Role of Polyamines in Immune Cell Functions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. B-Cell Lymphopoiesis and Activation
3. The Role of Polyamines in T-Lymphopoiesis
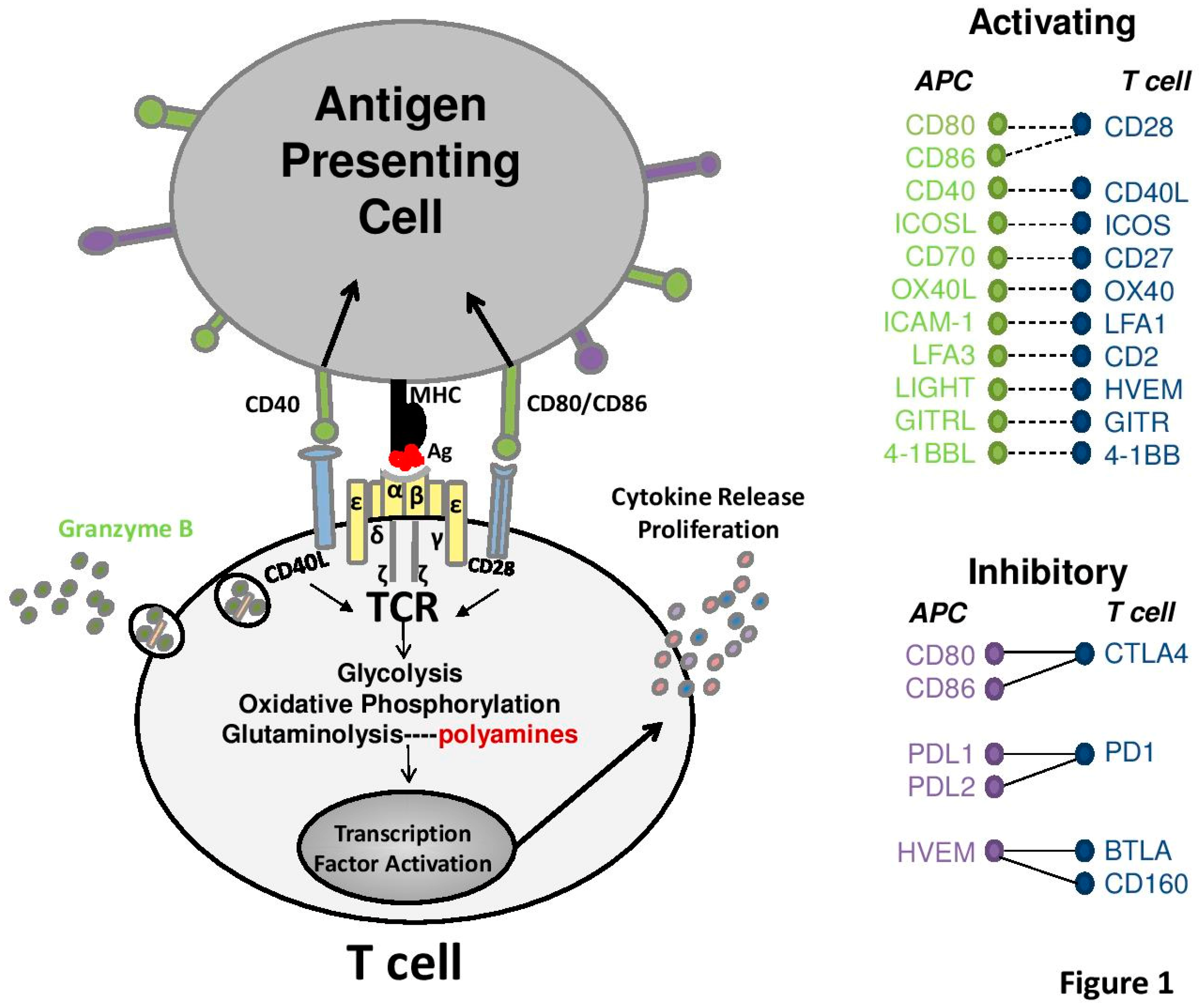
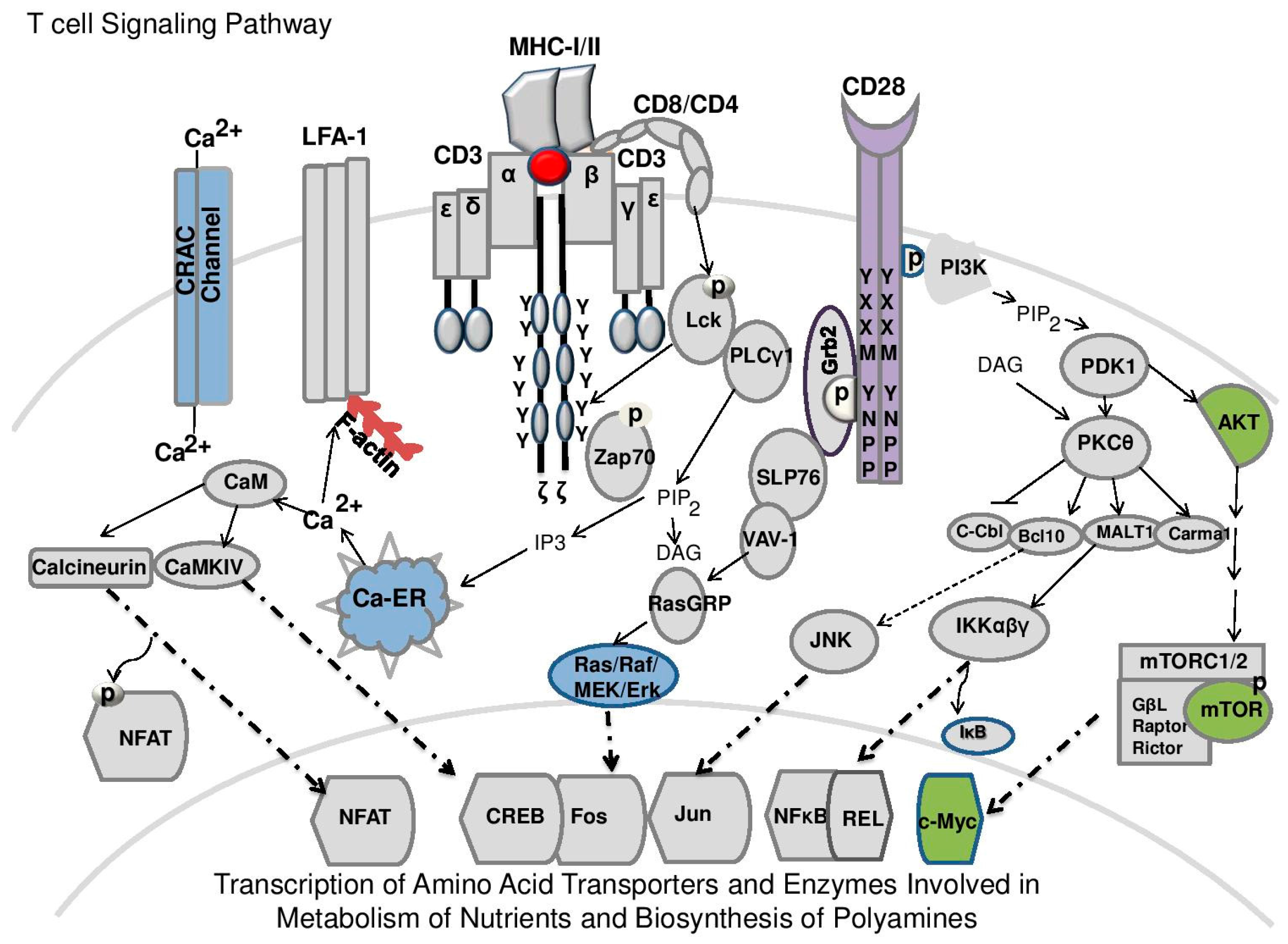
4. Role of Polyamines in Antigen Activated T-Cells
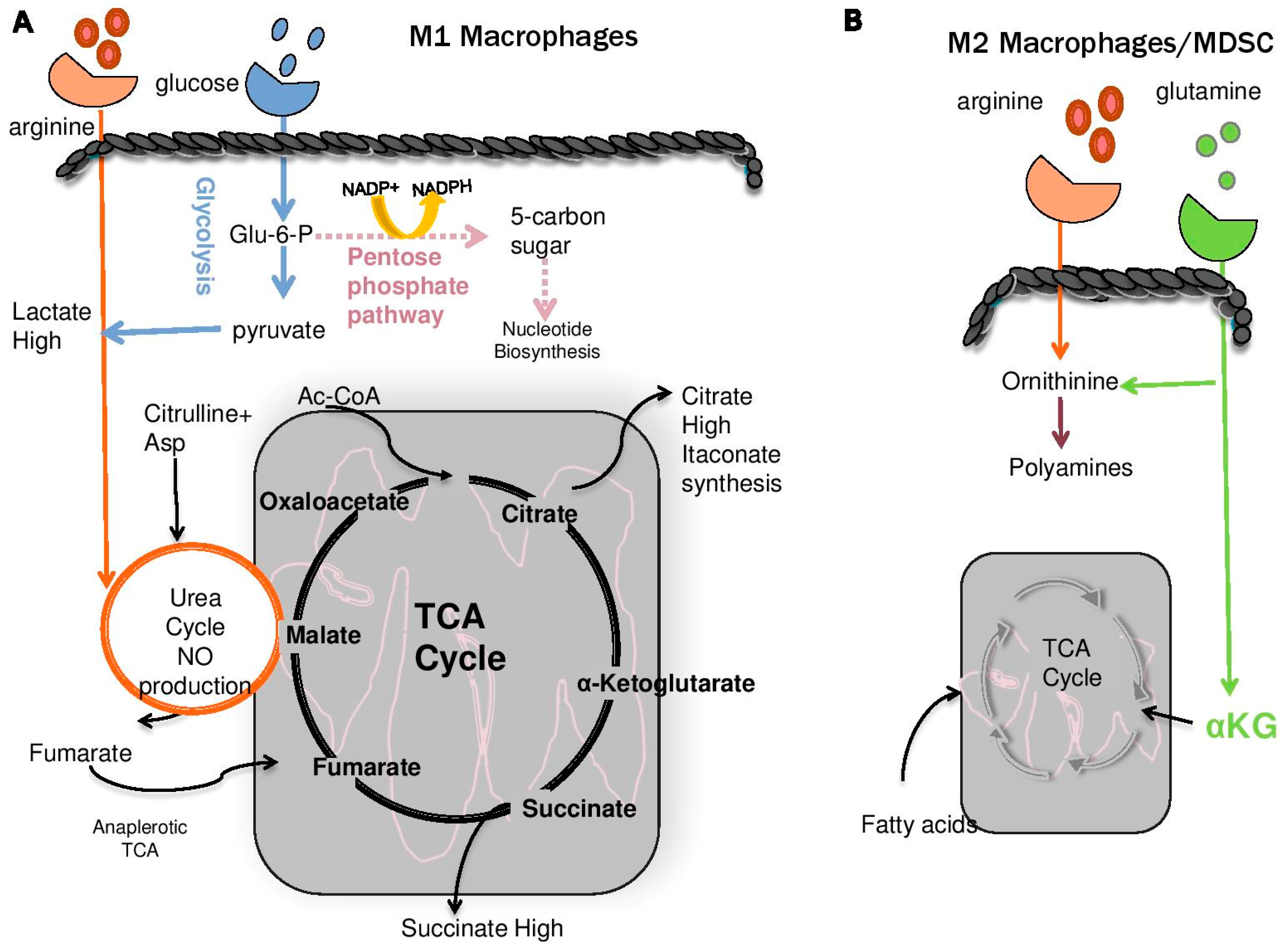
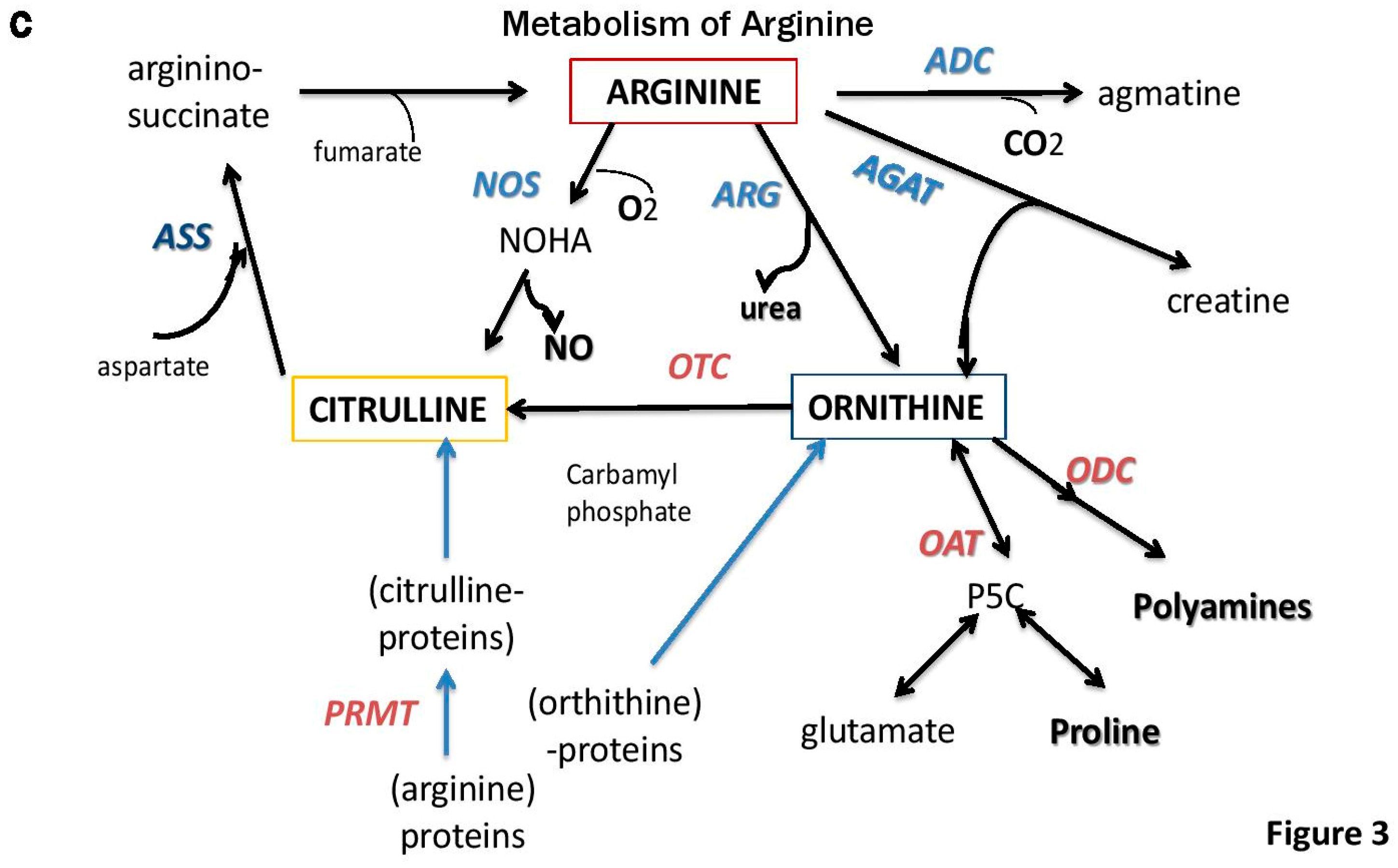
5. Role of Polyamines and Anti-Tumor Immunity
6. Polyamines in Autoimmune Disease
7. Concluding Remarks
Author Contributions
Conflicts of Interest
References
- Green, D.R. Metabolism and immunity: The old and the new. Semin. Immunol. 2012, 24, 383. [Google Scholar] [CrossRef] [PubMed]
- Halligan, D.N.; Murphy, S.J.; Taylor, C.T. The hypoxia-inducible factor (HIF) couples immunity with metabolism. Semin. Immunol. 2016, 28, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Scharping, N.E.; Delgoffe, G.M. Tumor Microenvironment Metabolism: A New Checkpoint for Anti-Tumor Immunity. Vaccines (Basel) 2016, 4, 46. [Google Scholar] [CrossRef] [PubMed]
- Pearce, E.L.; Pearce, E.J. Metabolic pathways in immune cell activation and quiescence. Immunity 2013, 38, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Michalek, R.D.; Rathmell, J.C. The metabolic life and times of a T-cell. Immunol. Rev. 2010, 236, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Gerriets, V.A.; Rathmell, J.C. Metabolic pathways in T cell fate and function. Trends Immunol. 2012, 33, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Olenchock, B.A.; Rathmell, J.C.; Vander Heiden, M.G. Biochemical Underpinnings of Immune Cell Metab.olic Phenotypes. Immunity 2017, 46, 703–713. [Google Scholar] [CrossRef] [PubMed]
- O‘Neill, L.A.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Goodnow, C.C.; Sprent, J.; Fazekas de St Groth, B.; Vinuesa, C.G. Cellular and genetic mechanisms of self tolerance and autoimmunity. Nature 2005, 435, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Flajnik, M.F.; Kasahara, M. Origin and evolution of the adaptive immune system: Genetic events and selective pressures. Nat. Rev. Genet. 2010, 11, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Weissman, I.L.; Akashi, K. Identification of clonogenic common lymphoid progenitors in mouse bone marrow. Cell 1997, 91, 661–672. [Google Scholar] [CrossRef]
- Akashi, K.; Traver, D.; Kondo, M.; Weissman, I.L. Lymphoid development from hematopoietic stem cells. Int. J. Hematol. 1999, 69, 217–226. [Google Scholar] [PubMed]
- Akashi, K.; Traver, D.; Miyamoto, T.; Weissman, I.L. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature 2000, 404, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Hemmati, H.D.; Wandycz, A.M.; Weissman, I.L. The purification and characterization of fetal liver hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 1995, 92, 10302–10306. [Google Scholar] [CrossRef] [PubMed]
- Venkitaraman, A.R.; Williams, G.T.; Dariavach, P.; Neuberger, M.S. The B-cell antigen receptor of the five immunoglobulin classes. Nature 1991, 352, 777–781. [Google Scholar] [CrossRef] [PubMed]
