

Executive Function and Transfer Effect Training in Children: A Behavioral and Event-Related Potential Pilot Study

Abstract
1. Introduction
2. Experiment 1
2.1. Methods
2.1.1. Participants
2.1.2. Pre- and Post-Training Test Tasks
Number 2-Back Task
Arrow Flanker Task
Number Switching Task
Raven’s Advanced Progressive Matrix Test
2.1.3. Training Task
Letter 2-Back Task
Fish Flanker Task
2.1.4. Procedures
2.2. Statistical Analysis
2.3. Results
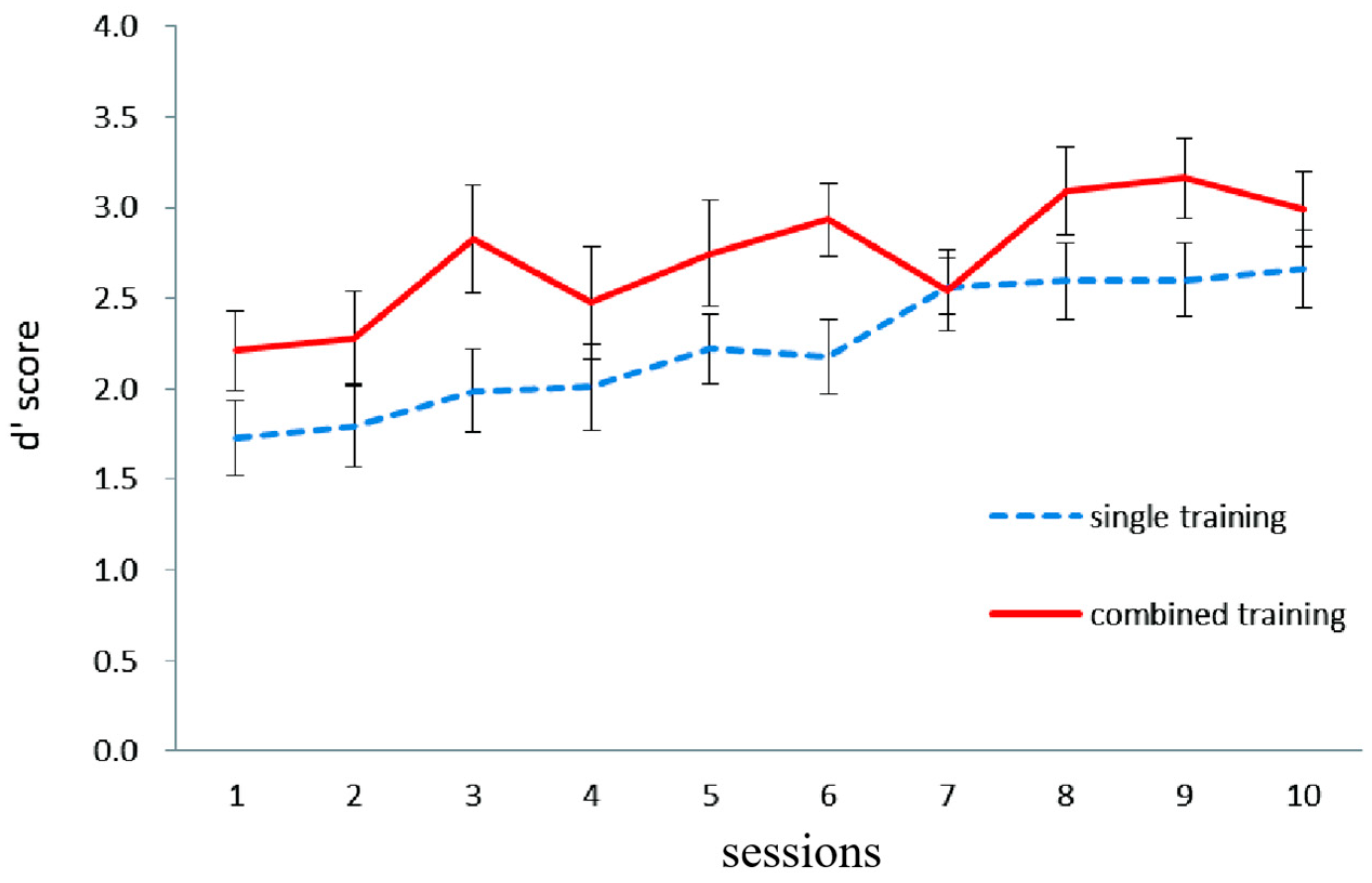
2.3.1. Training Results
Letter 2-Back Task
Fish Flanker Task
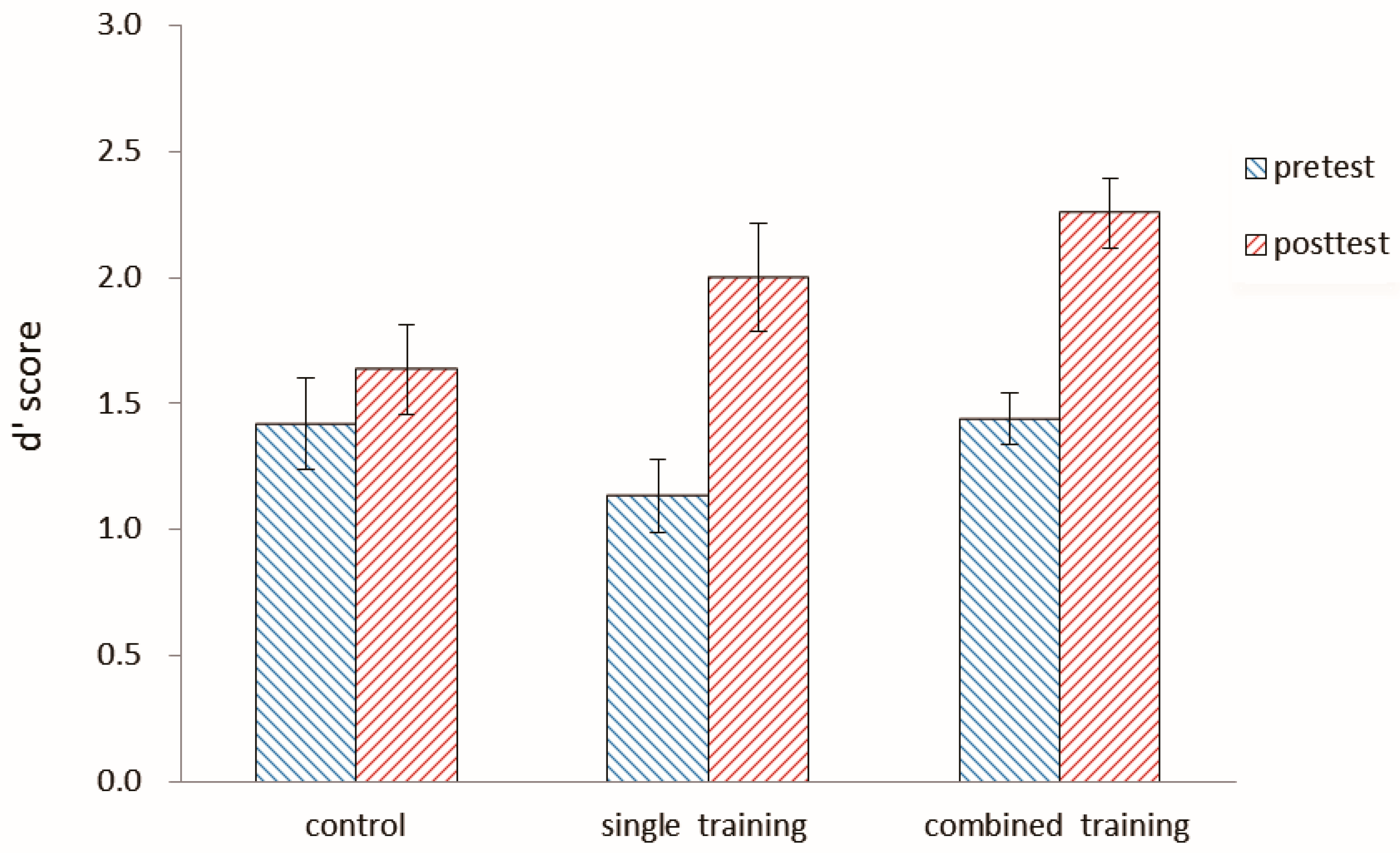
2.3.2. Near Transfer Results
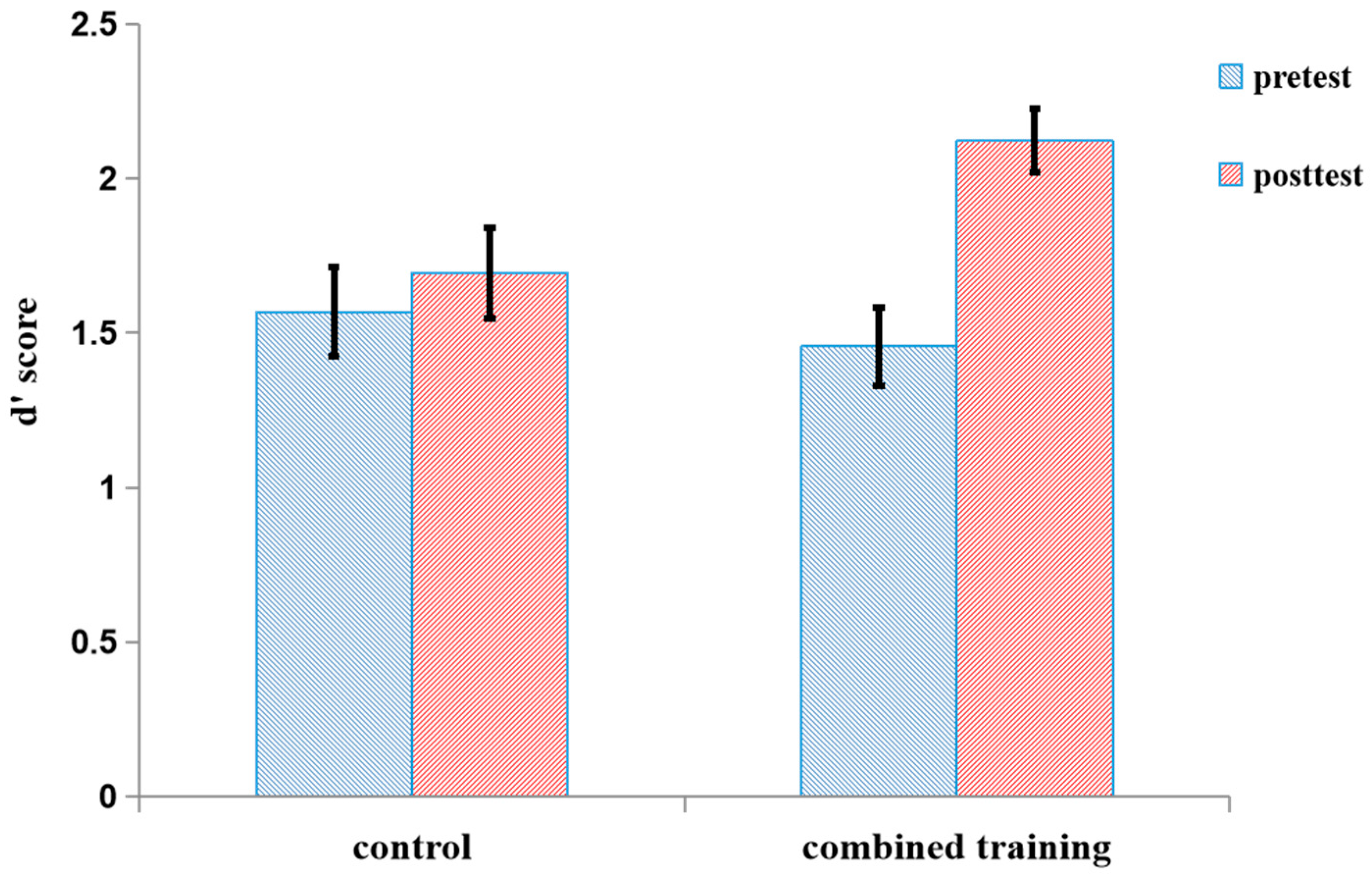
Number 2-Back
Arrow Flanker Task
2.3.3. Far Transfer Results on Both the Switching Task and Fluid Intelligence
Switching Task
Fluid Intelligence
2.4. Discussion
3. Experiment 2
3.1. Methods
3.1.1. Participants
3.1.2. Training Tasks and Procedures
3.1.3. EEG Recording and Analysis
3.1.4. Behavioral Data Analysis
3.2. Behavior Results
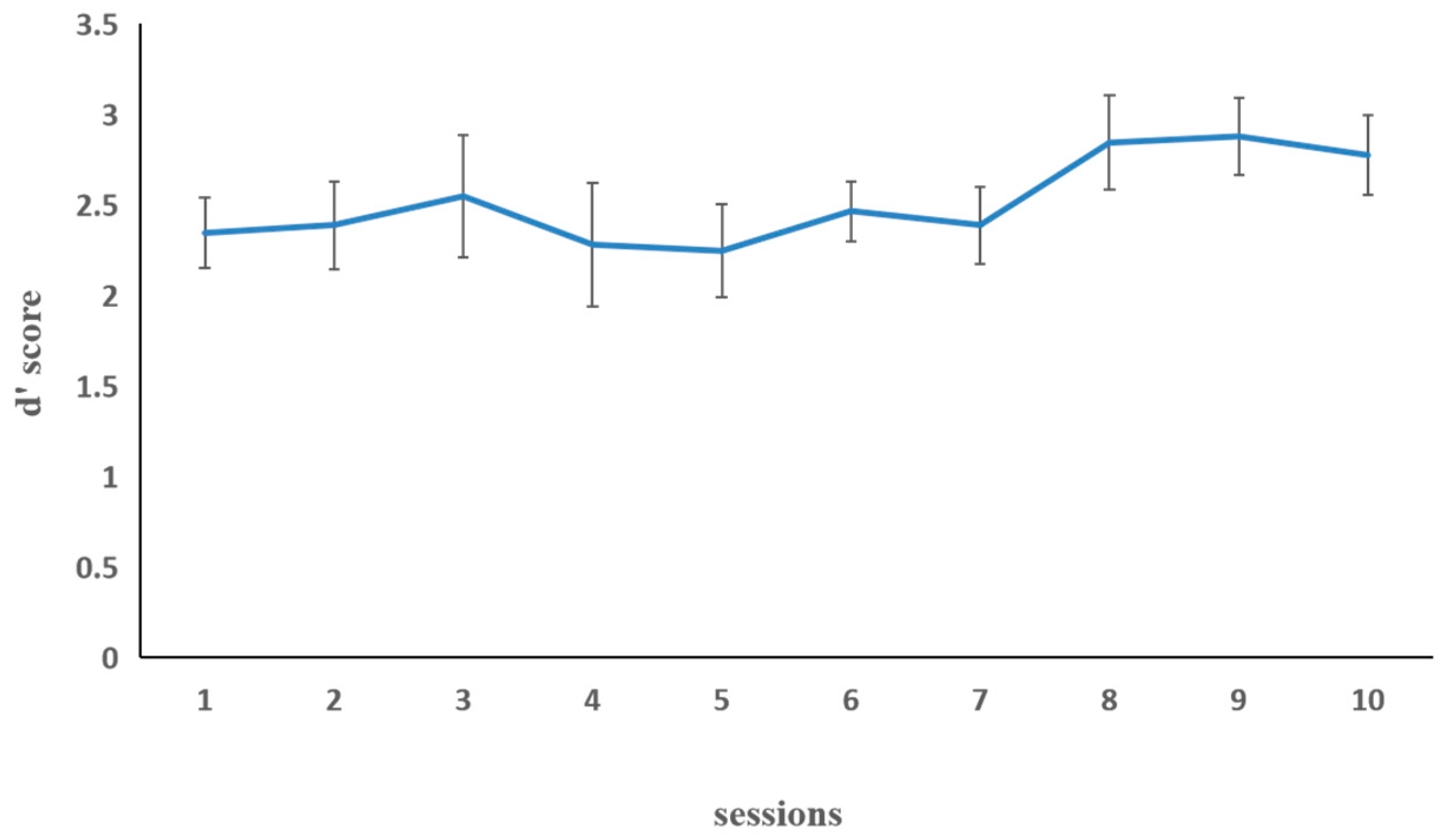
3.2.1. Training Results
Letter 2-Back Task
Fish Flanker Task
3.2.2. Near Transfer Results
Number 2-Back Task
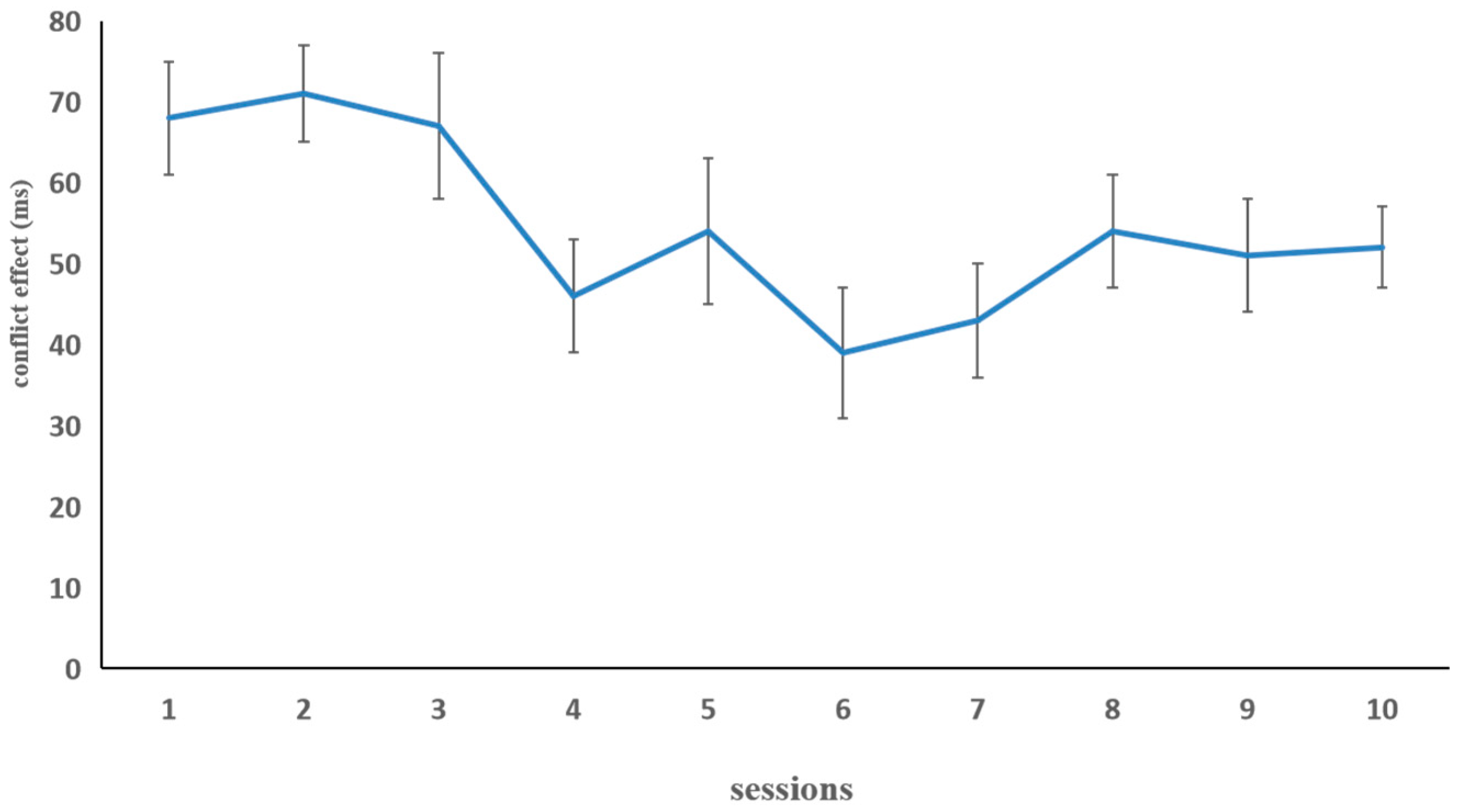
Arrow Flanker Task
3.3. ERP Results
- N2 Amplitude
- P300 Amplitude
3.4. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bailey, D., Duncan, G. J., Odgers, C. L., & Yu, W. (2017). Persistence and fadeout in the impacts of child and adolescent interventions. Journal of Research on Educational Effectiveness, 10(1), 7–39. [Google Scholar] [CrossRef] [PubMed]
- Baum, G. L., Ciric, R., Roalf, D. R., Betzel, R. F., Moore, T. M., Shinohara, R. T., Kahn, A. E., Vandekar, S. N., Rupert, P. E., Quarmley, M., Cook, P. A., Elliott, M. A., Ruparel, K., Gur, R. E., Gur, R. C., Bassett, D. S., & Satterthwaite, T. D. (2017). Modular segregation of structural brain networks supports the development of executive function in youth. Current Biology, 27(11), 1561–1572. [Google Scholar] [CrossRef]
- Best, J. R., & Miller, P. H. (2010). A developmental perspective on executive function. Child Development, 81(6), 1641–1660. [Google Scholar] [CrossRef]
- Bielak, A. A., Hatt, C. R., & Diehl, M. (2017). Cognitive performance in adults’ daily lives: Is there a lab-life gap? Research in Human Development, 14(3), 219–233. [Google Scholar] [CrossRef]
- Birtwistle, E., Chernikova, O., Wünsch, M., & Niklas, F. (2025). Training of executive functions in children: A meta-analysis of cognitive training interventions. SAGE Open, 15(1), 21582440241. [Google Scholar] [CrossRef]
- Cassotti, M., Agogué, M., Camarda, A., Houdé, O., & Borst, G. (2016). Inhibitory control as a core process of creative problem solving and idea generation from childhood to adulthood. New Directions for Child and Adolescent Development, 151, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Checa, P., Castellanos, M. C., Abundis-Gutiérrez, A., & Rosario Rueda, M. (2014). Development of neural mechanisms of conflict and error processing during childhood: Implications for self-regulation. Frontiers in Psychology, 5, 326. [Google Scholar] [CrossRef] [PubMed]
- Chen, T., & Li, D. (2007). The roles of working memory updating and processing speed in mediating age-related differences in fluid intelligence. Aging Neuropsychology and Cognition, 14(6), 631–646. [Google Scholar] [CrossRef]
