing Migwan’s Performance to That of the Gorillas
3.3. Discussion
4. General Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Leary, R.W. The learning of ambiguous cue problems by monkeys. Am. J. Psychol. 1958, 71, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R. Approach versus avoidance in an ambiguous-cue discrimination problem in chimpanzees. J. Comp. Physiol. Psychol. 1954, 47, 133–135. [Google Scholar] [CrossRef] [PubMed]
- McGuire, M.C.; Vonk, J.; Fuller, G.; Allard, S. Using an ambiguous cue paradigm to assess cognitive bias in gorillas (Gorilla gorilla gorilla) during a forage manipulation. Anim. Behav. Cogn. 2017, 4, 70–83. [Google Scholar] [CrossRef]
- Richards, R.W. Performance of the pigeon on the ambiguous-cue problem. Bull. Psychon. Soc. 1973, 1, 445–447. [Google Scholar] [CrossRef]
- Fletcher, H.J.; Grogg, T.M.; Garske, J.P. Ambiguous-cue problem performance of children, retardates, and monkeys. J. Comp. Physiol. Psychol. 1968, 66, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Boyer, W.N.; Polidora, V.J. An analysis of the solution of PAN ambiguous-cue problems by Rhesus monkeys. Learn. Motiv. 1972, 3, 325–333. [Google Scholar] [CrossRef]
- Boyer, W.N.; Polidora, V.J.; Fletcher, H.J.; Woodruff, B. Monkeys’ performance on ambiguous-cue problems’. Percept. Motor Skills 1966, 22, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, H.J.; Garske, J.P. Response competition in monkeys’ solution of PAN ambiguous-cue problems. Learn. Motiv. 1972, 3, 334–340. [Google Scholar] [CrossRef]
- Hall, G. An investigation of ambiguous-cue learning in pigeons. Anim. Learn. Behav. 1980, 8, 282–286. [Google Scholar] [CrossRef]
- Richards, R.W.; Marcattilio, A.J. Intermittency of reinforcement during NA trials and performance on the ambiguous-cue problem. Can. J. Exp. Psychol. 1975, 29, 210–223. [Google Scholar] [CrossRef]
- Vasconcelos, M.; Monteiro, T. European starlings unriddle the ambiguous-cue problem. Front. Psychol. 2014, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, H.J.; Bordow, A.M. Monkey’s solution of an ambigious-cue problem. Percept. Motor Skills 1965, 21, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Hallion, L.S.; Ruscio, A.M. A meta-analysis of the effect of cognitive bias modification on anxiety and depression. Psychol. Bull. 2011, 137, 940–958. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.S.; Harding, E.J.; Mendl, M. Measuring emotional processes in animals: The utility of a cognitive approach. Neurosci. Biobehav. Rev. 2005, 3, 469–491. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.; Burman, O.H.P.; Parker, R.M.A.; Paul, E.S. Cognitive bias as an indicator of animal emotion and welfare: Emerging evidence and underlying mechanisms. Appl. Anim. Behav. Sci. 2009, 118, 161–181. [Google Scholar] [CrossRef]
- Byrdges, N.M.; Leach, M.; Nicol, K.; Wright, R.; Bateson, M. Environmental enrichment induces optimistic cognitive bias in rats. Anim. Behav. 2011, 81, 169–175. [Google Scholar] [CrossRef]
- Matheson, S.M.; Asher, L.; Bateson, M. Larger, enriched cages are associated with “optimistic” response biases in captive European starlings (Sturnus vulgaris). Appl. Anim. Behav. Sci. 2008, 109, 374–383. [Google Scholar] [CrossRef]
- Keen, H.A.; Nelson, O.L.; Robbins, C.T.; Evans, M.; Shepherdson, D.J.; Newberry, R.C. Validation of a novel cognitive bias task based on difference in quantity of reinforcement for assessing environmental enrichment. Anim. Cogn. 2014, 17, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Fuller, G.A.; Vonk, J.; McGuire, M.; Allard, S. The influence of browse availabilityon behavior and cognitive bias in captive western lowland gorillas. (Gorilla gorilla gorilla). Am. J. Primatol. 2015, 77, 139. [Google Scholar]
