The 12-HHT/BLT2/NO Axis Is Associated to the Wound Healing and Skin Condition in Different Glycaemic States



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model of Type 2 Diabetes
2.2. Transepithelial/Transendothelial Electrical Resistance Measurement
2.3. 1-Hydroxyheptadecatrienoic Acid Quantification
2.4. Western Blot
2.5. Cell Culture
2.6. 2′,7′-dichlorodihydrofluorescein diacetate (DCF-2DA) Assay
2.7. Nitrites Colorimetric Measurement
2.8. In Vitro Wound Healing Assay
2.9. Reverse Transcription and Quantitative PCR
- Murine MMP9 (Mmp9)
- FW 5′-CCTACTCTGCCTGCACCACTAAA-3′
- RV 5′-CTGCTTGCCCAGGAAGACGAA-3′
- Human MMP9 (MMP9)
- FW 5′-GACGCAGACATCGTCATCCAGTTT-3′
- RV 5′-GCCGCGCCATCTGCGTTT-3′
- Murine iNOS (Nos2)
- FW 5′- CGAAACGCTTCACTTCCAA -3′
- RV 5′- TGAGCCTATATTGCTGTGGCT -3′
- Human iNOS (NOS2)
- FW 5′-5′-CAGCGGGATGACTTTCCAA-3′
- RV 5′-AGGCAAGATTTGGACCTGCA-3′
- Murine nNOS (Nos1)
- FW 5′- CTCACCCCGTCCTTTGAGTA-3′
- RV 5′- GGTCGCTTTGACTCTCTTGG-3′
- Human nNOS (NOS1)
- FW 5′- TCT CCT CCT ACT CTG ACT CC-3′
- RV 5′- TTG TGG ACA TTG GAT AGA CC-3′
- Murine BLT2) (Blt2)
- FW 5′-ACAGCCTTGGCTTTCTTCAG-3′
- RV 5′-TGCCCCATTACTTTCAGCTT-3′
- Murine β -actin (actb)
- FW 5′-CATCCGTAAAGACCTCTATGCCAAC-3′
- RV 5′-ATGGAGCCACCGATCCACA-3′
- Human β-actin (ACTB)
- FW 5′-TGGCACCCAGCACAATGAA-3′
- RV 5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′