- Chen, X., Ye, M., Chang, L., Chen, W., & Zhou, R. (2018). Effect of working memory updating training on retrieving symptoms of children with learning disabilities. Journal of Learning Disabilities, 51(5), 507–519. [Google Scholar] [CrossRef]
- Chueh, T. Y., Hung, C. L., Chang, Y. K., Huang, C. J., & Hung, T. M. (2023). Effects of cognitive demand during acute exercise on inhibitory control and its electrophysiological indices: A randomized crossover study. Physiology & Behavior, 265, 114148. [Google Scholar]
- Cohen, J. (2013). Statistical power analysis for the behavioral sciences. Academic Press. [Google Scholar]
- Cohen, J. D. (1992). A power primer. Psychological Bulletin, 112(1), 155–159. [Google Scholar] [CrossRef]
- Covey, T. J., Shucard, J. L., & Shucard, D. W. (2019). Working memory training and perceptual discrimination training impact overlapping and distinct neurocognitive processes: Evidence from event-related potentials and transfer of training gains. Cognition, 182, 50–72. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, E., Neely, A. S., Larsson, A., Bäkman, L., & Nyberg, L. (2008). Transfer of learning after updating training mediated by the striatum. Science, 320(5882), 1510–1512. [Google Scholar] [CrossRef] [PubMed]
- Danek, R. H., & Mordkoff, J. T. (2011). Unequal motor durations under simple-, go/no-go, and choice-RT tasks: Extension of miller and low (2001). Journal of Experimental Psychology: Human Perception and Performance, 37(4), 1323. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A. (2013). Executive functions. Annual Review of Psychology, 64(1), 135–168. [Google Scholar] [CrossRef]
- Dimmitt, C., & McCormick, C. B. (2012). Metacognition in education. In K. R. Harris, S. Graham, T. Urdan, C. B. McCormick, G. M. Sinatra, & J. Sweller (Eds.), APA educational psychology handbook, Vol 1: Theories, constructs, and critical issues (pp. 157–187). APA. [Google Scholar]
- Donchin, E., & Coles, M. G. (1988). Is the P300 component a manifestation of context updating? Behavioral and Brain Sciences, 11(3), 357–374. [Google Scholar] [CrossRef]
- Dong, P., Li, W., Hu, Q., Wu, T., Jiang, Y., Jin, H., Xu, C., Buschkuehl, M., Jaeggi, S. M., & Zhang, Q. (2024). The relation between effortful control and executive function training in preschoolers. Journal of Experimental Child Psychology, 238, 105778. [Google Scholar] [CrossRef]
- Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16(1), 143–149. [Google Scholar]
- Faul, F., Erdfelder, E., Buchner, A., & Lang, A. G. (2009). Statistical power analyses using G*power 3.1: Tests for correlation and regression analyses. Behavior Research Methods, 41(4), 1149–1160. [Google Scholar] [CrossRef]
- Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007). G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39, 175–191. [Google Scholar] [CrossRef]
- Fields, E. C. (2023). The P300, the LPP, context updating, and memory: What is the functional significance of the emotion-related late positive potential? International Journal of Psychophysiology, 192, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Fleming, K. A., Heintzelman, S. J., & Bartholow, B. D. (2016). Specifying associations between conscientiousness and executive functioning: Mental set shifting, not prepotent response inhibition or working memory updating. Journal of Personality, 84(3), 348–360. [Google Scholar] [CrossRef]