- Pape, K.A.; Catron, D.M.; Itano, A.A.; Jenkins, M.K. The humoral immune response is initiated in lymph nodes by B cells that acquire soluble antigen directly in the follicles. Immunity 2007, 26, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Tonegawa, S. Somatic generation of antibody diversity. Nature 1983, 302, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Garside, P.; Ingulli, E.; Merica, R.R.; Johnson, J.G.; Noelle, R.J.; Jenkins, M.K. Visualization of specific B and T lymphocyte interactions in the lymph node. Science 1998, 281, 96–99. [Google Scholar] [CrossRef] [PubMed]
- McAdam, A.J.; Greenwald, R.J.; Levin, M.A.; Chernova, T.; Malenkovich, N.; Ling, V.; Freeman, G.J.; Sharpe, A.H. ICOS is critical for CD40-mediated antibody class switching. Nature 2001, 409, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Honjo, T.; Kinoshita, K.; Muramatsu, M. Molecular mechanism of class switch recombination: Linkage with somatic hypermutation. Annu. Rev. Immunol. 2002, 20, 165–196. [Google Scholar] [CrossRef] [PubMed]
- Gong, S.; Nussenzweig, M.C. Regulation of an early developmental checkpoint in the B cell pathway by Ig β. Science 1996, 272, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Caro-Maldonado, A.; Wang, R.; Nichols, A.G.; Kuraoka, M.; Milasta, S.; Sun, L.D.; Gavin, A.L.; Abel, E.D.; Kelsoe, G.; Green, D.R.; et al. Metabolic reprogramming is required for antibody production that is suppressed in anergic but exaggerated in chronically BAFF-exposed B cells. J. Immunol. 2014, 192, 3626–3636. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Pearce, E.J. MenTORing Immunity: mTOR Signaling in the Development and Function of Tissue-Resident Immune Cells. Immunity 2017, 46, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Siska, P.J.; van der Windt, G.J.; Kishton, R.J.; Cohen, S.; Eisner, W.; MacIver, N.J.; Kater, A.P.; Weinberg, J.B.; Rathmell, J.C. Suppression of Glut1 and Glucose Metabolism by Decreased Akt/mTORC1 Signaling Drives T Cell Impairment in B Cell Leukemia. J. Immunol. 2016, 197, 2532–2540. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, J.A.; Keller, U.B.; Baudino, T.A.; Yang, C.; Norton, S.; Old, J.A.; Nilsson, L.M.; Neale, G.; Kramer, D.L.; Porter, C.W.; et al. Targeting ornithine decarboxylase in Myc-induced lymphomagenesis prevents tumor formation. Cancer Cell 2005, 7, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Bello-Fernandez, C.; Packham, G.; Cleveland, J.L. The ornithine decarboxylase gene is a transcriptional target of c-Myc. Proc. Natl. Acad. Sci. USA 1993, 90, 7804–7808. [Google Scholar] [CrossRef] [PubMed]
- Tessem, M.B.; Bertilsson, H.; Angelsen, A.; Bathen, T.F.; Drablos, F.; Rye, M.B. A Balanced Tissue Composition Reveals New Metabolic and Gene Expression Markers in Prostate Cancer. PLoS ONE 2016, 11, e0153727. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Regulation of ornithine decarboxylase. J. Biol. Chem. 2006, 281, 14529–14532. [Google Scholar] [CrossRef] [PubMed]
- Pendeville, H.; Carpino, N.; Marine, J.C.; Takahashi, Y.; Muller, M.; Martial, J.A.; Cleveland, J.L. The ornithine decarboxylase gene is essential for cell survival during early murine development. Mol. Cell. Biol. 2001, 21, 6549–6558. [Google Scholar] [CrossRef] [PubMed]
- Nitta, T.; Igarashi, K.; Yamashita, A.; Yamamoto, M.; Yamamoto, N. Involvement of polyamines in B cell receptor-mediated apoptosis: Spermine functions as a negative modulator. Exp. Cell Res. 2001, 265, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, H.; Egusa, H.; Ueura, N.; Matsumoto, Y.; Kanayama, N.; Hikida, M. Selective augmenting effects of nitric oxide on antigen-specific IgE response in mice. Immunopharmacology 2000, 46, 55–63. [Google Scholar] [CrossRef]