2.10. Statistical Analysis
3. Results
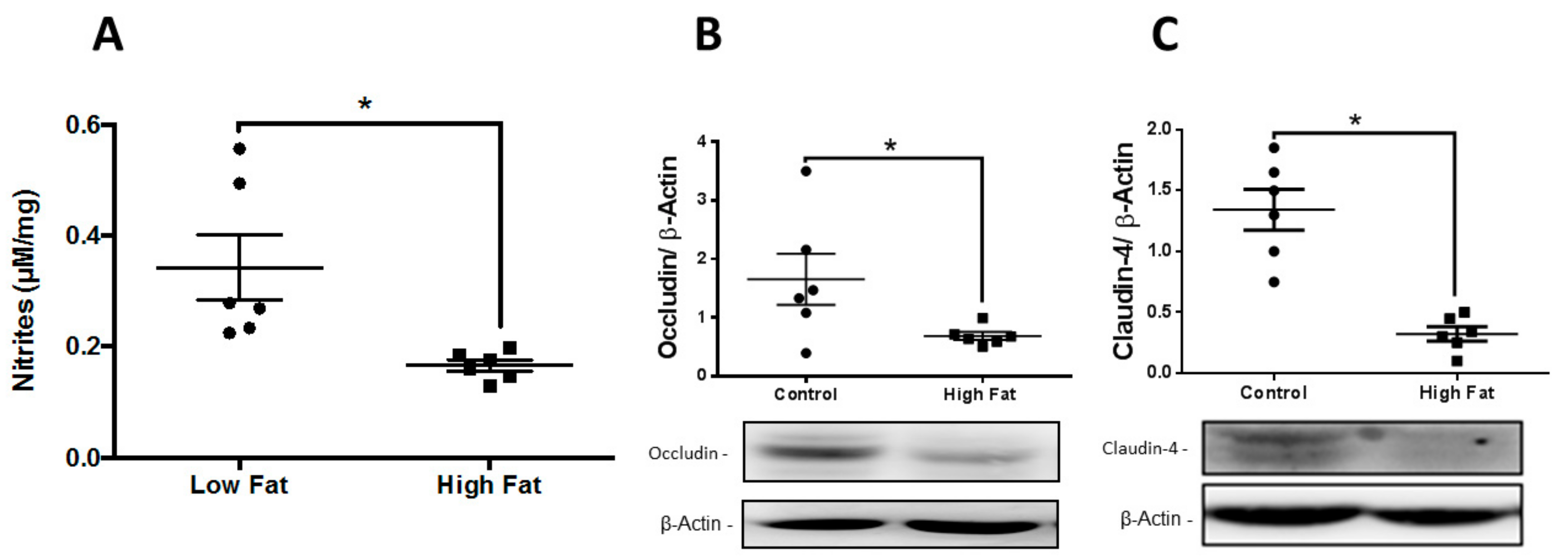
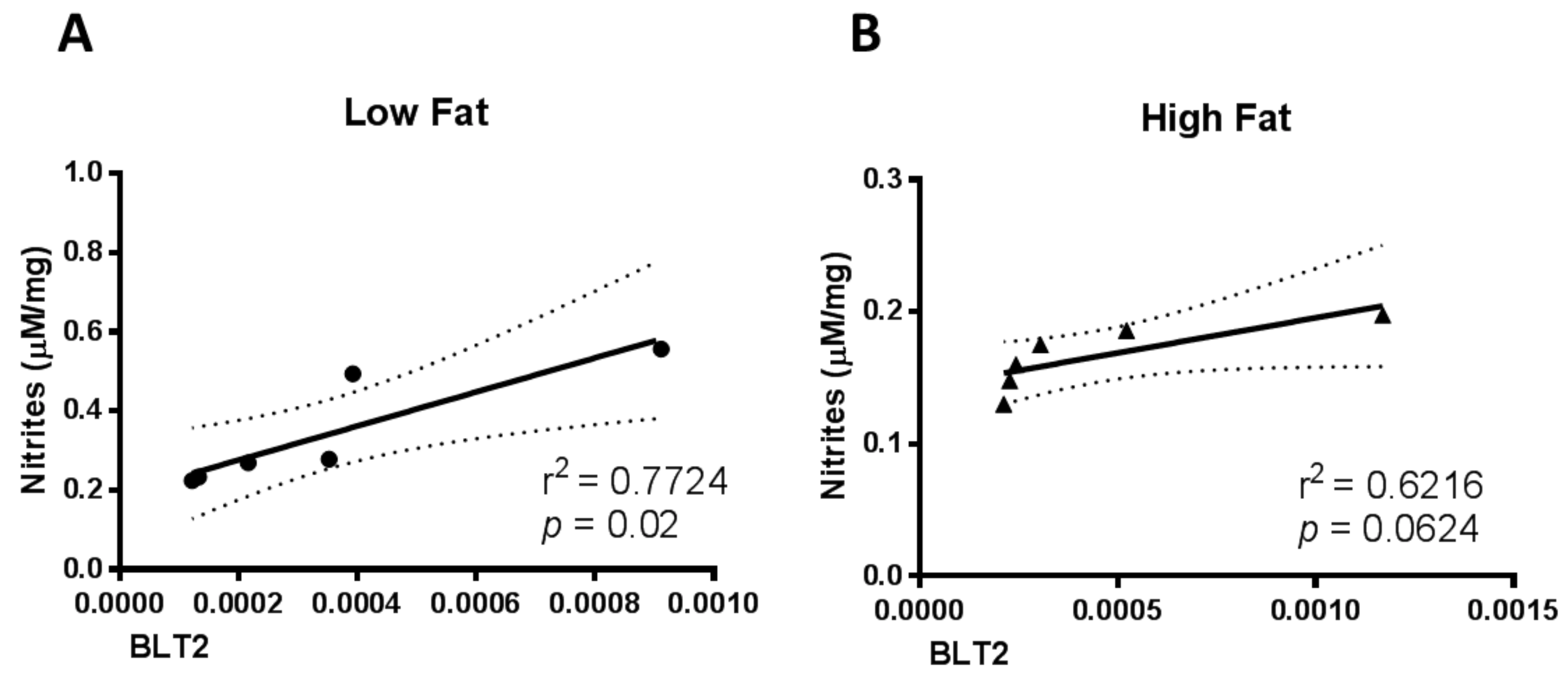
3.1. Skin Condition in T2D Animal Model
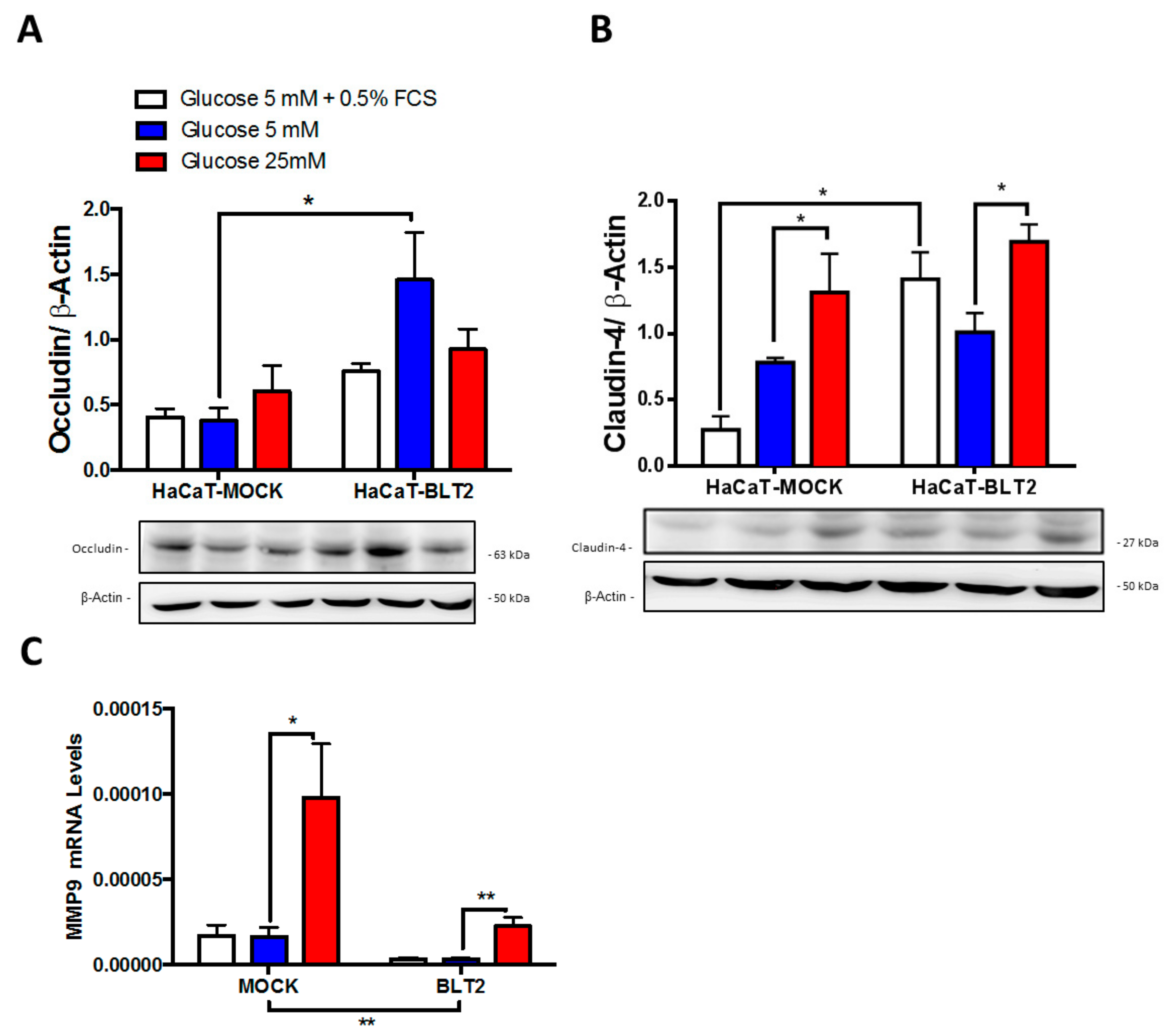
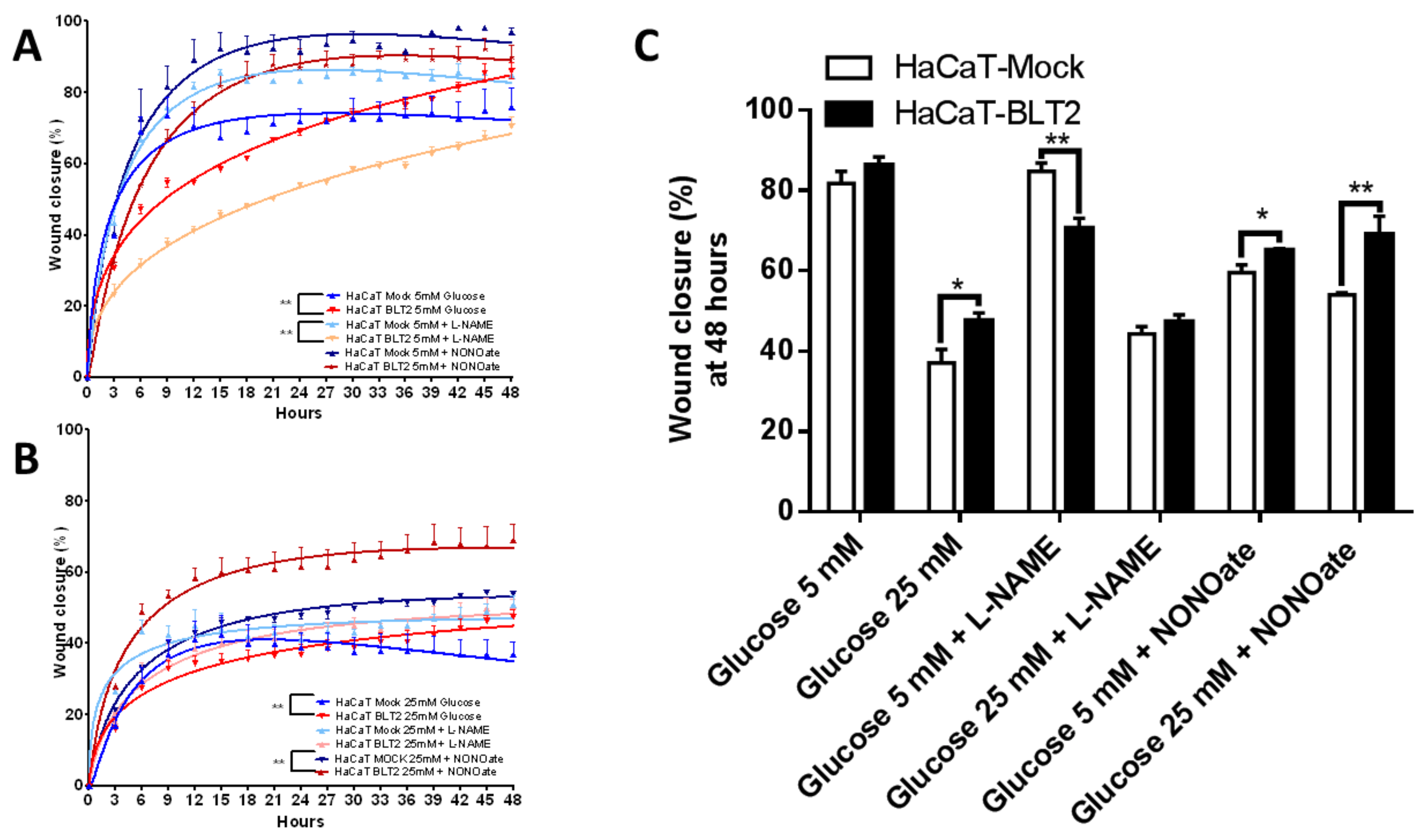
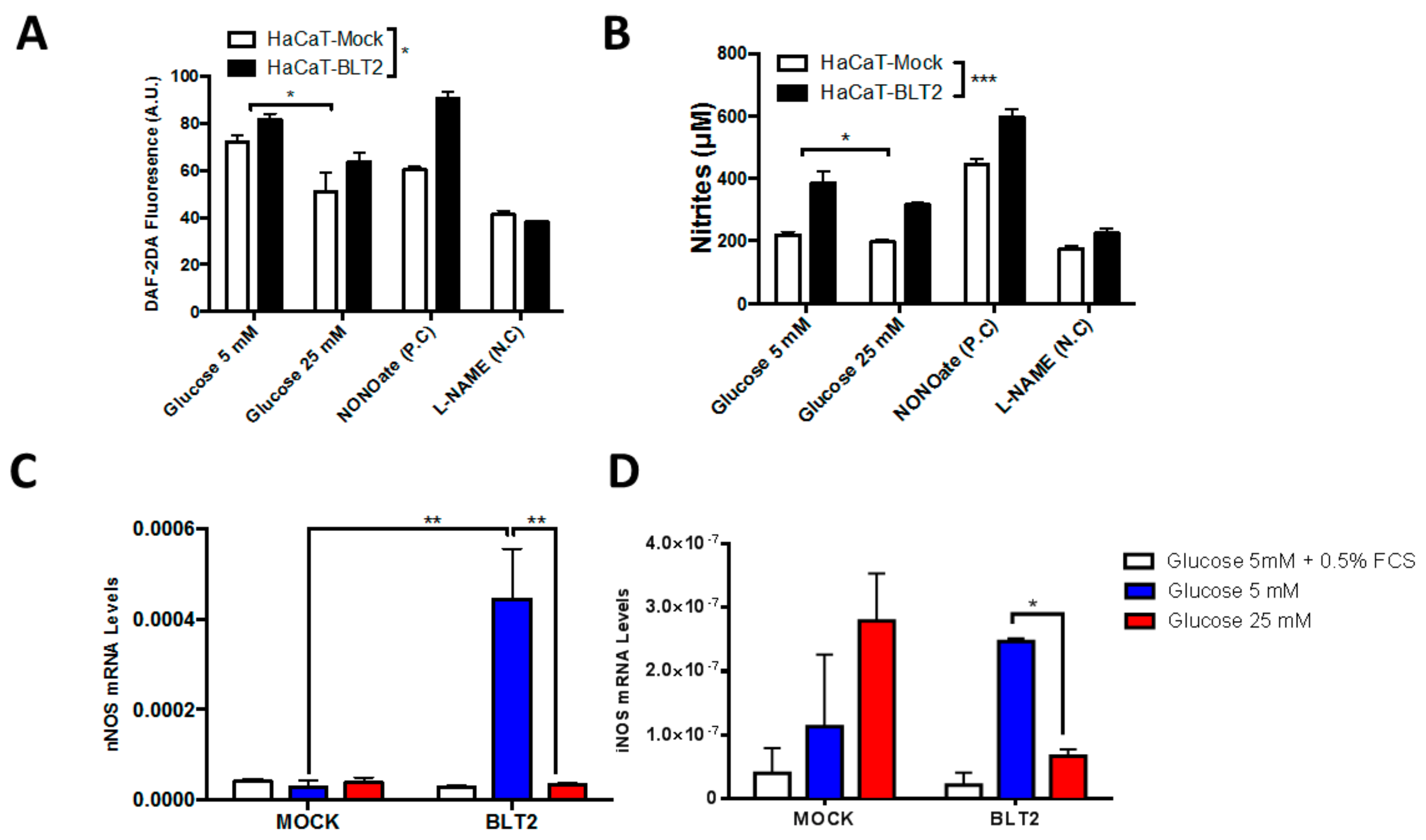
3.2. In Vitro Effect of High Glucose
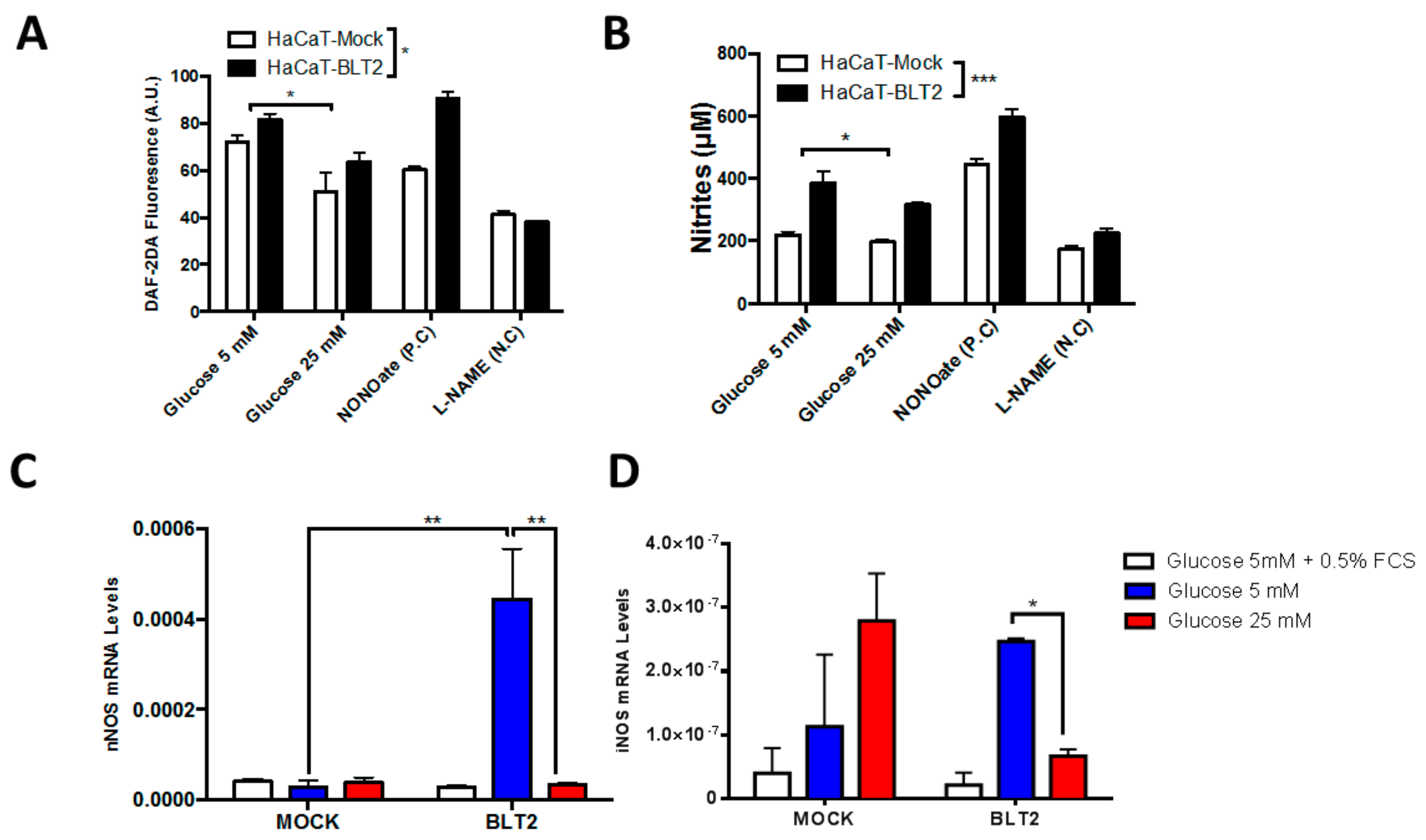
3.3. In Vitro Effect of NO Inhibition or Donation on Wound Healing
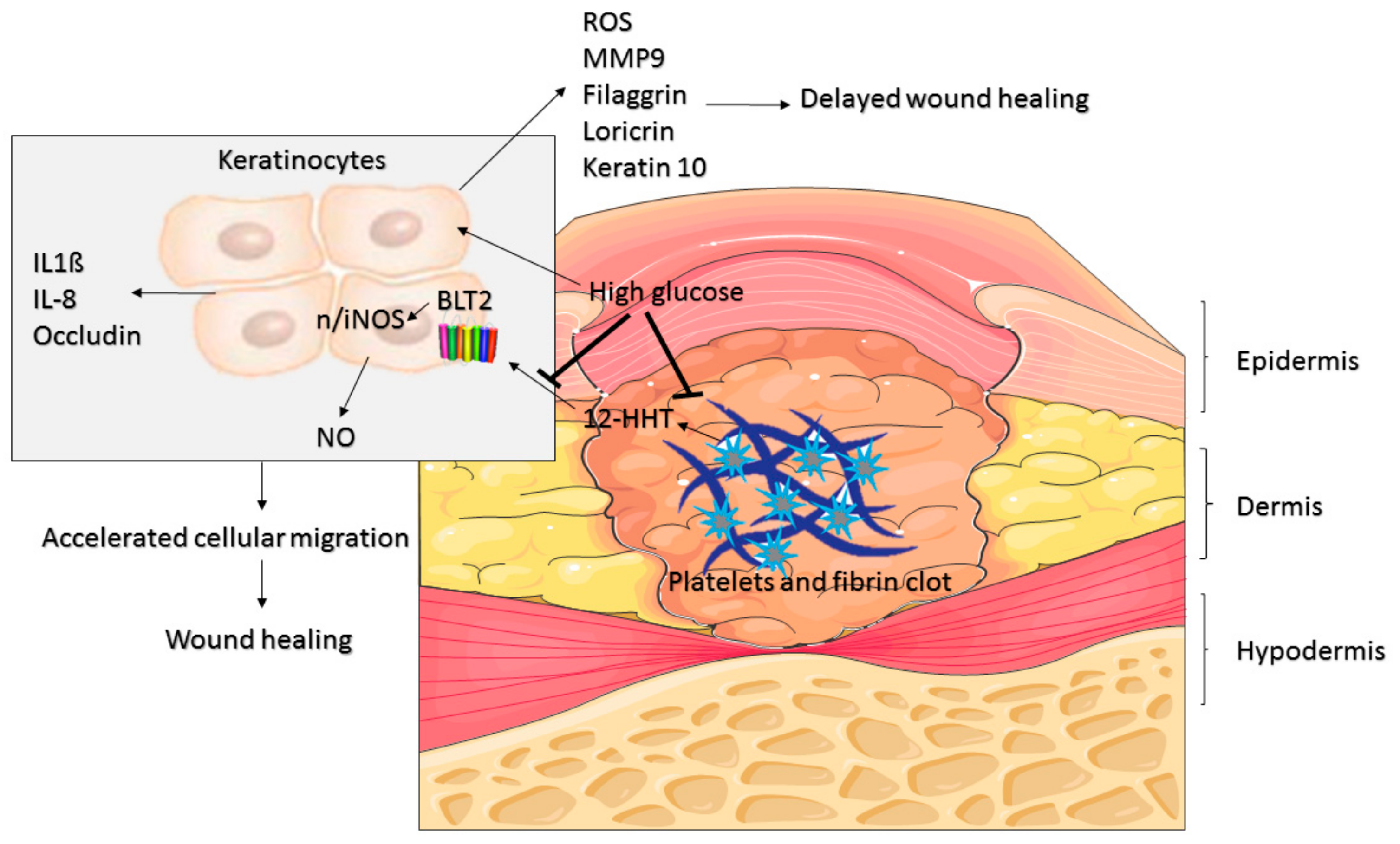
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Emerging Risk Factors Collaboration. Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease: A collaborative meta-analysis of 102 prospective studies. Lancet 2010, 375, 2215–2222. [Google Scholar] [CrossRef]
- Paneni, F.; Costantino, S.; Cosentino, F. Molecular mechanisms of vascular dysfunction and cardiovascular biomarkers in type 2 diabetes. Cardiovasc. Diagn. Ther. 2014, 4, 324–332. [Google Scholar] [CrossRef]
- Tsourdi, E.; Barthel, A.; Rietzsch, H.; Reichel, A.; Bornstein, S.R. Current aspects in the pathophysiology and treatment of chronic wounds in diabetes mellitus. BioMed Res. Int. 2013, 2013, 385641. [Google Scholar] [CrossRef]
- Li, P.; Oh, D.Y.; Bandyopadhyay, G.; Lagakos, W.S.; Talukdar, S.; Osborn, O.; Johnson, A.; Chung, H.; Maris, M.; Ofrecio, J.M.; et al. LTB4 promotes insulin resistance in obese mice by acting on macrophages, hepatocytes and myocytes. Nat. Med. 2015, 21, 239–247. [Google Scholar] [CrossRef]