- Gevins, A., & Smith, M. E. (2000). Neurophysiological measures of working memory and individual differences in cognitive ability and cognitive style. Cerebral Cortex, 10(9), 829–839. [Google Scholar] [CrossRef]
- Giedd, J. N., Shaw, P., Wallace, G., Gogtay, N., & Lenroot, R. K. (2015). Anatomic brain imaging studies of normal and abnormal brain development in children and adolescents. In Developmental psychopathology: Volume two: Developmental neuroscience (pp. 127–196). Wiley Online Library. [Google Scholar]
- Gongora, M., Nicoliche, E., Magalhães, J., Vicente, R., Teixeira, S., Bastos, V. H., Bittencourt, J., Cagy, M., Basile, L. F., Budde, H., Velasques, B., & Ribeiro, P. (2021). Event-related potential (P300): The effects of levetiracetam in cognitive performance. Neurological Sciences, 42, 2309–2316. [Google Scholar] [CrossRef]
- Guo, A., Yang, W., Yang, X., Lin, J., Li, Z., Ren, Y., Yang, J., & Wu, J. (2023). Audiovisual n-back training alters the neural processes of working memory and audiovisual integration: Evidence of changes in ERPs. Brain Sciences, 13(7), 992. [Google Scholar] [CrossRef] [PubMed]
- Haatveit, B. C., Sundet, K., Hugdahl, K., Ueland, T., Melle, I., & Andreassen, O. A. (2010). The validity of d prime as a working memory index: Results from the “Bergen n-back task”. Journal of Clinical and Experimental Neuropsychology, 32(8), 871–880. [Google Scholar] [CrossRef]
- Hartung, J., Engelhardt, L. E., Thibodeaux, M. L., Harden, K. P., & Tucker-Drob, E. M. (2020). Developmental transformations in the structure of executive functions. Journal of Experimental Child Psychology, 189, 104681. [Google Scholar] [CrossRef]
- Heil, M., Osman, A., Wiegelmann, J., Rolke, B., & Hennighausen, E. (2000). N200 in the Eriksen-task: Inhibitory executive process? Journal of Psychophysiology, 14(4), 218. [Google Scholar] [CrossRef]
- Heinzel, S., Lorenz, R. C., Pelz, P., Heinz, A., Walter, H., Kathmann, N., & Stelzel, C. (2016). Neural correlates of training and transfer effects in working memory in older adults. Neuroimage, 134, 236–249. [Google Scholar] [CrossRef]
- Hermansen, T. K., Yrttiaho, S., Røysamb, E., & Melinder, A. (2017). Perceptual interference processing in preschool children, with and without prenatal exposure to selective serotonin reuptake inhibitors. Psychopharmacology, 234, 339–351. [Google Scholar] [CrossRef]
- Hoyniak, C. P., & Petersen, I. T. (2019). A meta-analytic evaluation of the N2 component as an endophenotype of response inhibition and externalizing psychopathology in childhood. Neuroscience & Biobehavioral Reviews, 103, 200–215. [Google Scholar]
- Huizinga, M., Dolan, C. V., & Van der Molen, M. W. (2006). Age-related change in executive function: Developmental trends and a latent variable analysis. Neuropsychologia, 44(11), 2017–2036. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, S. J., & Galletta, D. (2013). Event-rate effects in the flanker task: ERPs and task performance in children with and without AD/HD. International Journal of Psychophysiology, 87(3), 340–348. [Google Scholar] [CrossRef] [PubMed]