- Kramer, D.L.; Diegelman, P.; Jell, J.; Vujcic, S.; Merali, S.; Porter, C.W. Polyamine acetylation modulates polyamine metabolic flux, a prelude to broader metabolic consequences. J. Biol. Chem. 2008, 283, 4241–4251. [Google Scholar] [CrossRef] [PubMed]
- Akamatsu, Y.; Monroe, R.; Dudley, D.D.; Elkin, S.K.; Gartner, F.; Talukder, S.R.; Takahama, Y.; Alt, F.W.; Bassing, C.H.; Oettinger, M.A. Deletion of the RAG2 C terminus leads to impaired lymphoid development in mice. Proc. Natl. Acad. Sci. USA 2003, 100, 1209–1214. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P.; Iacomini, J.; Johnson, R.S.; Herrup, K.; Tonegawa, S.; Papaioannou, V.E. RAG-1-deficient mice have no mature B and T lymphocytes. Cell 1992, 68, 869–877. [Google Scholar] [CrossRef]
- Zhu, C.; Roth, D.B. Characterization of coding ends in thymocytes of scid mice: Implications for the mechanism of V(D)J recombination. Immunity 1995, 2, 101–112. [Google Scholar] [CrossRef][Green Version]
- Zlotoff, D.A.; Sambandam, A.; Logan, T.D.; Bell, J.J.; Schwarz, B.A.; Bhandoola, A. CCR7 and CCR9 together recruit hematopoietic progenitors to the adult thymus. Blood 2010, 115, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Zlotoff, D.A.; Zhang, S.L.; De Obaldia, M.E.; Hess, P.R.; Todd, S.P.; Logan, T.D.; Bhandoola, A. Delivery of progenitors to the thymus limits T-lineage reconstitution after bone marrow transplantation. Blood 2011, 118, 1962–1970. [Google Scholar] [CrossRef] [PubMed]
- Meredith, M.; Zemmour, D.; Mathis, D.; Benoist, C. Aire controls gene expression in the thymic epithelium with ordered stochasticity. Nat. Immunol. 2015, 16, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Wolfer, A.; Wilson, A.; Nemir, M.; MacDonald, H.R.; Radtke, F. Inactivation of Notch1 impairs VDJβ rearrangement and allows pre-TCR-independent survival of early αβ Lineage Thymocytes. Immunity 2002, 16, 869–879. [Google Scholar] [CrossRef]
- Tanigaki, K.; Tsuji, M.; Yamamoto, N.; Han, H.; Tsukada, J.; Inoue, H.; Kubo, M.; Honjo, T. Regulation of αβ/γδ T cell lineage commitment and peripheral T cell responses by Notch/RBP-J signaling. Immunity 2004, 20, 611–622. [Google Scholar] [CrossRef]
- Mombaerts, P.; Clarke, A.R.; Rudnicki, M.A.; Iacomini, J.; Itohara, S.; Lafaille, J.J.; Wang, L.; Ichikawa, Y.; Jaenisch, R.; Hooper, M.L.; et al. Mutations in T-cell antigen receptor genes α and β block thymocyte development at different stages. Nature 1992, 360, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Haas, W.; Pereira, P.; Tonegawa, S. Gamma/delta cells. Annu. Rev. Immunol. 1993, 11, 637–685. [Google Scholar] [CrossRef] [PubMed]
- Germain, R.N. T-cell development and the CD4-CD8 lineage decision. Nat. Rev. Immunol. 2002, 2, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.F.; Tormo, J.; Gerth, U.C.; Wyer, J.R.; McMichael, A.J.; Stuart, D.I.; Bell, J.I.; Jones, E.Y.; Jakobsen, B.K. Crystal structure of the complex between human CD8αα and HLA-A2. Nature 1997, 387, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Josefowicz, S.Z.; Rudensky, A. Control of regulatory T cell lineage commitment and maintenance. Immunity 2009, 30, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Bendelac, A.; Savage, P.B.; Teyton, L. The biology of NKT cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [PubMed]