- Liu, M.; Saeki, K.; Matsunobu, T.; Okuno, T.; Koga, T.; Sugimoto, Y.; Yokoyama, C.; Nakamizo, S.; Kabashima, K.; Narumiya, S.; et al. 12-Hydroxyheptadecatrienoic acid promotes epidermal wound healing by accelerating keratinocyte migration via the BLT2 receptor. J. Exp. Med. 2014, 211, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Kamohara, M.; Takasaki, J.; Matsumoto, M.; Saito, T.; Ohishi, T.; Ishii, H.; Furuichi, K. Molecular cloning and characterization of another leukotriene B4 receptor. J. Biol. Chem. 2000, 275, 27000–27004. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, T.; Kato, K.; Hagiya, H.; Izumi, T.; Shimizu, T. Hydroxyeicosanoids bind to and activate the low affinity leukotriene B4 receptor, BLT2. J. Biol. Chem. 2001, 276, 12454–12459. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, Y.; Yokomizo, T.; Terawaki, K.; Komine, M.; Tamaki, K.; Shimizu, T. Characterization of a mouse second leukotriene B4 receptor, mBLT2: BLT2-dependent ERK activation and cell migration of primary mouse keratinocytes. J. Biol. Chem. 2005, 280, 24816–24823. [Google Scholar] [CrossRef] [PubMed]