- Jones, J. S., Adlam, A. L. R., Benattayallah, A., & Milton, F. N. (2022). The neural correlates of working memory training in typically developing children. Child Development, 93(3), 815–830. [Google Scholar] [CrossRef]
- Juras, L., Sedlar, M. K., Vrhovski, G., & Vranic, A. (2024). N-back training in middle adulthood: Evidence for transfer only to structurally similar task. Current Psychology, 43(21), 19110–19120. [Google Scholar] [CrossRef]
- Karbach, J., & Kray, J. (2016). Plasticity of different cognitive domains: Executive functions. In T. Strobach, & J. Karbach (Eds.), Cognitive training: An overview of features and applications (pp. 93–103). Springer. [Google Scholar]
- Karbach, J., & Verhaeghen, P. (2014). Making working memory work: A meta-analysis of executive-control and working memory training in older adults. Psychological Science, 25(11), 2027–2037. [Google Scholar] [CrossRef]
- Kassai, R., Futo, J., Demetrovics, Z., & Takacs, Z. K. (2019). A meta-analysis of the experimental evidence on the near-and far-transfer effects among children’s executive function skills. Psychological Bulletin, 145(2), 165. [Google Scholar] [CrossRef]
- Kim, J., Walk, A. M., Keye, S. A., Kinder, C. J., Cannavale, C. N., Burd, N. A., & Khan, N. A. (2024). Adiposity influences intraindividual variability in behavioral and neuroelectric indices of attentional inhibition. Psychophysiology, 61(12), e14677. [Google Scholar] [CrossRef]
- Klingberg, T. (2010). Training and plasticity of working memory. Trends in Cognitive Sciences, 14(7), 317–324. [Google Scholar] [CrossRef]
- Kubota, M., Hadley, L. V., Schaeffner, S., Könen, T., Meaney, J. A., Morey, C. C., Auyeung, B., Moriguchi, Y., Karbach, J., & Chevalier, N. (2023). The effect of metacognitive executive function training on children’s executive function, proactive control, and academic skills. Developmental Psychology, 59(11), 2002. [Google Scholar] [CrossRef]
- McAlister, C., & Schmitter-Edgecombe, M. (2016). Executive function subcomponents and their relations to everyday functioning in healthy older adults. Journal of Clinical and Experimental Neuropsychology, 38(8), 925–940. [Google Scholar] [CrossRef] [PubMed]
- Melby-Lervåg, M., & Hulme, C. (2013). Is working memory training effective? A meta-analytic review. Developmental Psychology, 49(2), 270. [Google Scholar] [CrossRef]
- Melby-Lervåg, M., & Hulme, C. (2016). There is no convincing evidence that working memory training is effective: A reply to Au et al. (2014) and Karbach and Verhaeghen (2014). Psychonomic Bulletin & Review, 23, 324–330. [Google Scholar]
- Menu, I., Rezende, G., Le Stanc, L., Borst, G., & Cachia, A. (2022). Inhibitory control training on executive functions of children and adolescents: A latent change score model approach. Cognitive Development, 64, 101231. [Google Scholar] [CrossRef]
- Millner, A. J., Jaroszewski, A. C., Chamarthi, H., & Pizzagalli, D. A. (2012). Behavioral and electrophysiological correlates of training-induced cognitive control improvements. NeuroImage, 63(2), 742–753. [Google Scholar] [CrossRef]
- Minear, M., Brasher, F., Guerrero, C. B., Brasher, M., Moore, A., & Sukeena, J. (2016). A simultaneous examination of two forms of working memory training: Evidence for near transfer only. Memory & Cognition, 44, 1014–1037. [Google Scholar]