- McDonald, B.D.; Bunker, J.J.; Ishizuka, I.E.; Jabri, B.; Bendelac, A. Elevated T cell receptor signaling identifies a thymic precursor to the TCRαβ+ CD4−CD8β− intraepithelial lymphocyte lineage. Immunity 2014, 41, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Cheroutre, H. IELs: Enforcing law and order in the court of the intestinal epithelium. Immunol. Rev. 2005, 206, 114–131. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Gupta, K.; Pisarev, V.; Kinarsky, L.; Sherman, S.; Kang, L.; Herber, D.L.; Schneck, J.; Gabrilovich, D.I. Altered recognition of antigen is a mechanism of CD8+ T cell tolerance in cancer. Nat. Med. 2007, 13, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Bertram, E.M.; Dawicki, W.; Watts, T.H. Role of T cell costimulation in anti-viral immunity. Semin. Immunol. 2004, 16, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Croft, M. Costimulation of T cells by OX40, 4-1BB, and CD27. Cytokine Growth Factor Rev. 2003, 14, 265–273. [Google Scholar] [CrossRef]
- Gramaglia, I.; Weinberg, A.D.; Lemon, M.; Croft, M. Ox-40 ligand: A potent costimulatory molecule for sustaining primary CD4 T cell responses. J. Immunol. 1998, 161, 6510–6517. [Google Scholar] [PubMed]
- Rogers, P.R.; Croft, M. CD28, Ox-40, LFA-1, and CD4 modulation of Th1/Th2 differentiation is directly dependent on the dose of antigen. J. Immunol. 2000, 164, 2955–2963. [Google Scholar] [CrossRef] [PubMed]
- Cantrell, D.A. Transgenic analysis of thymocyte signal transduction. Nat. Rev. Immunol. 2002, 2, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Derbinski, J.; Schulte, A.; Kyewski, B.; Klein, L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat. Immunol. 2001, 2, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Giraud, M.; Benoist, C.; Mathis, D. Aire‘s partners in the molecular control of immunological tolerance. Cell 2010, 140, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Danan-Gotthold, M.; Guyon, C.; Giraud, M.; Levanon, E.Y.; Abramson, J. Extensive RNA editing and splicing increase immune self-representation diversity in medullary thymic epithelial cells. Genome Biol. 2016, 17, 219. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.S.; Kingston, R.E.; Benoist, C.; Mathis, D. Global relevance of Aire binding to hypomethylated lysine-4 of histone-3. Proc. Natl. Acad. Sci. USA 2010, 107, 13016–13021. [Google Scholar] [CrossRef] [PubMed]
- Giraud, M.; Yoshida, H.; Abramson, J.; Rahl, P.B.; Young, R.A.; Mathis, D.; Benoist, C. Aire unleashes stalled RNA polymerase to induce ectopic gene expression in thymic epithelial cells. Proc. Natl. Acad. Sci. USA 2012, 109, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Herzig, Y.; Nevo, S.; Bornstein, C.; Brezis, M.R.; Ben-Hur, S.; Shkedy, A.; Eisenberg-Bord, M.; Levi, B.; Delacher, M.; Goldfarb, Y.; et al. Transcriptional programs that control expression of the autoimmune regulator gene Aire. Nat. Immunol. 2017, 18, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Flies, D.B. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Igarashi, K. Polyamine regulation of the synthesis of thymidine kinase in bovine lymphocytes. Arch. Biochem. Biophys. 1990, 278, 277–283. [Google Scholar] [CrossRef]