- Leguina-Ruzzi, A.; Valderas, J.P. BLT2 expression improves skin integrity and protects from alterations caused by hyperglycemia in type 2 diabetes. Dermato-Endocrinology 2016, 9, e1267078. [Google Scholar] [CrossRef] [PubMed]
- Leguina-Ruzzi, A.; Ortiz, R.; Velarde, V. The streptozotocin-high fat diet induced diabetic mouse model exhibits severe skin damage and alterations in local lipid mediators. Biomed. J. 2018, 41, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Volksdorf, T.; Heilmann, J.; Eming, S.A.; Schawjinski, K.; Zorn-Kruppa, M.; Ueck, C.; Vidal-Y-Sy, S.; Windhorst, S.; Jücker, M.; Moll, I.; et al. Tight Junction Proteins Claudin-1 and Occludin Are Important for Cutaneous Wound Healing. Am. J. Pathol. 2017, 187, 1301–1312. [Google Scholar] [CrossRef]
- Malone-Povolny, M.J.; Maloney, S.E.; Schoenfisch, M.H. Nitric Oxide Therapy for Diabetic Wound Healing. Adv. Healthc. Mater. 2019, e1801210. [Google Scholar] [CrossRef] [PubMed]
- Zhan, R.; Yang, S.; He, W.; Wang, F.; Tan, J.; Zhou, J.; Yang, S.; Yao, Z.; Wu, J.; Luo, G. Nitric oxide enhances keratinocyte cell migration by regulating Rho GTPase via cGMP-PKG signalling. PLoS ONE 2015, 10, e0121551. [Google Scholar] [CrossRef] [PubMed]
- Tessari, P.; Cecchet, D.; Cosma, A.; Vettore, M.; Coracina, A.; Millioni, R.; Iori, E.; Puricelli, L.; Avogaro, A.; Vedovato, M. Nitric oxide synthesis is reduced in subjects with type 2 diabetes and nephropathy. Diabetes 2010, 59, 2152–2159. [Google Scholar] [CrossRef] [PubMed]