- Miyake, A., Friedman, N. P., Emerson, M. J., Witzki, A. H., Howerter, A., & Wager, T. D. (2000). The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cognitive Psychology, 41(1), 49–100. [Google Scholar] [CrossRef]
- Muñez, D., Lee, K., Bull, R., Khng, K. H., Cheam, F., & Rahim, R. A. (2022). Working memory and numeracy training for children with math learning difficulties: Evidence from a large-scale implementation in the classroom. Journal of Educational Psychology, 114(8), 1866. [Google Scholar] [CrossRef]
- Nelson, G., & McMaster, K. L. (2019). The effects of early numeracy interventions for students in preschool and early elementary: A meta-analysis. Journal of Educational Psychology, 111(6), 1001. [Google Scholar] [CrossRef]
- Ortiz-Rios, M., Agayby, B., Balezeau, F., Haag, M., Rima, S., Cadena-Valencia, J., & Schmid, M. C. (2023). Optogenetic stimulation of the primary visual cortex drives activity in the visual association cortex. Current Research in Neurobiology, 4, 100087. [Google Scholar] [CrossRef]
- Pereg, M., Shahar, N., & Meiran, N. (2013). Task switching training effects are mediated by working-memory management. Intelligence, 41(5), 467–478. [Google Scholar] [CrossRef]
- Pergher, V., Wittevrongel, B., Tournoy, J., Schoenmakers, B., & Van Hulle, M. M. (2018). N-back training and transfer effects revealed by behavioral responses and EEG. Brain and Behavior, 8(11), e01136. [Google Scholar] [CrossRef] [PubMed]
- Pronk, T., Hirst, R. J., Wiers, R. W., & Murre, J. M. (2023). Can we measure individual differences in cognitive measures reliably via smartphones? A comparison of the flanker effect across device types and samples. Behavior Research Methods, 55(4), 1641–1652. [Google Scholar] [CrossRef]
- Purpura, D. J., Schmitt, S. A., & Ganley, C. M. (2017). Foundations of mathematics and literacy: The role of executive functioning components. Journal of Experimental Child Psychology, 153, 15–34. [Google Scholar] [CrossRef]
- Raven, J., Raven, J. C., & Cour, J. H. (2000). Standard progressive matrices. Psychology Press. [Google Scholar]
- Rogers, R. D., & Monsell, S. (1995). Costs of a predictible switch between simple cognitive tasks. Journal of Experimental Psychology: General, 124(2), 207. [Google Scholar] [CrossRef]
- Röthlisberger, M., Neuenschwander, R., Cimeli, P., Michel, E., & Roebers, C. M. (2012). Improving executive functions in 5-and 6-year-olds: Evaluation of a small group intervention in prekindergarten and kindergarten children. Infant and Child Development, 21(4), 411–429. [Google Scholar] [CrossRef]
- Rueda, M. R., Checa, P., & Cómbita, L. M. (2012). Enhanced efficiency of the executive attention network after training in preschool children: Immediate changes and effects after two months. Developmental Cognitive Neuroscience, 2, S192–S204. [Google Scholar] [CrossRef]
- Rueda, M. R., Fan, J., McCandliss, B. D., Halparin, J. D., Gruber, D. B., Lercari, L. P., & Posner, M. I. (2004). Development of attentional networks in childhood. Neuropsychologia, 42(8), 1029–1040. [Google Scholar] [CrossRef]