- Wang, R.; Dillon, C.P.; Shi, L.Z.; Milasta, S.; Carter, R.; Finkelstein, D.; McCormick, L.L.; Fitzgerald, P.; Chi, H.; Munger, J.; et al. The transcription factor Myc controls metabolic reprogramming upon T lymphocyte activation. Immunity 2011, 35, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, J.; Gravestein, L.A.; Tesselaar, K.; van Lier, R.A.; Schumacher, T.N.; Borst, J. CD27 is required for generation and long-term maintenance of T cell immunity. Nat. Immunol. 2000, 1, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Agematsu, K.; Hokibara, S.; Nagumo, H.; Komiyama, A. CD27: A memory B-cell marker. Immunol. Today 2000, 21, 204–206. [Google Scholar] [CrossRef]
- Takeda, K.; Oshima, H.; Hayakawa, Y.; Akiba, H.; Atsuta, M.; Kobata, T.; Kobayashi, K.; Ito, M.; Yagita, H.; Okumura, K. CD27-mediated activation of murine NK cells. J. Immunol. 2000, 164, 1741–1745. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Stanley, P.; Jones, K.; Svensson, L.; McDowall, A.; Hogg, N. The role of the integrin LFA-1 in T-lymphocyte migration. Immunol. Rev. 2007, 218, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Nocentini, G.; Giunchi, L.; Ronchetti, S.; Krausz, L.T.; Bartoli, A.; Moraca, R.; Migliorati, G.; Riccardi, C. A new member of the tumor necrosis factor/nerve growth factor receptor family inhibits T cell receptor-induced apoptosis. Proc. Natl. Acad. Sci. USA 1997, 94, 6216–6221. [Google Scholar] [CrossRef] [PubMed]
- Khayyamian, S.; Hutloff, A.; Buchner, K.; Grafe, M.; Henn, V.; Kroczek, R.A.; Mages, H.W. ICOS-ligand, expressed on human endothelial cells, costimulates Th1 and Th2 cytokine secretion by memory CD4+ T cells. Proc. Natl. Acad. Sci. USA 2002, 99, 6198–6203. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Freeman, G.J. The CD160, BTLA, LIGHT/HVEM pathway: A bidirectional switch regulating T-cell activation. Immunol. Rev. 2009, 229, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.T.; Dudl, E.; LeRoy, E.; Murray, R.; Sprent, J.; Weinberg, K.I.; Surh, C.D. IL-7 is critical for homeostatic proliferation and survival of naive T cells. Proc. Natl. Acad. Sci. USA 2001, 98, 8732–8737. [Google Scholar] [CrossRef] [PubMed]
- Wofford, J.A.; Wieman, H.L.; Jacobs, S.R.; Zhao, Y.; Rathmell, J.C. IL-7 promotes Glut1 trafficking and glucose uptake via STAT5-mediated activation of Akt to support T-cell survival. Blood 2008, 111, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Pallard, C.; Stegmann, A.P.; van Kleffens, T.; Smart, F.; Venkitaraman, A.; Spits, H. Distinct roles of the phosphatidylinositol 3-kinase and STAT5 pathways in IL-7-mediated development of human thymocyte precursors. Immunity 1999, 10, 525–535. [Google Scholar] [CrossRef]
- Jacobs, S.R.; Michalek, R.D.; Rathmell, J.C. IL-7 is essential for homeostatic control of T cell metabolism in vivo. J. Immunol. 2010, 184, 3461–3469. [Google Scholar] [CrossRef] [PubMed]
- MacIver, N.J.; Michalek, R.D.; Rathmell, J.C. Metabolic regulation of T lymphocytes. Annu. Rev. Immunol. 2013, 31, 259–283. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Li, Y.; Zhu, T.; Wu, J.; Guan, K.L. TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nat. Cell Biol. 2002, 4, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Li, Y.; Xu, T.; Guan, K.L. Rheb GTPase is a direct target of TSC2 GAP activity and regulates mTOR signaling. Genes Dev. 2003, 17, 1829–1834. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Zhu, T.; Guan, K.L. TSC2 mediates cellular energy response to control cell growth and survival. Cell 2003, 115, 577–590. [Google Scholar] [CrossRef]