- Harding, E.J.; Paul, E.S.; Mendl, M. Animal Behabiour: Cognitive bias and affective state. Nature 2004, 427, 312. [Google Scholar] [CrossRef] [PubMed]
- Vonk, J.; Beran, M.J. Bears “count” too: Quantity estimation and comparison in black bears (Ursus Americanus). Anim. Behav. 2012, 84, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Vonk, J.; Galvan, M. What do natural categorization studies tell us about apes and bears. Anim. Behav. Cogn. 2014, 1, 309–330. [Google Scholar] [CrossRef]
- Vonk, J.; Johnson-Ulrich, Z. Social and nonsocial category discriminations in a chimpanzee (Pan troglodytes) and American black bears (Ursus americanus). Learn. Behav. 2014, 42, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Vonk, J.; Jett, S.E.; Mosteller, K.W. Concept formation in American black bears, Ursus americanus. Anim. Behav. 2012, 84, 953–964. [Google Scholar] [CrossRef]
- Vonk, J.; Jett, S.E.; Mosteller, K.W.; Galvan, M. Natural category discrimination in chimpanzees (Pan troglodytes) at three levels of abstraction. Learn. Behav. 2013, 41, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Vonk, J.; Torgerson-White, L.; McGuire, M.; Thueme, M.; Thomas, J.; Beran, M.J. Quantity estimation and comparison in Western Lowland Gorillas (Gorilla gorilla gorilla). Anim. Cogn. 2014, 17, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Benson-Amram, S.; Dantzer, B.; Stricker, G.; Swanson, E.M.; Holekamp, K.E. Brain size predicts problem-solving ability in mammalian carnivores. Proc. Natl. Acad. Sci. USA 2016, 113, 2532–2537. [Google Scholar] [CrossRef] [PubMed]
- Rosati, A.G. Foraging cognition: Revising the ecological intelligence hypothesis. Trends Cogn. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Gittleman, J.R. Carnivore brain size, behavioral ecology, and phylogeny. J. Mammal. 1986, 67, 23–36. [Google Scholar] [CrossRef]
- McGuire, M.C. Influences on Cognitive Bias in Nonhuman Animals. Ph.D. Dissertation, Oakland University, Rochester, MI, USA, 2017. [Google Scholar]
- Johnson-Ulrich, Z.; Vonk, J.; Humbyrd, M.; Crowley, M.; Wojtkowski, E.; Yates, F.; Allard, S. Picture object recognition in an American black bear (Ursus americanus). Anim. Cogn. 2016, 19, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Tøien, Ø.; Blake, J.; Edgar, D.M.; Grahn, D.A.; Heller, H.C.; Barnes, B.M. Hibernation in black bears: Independence of metabolic suppression from body temperature. Science 2011, 331, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Tomasi, T.E.; Hellgren, E.C.; Tucker, T.J. Thyroid hormone concentrations in black bears (Ursus americanus): Hibernation and pregnancy effects. Gen. Comp. Endocr. 1998, 109, 192–199. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGuire, M.C.; Vonk, J.; Johnson-Ulrich, Z. Ambiguous Results When Using the Ambiguous-Cue Paradigm to Assess Learning and Cognitive Bias in Gorillas and a Black Bear. Behav. Sci. 2017, 7, 51. https://doi.org/10.3390/bs7030051
McGuire MC, Vonk J, Johnson-Ulrich Z. Ambiguous Results When Using the Ambiguous-Cue Paradigm to Assess Learning and Cognitive Bias in Gorillas and a Black Bear. Behavioral Sciences. 2017; 7(3):51. https://doi.org/10.3390/bs7030051
Chicago/Turabian StyleMcGuire, Molly C., Jennifer Vonk, and Zoe Johnson-Ulrich. 2017. "Ambiguous Results When Using the Ambiguous-Cue Paradigm to Assess Learning and Cognitive Bias in Gorillas and a Black Bear" Behavioral Sciences 7, no. 3: 51. https://doi.org/10.3390/bs7030051
APA StyleMcGuire, M. C., Vonk, J., & Johnson-Ulrich, Z. (2017). Ambiguous Results When Using the Ambiguous-Cue Paradigm to Assess Learning and Cognitive Bias in Gorillas and a Black Bear. Behavioral Sciences, 7(3), 51. https://doi.org/10.3390/bs7030051