- Sala, G., & Gobet, F. (2017). Working memory training in typically developing children: A meta-analysis of the available evidence. Developmental Psychology, 53(4), 671–685. [Google Scholar] [CrossRef]
- Sala, G., & Gobet, F. (2020). Working memory training in typically developing children: A multilevel meta-analysis. Psychonomic Bulletin and Review, 27(3), 423–434. [Google Scholar] [CrossRef]
- Salmi, J., Vilà-Balló, A., Soveri, A., Rostan, C., Rodríguez-Fornells, A., Lehtonen, M., & Laine, M. (2019). Working memory updating training modulates a cascade of event-related potentials depending on task load. Neurobiology of Learning and Memory, 166, 107085. [Google Scholar] [CrossRef] [PubMed]
- Salthouse, T. A., Fristoe, N., McGuthry, K. E., & Hambrick, D. Z. (1998). Relation of task switching to speed, age, and fluid intelligence. Psychology and Aging, 13(3), 445–461. [Google Scholar] [CrossRef]
- Sari, B. A., Koster, E. H., Pourtois, G., & Derakshan, N. (2016). Training working memory to improve attentional control in anxiety: A proof-of-principle study using behavioral and electrophysiological measures. Biological Psychology, 121, 203–212. [Google Scholar] [CrossRef]
- Schmiedek, F., Lövdén, M., & Lindenberger, U. (2010). Hundred days of cognitive training enhance broad cognitive abilities in adulthood: Findings from the COGITO study. Frontiers in Aging Neuroscience, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W., & Lockl, K. (2008). Procedural metacognition in children: Evidence for developmental trends. In J. Dunlosky, & R. A. Bjork (Eds.), Handbook of metamemory and memory (pp. 391–409). Psychology Press. [Google Scholar]
- Schneider, W., & Löffler, E. (2016). The development of metacognitive knowledge in children and adolescents. In J. Dunosky, & S. K. Tauber (Eds.), Oxford handbook of metamemory (pp. 491–518). Oxford University Press. [Google Scholar]
- Schwaighofer, M., Fischer, F., & Bühner, M. (2015). Does working memory training transfer? A meta-analysis including training conditions as moderators. Educational Psychologist, 50(2), 138–166. [Google Scholar] [CrossRef]
- Shipstead, Z., Redick, T. S., & Engle, R. W. (2012). Is working memory training effective? Psychological Bulletin, 138(4), 628. [Google Scholar] [CrossRef] [PubMed]
- Sluis, S. V. D., Jong, P. F. D., & Leij, A. V. D. (2007). Executive functioning in children, and its relations with reasoning, reading, and arithmetic. Intelligence, 35(5), 427–449. [Google Scholar] [CrossRef]
- Smortchkova, J., & Shea, N. (2020). Metacognitive development and conceptual change in children. Review of Philosophy and Psychology, 11(4), 745–763. [Google Scholar] [CrossRef]
- Soveri, A., Antfolk, J., Karlsson, L., Salo, B., & Laine, M. (2017). Working memory training revisited: A multi-level meta-analysis of n-back training studies. Psychonomic Bulletin & Review, 24, 1077–1096. [Google Scholar]
- Sprenger, A. M., Atkins, S. M., Bolger, D. J., Harbison, J. I., Novick, J. M., Chrabaszcz, J. S., Weems, S. A., Smith, V., Bobb, S., Bunting, M. F., & Dougherty, M. R. (2013). Training working memory: Limits of transfer. Intelligence, 41(5), 638–663. [Google Scholar] [CrossRef]
- Stahl, C., Voss, A., Schmitz, F., Nuszbaum, M., Tüscher, O., Lieb, K., & Klauer, K. C. (2014). Behavioral components of impulsivity. Journal of Experimental Psychology General, 143, 850–886. [Google Scholar] [CrossRef] [PubMed]