- Iwata, T.N.; Ramirez, J.A.; Tsang, M.; Park, H.; Margineantu, D.H.; Hockenbery, D.M.; Iritani, B.M. Conditional Disruption of Raptor Reveals an Essential Role for mTORC1 in B Cell Development, Survival, and Metabolism. J. Immunol. 2016, 197, 2250–2260. [Google Scholar] [CrossRef] [PubMed]
- Delgoffe, G.M.; Kole, T.P.; Zheng, Y.; Zarek, P.E.; Matthews, K.L.; Xiao, B.; Worley, P.F.; Kozma, S.C.; Powell, J.D. The mTOR kinase differentially regulates effector and regulatory T cell lineage commitment. Immunity 2009, 30, 832–844. [Google Scholar] [CrossRef] [PubMed]
- Guertin, D.A.; Stevens, D.M.; Thoreen, C.C.; Burds, A.A.; Kalaany, N.Y.; Moffat, J.; Brown, M.; Fitzgerald, K.J.; Sabatini, D.M. Ablation in mice of the mTORC components raptor, rictor, or mLST8 reveals that mTORC2 is required for signaling to Akt-FOXO and PKCalpha, but not S6K1. Dev. Cell 2006, 11, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.J.; Kosmatka, M.; Bardeesy, N.; Hurley, R.L.; Witters, L.A.; DePinho, R.A.; Cantley, L.C. The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress. Proc. Natl. Acad. Sci. USA 2004, 101, 3329–3335. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.; Johnstone, S.R.; Dickerson, K.; Leiper, F.C.; Fryer, L.G.; Neumann, D.; Schlattner, U.; Wallimann, T.; Carlson, M.; Carling, D. LKB1 is the upstream kinase in the AMP-activated protein kinase cascade. Curr. Biol. 2003, 13, 2004–2008. [Google Scholar] [CrossRef] [PubMed]
- Van der Windt, G.J.; Pearce, E.L. Metabolic switching and fuel choice during T-cell differentiation and memory development. Immunol. Rev. 2012, 249, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Frauwirth, K.A.; Riley, J.L.; Harris, M.H.; Parry, R.V.; Rathmell, J.C.; Plas, D.R.; Elstrom, R.L.; June, C.H.; Thompson, C.B. The CD28 signaling pathway regulates glucose metabolism. Immunity 2002, 16, 769–777. [Google Scholar] [CrossRef]
- Boomer, J.S.; Green, J.M. An enigmatic tail of CD28 signaling. Cold Spring Harb. Perspect. Biol. 2010, 2, a002436. [Google Scholar] [CrossRef] [PubMed]
- Park, S.G.; Schulze-Luehrman, J.; Hayden, M.S.; Hashimoto, N.; Ogawa, W.; Kasuga, M.; Ghosh, S. The kinase PDK1 integrates T cell antigen receptor and CD28 coreceptor signaling to induce NF-κB and activate T cells. Nat. Immunol. 2009, 10, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Balagopalan, L.; Barr, V.A.; Sommers, C.L.; Barda-Saad, M.; Goyal, A.; Isakowitz, M.S.; Samelson, L.E. c-Cbl-mediated regulation of LAT-nucleated signaling complexes. Mol. Cell. Biol. 2007, 27, 8622–8636. [Google Scholar] [CrossRef] [PubMed]
- Verbist, K.C.; Guy, C.S.; Milasta, S.; Liedmann, S.; Kaminski, M.M.; Wang, R.; Green, D.R. Metabolic maintenance of cell asymmetry following division in activated T lymphocytes. Nature 2016, 532, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.C.; Bihuniak, J.D.; Macintyre, A.N.; Staron, M.; Liu, X.; Amezquita, R.; Tsui, Y.C.; Cui, G.; Micevic, G.; Perales, J.C.; et al. Phosphoenolpyruvate Is a Metabolic Checkpoint of Anti-tumor T Cell Responses. Cell 2015, 162, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Wakasawa, T.; Shima, Y.; Tsuboi, I.; Aizawa, S.; Tamai, I. Role of polyamines derived from arginine in differentiation and proliferation of human blood cells. Biol. Pharm. Bull. 2006, 29, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y.; Maeda, T.; Aizawa, S.; Tsuboi, I.; Kobayashi, D.; Kato, R.; Tamai, I. l-arginine import via cationic amino acid transporter CAT1 is essential for both differentiation and proliferation of erythrocytes. Blood 2006, 107, 1352–1356. [Google Scholar] [CrossRef] [PubMed]