- Kuo, I.H.; Carpenter-Mendini, A.; Yoshida, T.; McGirt, L.Y.; Ivanov, A.I.; Barnes, K.C.; Gallo, R.L.; Borkowski, A.W.; Yamasaki, K.; Leung, D.Y.; et al. Activation of epidermal toll-like receptor 2 enhances tight junction function: Implications for atopic dermatitis and skin barrier repair. J. Investig. Dermatol. 2013, 133, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Winzell, M.S.; Ahrén, B. The high-fat diet-fed mouse: A model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53 (Suppl. 3), S215–S219. [Google Scholar] [CrossRef]
- King, A.J. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.R.; Fu, Z.; Liu, D. Development of a nongenetic mouse model of type 2 diabetes. Exp. Diabetes Res. 2011, 2011, 416254. [Google Scholar] [CrossRef]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef]
- Reiss, M.J.; Han, Y.P.; Garcia, E.; Goldberg, M.; Yu, H.; Garner, W.L. Matrix metalloproteinase-9 delays wound healing in a murine wound model. Surgery 2010, 147, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Campos, T.M.; Passos, S.T.; Novais, F.O.; Beiting, D.P.; Costa, R.S.; Queiroz, A.; Mosser, D.; Scott, P.; Carvalho, E.M.; Carvalho, L.P. Matrix metalloproteinase 9 production by monocytes is enhanced by TNF and participates in the pathology of human cutaneous Leishmaniasis. PLoS Negl. Trop. Dis. 2014, 8, e3282. [Google Scholar] [CrossRef]