- Torres, R. E., Salahub, C., Campbell, K. L., & Emrich, S. (2024). Reduced distractor filtering with age: Evidence from the distractor positivity ERP. Journal of Vision, 24(10), 628. [Google Scholar] [CrossRef]
- Tusch, E. S., Alperin, B. R., Ryan, E., Holcomb, P. J., Mohammed, A. H., & Daffner, K. R. (2016). Changes in neural activity underlying working memory after computerized cognitive training in older adults. Frontiers in Aging Neuroscience, 8, 255. [Google Scholar] [CrossRef] [PubMed]
- Volckaert, A. M. S., & Noël, M. P. (2015). Training executive function in preschoolers reduce externalizing behaviors. Trends in Neuroscience and Education, 4(1–2), 37–47. [Google Scholar] [CrossRef]
- Von Bastian, C. C., & Oberauer, K. (2014). Effects and mechanisms of working memory training: A review. Psychological Research, 78, 803–820. [Google Scholar] [CrossRef] [PubMed]
- Wang, L., Sheng, A., Chang, L., & Zhou, R. (2022). Improving fluid intelligence of children through working memory training: The role of inhibition control. Frontiers in Psychology, 13, 1025036. [Google Scholar] [CrossRef]
- Xie, L., Ren, M., Cao, B., & Li, F. (2017). Distinct brain responses to different inhibitions: Evidence from a modified Flanker Task. Scientific Reports, 7(1), 6657. [Google Scholar] [CrossRef]
- Xu, F., Han, Y., Sabbagh, M. A., Wang, T., Ren, X., & Li, C. (2013). Developmental differences in the structure of executive function in middle childhood and adolescence. PLoS ONE, 8(10), e77770. [Google Scholar] [CrossRef]
- Xu, J., Deng, M., Nan, W., & Cai, D. (2021). The effects of working memory training in children revealed by behavioral responses and ERP. Brain and Behavior, 11(8), e2310. [Google Scholar] [CrossRef]
- Xu, P., Wu, D., Chen, Y., Wang, Z., & Xiao, W. (2020). The effect of response inhibition training on risky decision-making task performance. Frontiers in Psychology, 11, 1806. [Google Scholar] [CrossRef]
- Zarantonello, L., Brigadoi, S., Schiff, S., Bisiacchi, P. S., Cutini, S., Montagnese, S., & Amodio, P. (2024). Electrophysiological and hemodynamic mechanisms underlying load modulations in visuospatial working memory: A functional near-infrared spectroscopy (fNIRS) and electroencephalogram (EEG) study. Behavioral Neuroscience, 138(5), 331. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X., & Jia, L. (2019). Training and transfer effects of interference control training in children and young adults. Psychological Research, 83(7), 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X., Xu, Y., Fu, J., & Maes, J. H. (2018). Are training and transfer effects of working memory updating training modulated by achievement motivation? Memory & Cognition, 46, 398–409. [Google Scholar]
- Zhao, X., Zhou, R., & Fu, L. (2013). Working memory updating function training influenced brain activity. PLoS ONE, 8(8), e71063. [Google Scholar] [CrossRef]
- Zuber, S., Joly-Burra, E., Mahy, C. E., Loaiza, V., & Kliegel, M. (2023). Are facet-specific task trainings efficient in improving children’s executive functions and why (they might not be)? A multi-facet latent change score approach. Journal of Experimental Child Psychology, 227, 105602. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Training Group | Pretest | Posttest | ||