- Bachrach, U.; Persky, S. Interaction of oxidized polyamines with DNA. V. Inhibition of nucleic acid synthesis. Biochim. Biophys. Acta 1969, 179, 484–493. [Google Scholar] [CrossRef]
- Francke, B. Cell-free synthesis of herpes simplex virus DNA: The influence of polyamines. Biochemistry 1978, 17, 5494–5499. [Google Scholar] [CrossRef] [PubMed]
- Leveque, J.; Burtin, F.; Catros-Quemener, V.; Havouis, R.; Moulinoux, J.P. The gastrointestinal polyamine source depletion enhances DFMO induced polyamine depletion in MCF-7 human breast cancer cells in vivo. Anticancer Res. 1998, 18, 2663–2668. [Google Scholar] [PubMed]
- Hessels, J.; Kingma, A.W.; Ferwerda, H.; Keij, J.; van den Berg, G.A.; Muskiet, F.A. Microbial flora in the gastrointestinal tract abolishes cytostatic effects of α-difluoromethylornithine in vivo. Int. J. Cancer 1989, 43, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, S.; Vassylyev, D.G.; Matsushima, M.; Kashiwagi, K.; Igarashi, K.; Morikawa, K. Crystal structure of PotD, the primary receptor of the polyamine transport system in Escherichia coli. J. Biol. Chem. 1996, 271, 9519–9525. [Google Scholar] [CrossRef] [PubMed]
- Tomitori, H.; Kashiwagi, K.; Asakawa, T.; Kakinuma, Y.; Michael, A.J.; Igarashi, K. Multiple polyamine transport systems on the vacuolar membrane in yeast. Biochem. J. 2001, 353, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Satriano, J.; Isome, M.; Casero, R.A., Jr.; Thomson, S.C.; Blantz, R.C. Polyamine transport system mediates agmatine transport in mammalian cells. Am. J. Physiol. Cell Physiol. 2001, 281, C329–C334. [Google Scholar] [CrossRef] [PubMed]
- Sakata, K.; Kashiwagi, K.; Igarashi, K. Properties of a polyamine transporter regulated by antizyme. Biochem. J. 2000, 347, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Uemura, T.; Stringer, D.E.; Blohm-Mangone, K.A.; Gerner, E.W. Polyamine transport is mediated by both endocytic and solute carrier transport mechanisms in the gastrointestinal tract. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G517–G522. [Google Scholar] [CrossRef] [PubMed]
- Wolfer, A.; Bakker, T.; Wilson, A.; Nicolas, M.; Ioannidis, V.; Littman, D.R.; Lee, P.P.; Wilson, C.B.; Held, W.; MacDonald, H.R.; et al. Inactivation of Notch 1 in immature thymocytes does not perturb CD4 or CD8T cell development. Nat. Immunol. 2001, 2, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Wildin, R.S.; Garvin, A.M.; Pawar, S.; Lewis, D.B.; Abraham, K.M.; Forbush, K.A.; Ziegler, S.F.; Allen, J.M.; Perlmutter, R.M. Developmental regulation of lck gene expression in T lymphocytes. J. Exp. Med. 1991, 173, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Dose, M.; Khan, I.; Guo, Z.; Kovalovsky, D.; Krueger, A.; von Boehmer, H.; Khazaie, K.; Gounari, F. c-Myc mediates pre-TCR-induced proliferation but not developmental progression. Blood 2006, 108, 2669–2677. [Google Scholar] [CrossRef] [PubMed]