- Brandner, J.M.; Zorn-Kruppa, M.; Yoshida, T.; Moll, I.; Beck, L.A.; De Benedetto, A. Epidermal tight junctions in health and disease. Tissue Barriers 2015, 3, e974451. [Google Scholar] [CrossRef] [PubMed]
- Guth, K.; Schäfer-Korting, M.; Fabian, E.; Landsiedel, R.; van Ravenzwaay, B. Suitability of skin integrity tests for dermal absorption studies in vitro. Toxicol. In Vitro 2015, 29, 113–123. [Google Scholar] [CrossRef]
- Luiking, Y.C.; Engelen, M.P.; Deutz, N.E. Regulation of nitric oxide production in health and disease. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 97–104. [Google Scholar] [CrossRef]
- Adler, B.L.; Friedman, A.J. Nitric oxide therapy for dermatologic disease. Future Sci. OA 2015, 1, FSO37. [Google Scholar] [CrossRef] [PubMed]
- Cals-Grierson, M.M.; Ormerod, A.D. Nitric oxide function in the skin. Nitric Oxide 2004, 10, 179–193. [Google Scholar] [CrossRef]
- Paulsen, S.M.; Wurster, S.H.; Nanney, L.B. Expression of inducible nitric oxide synthase in human burn wounds. Wound Repair Regen. 1998, 6, 142–148. [Google Scholar] [CrossRef]
- Amadeu, T.P.; Costa, A.M. Nitric oxide synthesis inhibition alters rat cutaneous wound healing. J. Cutan. Pathol. 2006, 33, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Kutner, A.; Adam Friedman, A. Nitric oxide nanoparticles for wound healing: Future directions to overcome challenges. Expert Rev. Dermatol. 2013, 8, 451–461. [Google Scholar] [CrossRef]