|---|---|---|---|---|
| M (SD) | Range | M (SD) | Range | |
| control | 1.42 (0.81) | 0.13–2.35 | 1.63 (0.79) | −0.41–3.45 |
| single | 1.14 (0.69) | 0.28–3.45 | 2.00 (0.96) | −0.20–3.56 |
| combined | 1.44 (0.43) | 0.13–3.45 | 2.26 (0.59) | −0.41–3.73 |
| Training Group | Pretest | Posttest | ||||||
|---|---|---|---|---|---|---|---|---|
| Congruent | Range | Incongruent | Range | Congruent | Rang | Incongruent | Range | |
| control | 477 (84) | 380–695 | 478 (126) | 431–974 | 451 (59) | 371–606 | 450 (91) | 415–816 |
| single | 512 (56) | 423–667 | 663 (123) | 521–1018 | 500 (85) | 401–771 | 590 (101) | 472–902 |
| combined | 515 (81) | 389–690 | 693 (136) | 442–934 | 455 (65) | 365–563 | 554 (99) | 431–773 |
| Training Group | Pretest | Posttest | ||||||
|---|---|---|---|---|---|---|---|---|
| Repeating | Switching | Repeating | Switching | |||||
| RT | ACC | RT | ACC | RT | ACC | RT | ACC | |
| control | 813 (27) | 0.80 (0.03) | 926 (30) | 0.63 (0.03) | 790 (21) | 0.83 (0.03) | 875 (25) | 0.71 (0.02) |
| single | 846 (21) | 0.74 (0.03) | 908 (25) | 0.61 (0.03) | 731 (31) | 0.75 (0.03) | 812 (40) | 0.65 (0.03) |
| combined | 866 (30) | 0.80 (0.04) | 1005 (42) | 0.59 (0.04) | 807 (33) | 0.82 (0.03) | 935 (39) | 0.67 (0.03) |
| Training Group | Pretest | Posttest | Tracking Test | |||
|---|---|---|---|---|---|---|
| M (SD) | Range | M (SD) | Range | M (SD) | Range | |
| control | −0.35 (0.46) | −0.92–0.50 | −0.22 (0.42) | −0.92–0.50 | −0.24 (0.69) | −1.34–1.13 |
| single | −0.30 (0.41) | −1–0.36 | −0.38 (0.37) | −1.17–0.31 | −0.20 (0.46) | −1–0.64 |
| combined | −0.38 (0.60) | −1.56–0.67 | −0.22 (0.50) | −1.08–0.67 | −0.22 (0.53) | −0.95–1.23 |
| Training Group | Pretest | Posttest | ||
|---|---|---|---|---|
| M (SD) | Range | M (SD) | Range | |
| control | 1.57 (0.64) | 0.13–2.42 | 1.69 (0.56) | 0.59–3.06 |
| combined | 1.46 (0.58) | 0.38–2.47 | 2.12 (0.42) | 1.56–3.11 |
| Training Group | Pretest | Posttest | ||||||
|---|---|---|---|---|---|---|---|---|
| Congruent | Range | Incongruent | Range | Congruent | Range | Incongruent | Range | |
| control | 468 (100) | 383–833 | 596 (116) | 461–914 | 456 (83) | 351–701 | 546 (102) | 377–797 |
| combined | 503 (83) | 388–690 | 669 (153) | 442–934 | 452 (62) | 365–560 | 533 (95) | 427–773 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, C.; Wang, B. Executive Function and Transfer Effect Training in Children: A Behavioral and Event-Related Potential Pilot Study. Behav. Sci. 2025, 15, 956. https://doi.org/10.3390/bs15070956
Cheng C, Wang B. Executive Function and Transfer Effect Training in Children: A Behavioral and Event-Related Potential Pilot Study. Behavioral Sciences. 2025; 15(7):956. https://doi.org/10.3390/bs15070956
Chicago/Turabian StyleCheng, Chen, and Baoxi Wang. 2025. "Executive Function and Transfer Effect Training in Children: A Behavioral and Event-Related Potential Pilot Study" Behavioral Sciences 15, no. 7: 956. https://doi.org/10.3390/bs15070956
APA StyleCheng, C., & Wang, B. (2025). Executive Function and Transfer Effect Training in Children: A Behavioral and Event-Related Potential Pilot Study. Behavioral Sciences, 15(7), 956. https://doi.org/10.3390/bs15070956