- Shabani, M.; Pulfer, S.K.; Bulgrin, J.P.; Smith, D.J. Enhancement of wound repair with a topically applied nitric oxide-releasing polymer. Wound Repair Regen. 1996, 4, 353–362. [Google Scholar] [CrossRef]
- Qiu, H.; Johansson, A.S.; Sjöström, M.; Wan, M.; Schröder, O.; Palmblad, J.; Haeggström, J.Z. Differential induction of BLT receptor expression on human endothelial cells by lipopolysaccharide, cytokines, and leukotriene B4. Proc. Natl. Acad. Sci. USA 2006, 103, 6913–6918. [Google Scholar] [CrossRef] [PubMed]
- Leguina-Ruzzi, A.; Pereira, J.; Pereira-Flores, K.; Valderas, J.P.; Mezzano, D.; Velarde, V.; Sáez, C.G. Increased RhoA/Rho-Kinase Activity and Markers of Endothelial Dysfunction in Young Adult Subjects with Metabolic Syndrome. Metab. Syndr. Relat. Disord. 2015, 13, 373–380. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical and Biochemical Characteristics | ||
|---|---|---|
| Low Fat | High Fat | |
| Weight (g) | 26.88 ± 0.44 | 34.35 ± 0.61 ** |
| Glucose (mg/dL) | 111.7 ± 2.21 | 225 ± 7.13 ** |
| Insulin (µg/L) | 1.41 ± 0.10 | 8.04 ± 2.38 * |
| Triglycerides (mg/dL) | 156.3 ± 2.23 | 192 ± 6.63 ** |
| Skin Characteristics | ||
| TEER (Ωcm2) | 23.70 ± 1.30 | 11.30 ± 2.37 ** |
| 12-HHT (pg/mg) | 15.22 ± 6.11 | 6.73 ± 2.35 * |
| iNOS mRNA (A.U) | 4.47 × 10−6 ± 6.58 × 10−7 | 0.48 × 10−6 ± 2.68 × 10−7 ** |
| nNOS mRNA (A.U) | 4.8 × 10−4 ± 1.3 × 10−4 | 5.2 × 10−4 ± 1.5 × 10−4 |
| MMP9 mRNA (A.U) | 0.002 ± 0.0009 | 0.013 ± 0.0025 ** |
| BLT2 mRNA (A.U) | 3.5 × 10−4 ± 1.2 × 10−4 | 4.4 × 10−4 ± 1.5 × 10−4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leguina-Ruzzi, A.; Diban, R.O.; Velarde, V. The 12-HHT/BLT2/NO Axis Is Associated to the Wound Healing and Skin Condition in Different Glycaemic States. Med. Sci. 2019, 7, 65. https://doi.org/10.3390/medsci7040065
Leguina-Ruzzi A, Diban RO, Velarde V. The 12-HHT/BLT2/NO Axis Is Associated to the Wound Healing and Skin Condition in Different Glycaemic States. Medical Sciences. 2019; 7(4):65. https://doi.org/10.3390/medsci7040065
Chicago/Turabian StyleLeguina-Ruzzi, Alberto, Rina Ortiz Diban, and Victoria Velarde. 2019. "The 12-HHT/BLT2/NO Axis Is Associated to the Wound Healing and Skin Condition in Different Glycaemic States" Medical Sciences 7, no. 4: 65. https://doi.org/10.3390/medsci7040065
APA StyleLeguina-Ruzzi, A., Diban, R. O., & Velarde, V. (2019). The 12-HHT/BLT2/NO Axis Is Associated to the Wound Healing and Skin Condition in Different Glycaemic States. Medical Sciences, 7(4), 65. https://doi.org/10.3390/medsci7040065