- Dezfouli, S.; Bakke, A.; Huang, J.; Wynshaw-Boris, A.; Hurlin, P.J. Inflammatory disease and lymphomagenesis caused by deletion of the Myc antagonist Mnt in T cells. Mol. Cell. Biol. 2006, 26, 2080–2092. [Google Scholar] [CrossRef] [PubMed]
- Jain, J.; Nalefski, E.A.; McCaffrey, P.G.; Johnson, R.S.; Spiegelman, B.M.; Papaioannou, V.; Rao, A. Normal peripheral T-cell function in c-Fos-deficient mice. Mol. Cell. Biol. 1994, 14, 1566–1574. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wrighton, C.; Busslinger, M. Direct transcriptional stimulation of the ornithine decarboxylase gene by Fos in PC12 cells but not in fibroblasts. Mol. Cell. Biol. 1993, 13, 4657–4669. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Choi, B.S.; Martinez-Falero, I.C.; Corset, C.; Munder, M.; Modolell, M.; Muller, I.; Kropf, P. Differential impact of l-arginine deprivation on the activation and effector functions of T cells and macrophages. J. Leukoc. Biol. 2009, 85, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Carr, E.L.; Kelman, A.; Wu, G.S.; Gopaul, R.; Senkevitch, E.; Aghvanyan, A.; Turay, A.M.; Frauwirth, K.A. Glutamine uptake and metabolism are coordinately regulated by ERK/MAPK during T lymphocyte activation. J. Immunol. 2010, 185, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.H.; Fragonas, J.C. Effects of anti-oxidants on ornithine decarboxylase in mitogenically-activated T lymphocytes. Biochim. Biophys. Acta 1992, 1133, 261–267. [Google Scholar] [CrossRef]
- Widjaja, C.E.; Olvera, J.G.; Metz, P.J.; Phan, A.T.; Savas, J.N.; de Bruin, G.; Leestemaker, Y.; Berkers, C.R.; de Jong, A.; Florea, B.I.; et al. Proteasome activity regulates CD8+ T lymphocyte metabolism and fate specification. J. Clin. Investig. 2017, 127, 3609–3623. [Google Scholar] [CrossRef] [PubMed]
- Geiger, R.; Rieckmann, J.C.; Wolf, T.; Basso, C.; Feng, Y.; Fuhrer, T.; Kogadeeva, M.; Picotti, P.; Meissner, F.; Mann, M.; et al. l-Arginine Modulates T Cell Metab.olism and Enhances Survival and Anti-tumor Activity. Cell 2016, 167, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Buck, M.D.; O‘Sullivan, D.; Klein Geltink, R.I.; Curtis, J.D.; Chang, C.H.; Sanin, D.E.; Qiu, J.; Kretz, O.; Braas, D.; van der Windt, G.J.; et al. Mitochondrial Dynamics Controls T Cell Fate through Metabolic Programming. Cell 2016, 166, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Klein Geltink, R.I.; O‘Sullivan, D.; Corrado, M.; Bremser, A.; Buck, M.D.; Buescher, J.M.; Firat, E.; Zhu, X.; Niedermann, G.; Caputa, G.; et al. Mitochondrial Priming by CD28. Cell 2017, 171, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Pearce, E.L.; Poffenberger, M.C.; Chang, C.H.; Jones, R.G. Fueling immunity: Insights into metabolism and lymphocyte function. Science 2013, 342, 1242454. [Google Scholar] [CrossRef] [PubMed]
- Alexander, E.T.; Minton, A.; Peters, M.C.; Phanstiel, O.t.; Gilmour, S.K. A novel polyamine blockade therapy activates an anti-tumor immune response. Oncotarget 2017, 8, 84140–84152. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Mandal, A.; Johansson, H.E.; Orjalo, A.V.; Park, M.H. Depletion of cellular polyamines, spermidine and spermine, causes a total arrest in translation and growth in mammalian cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2169–2174. [Google Scholar] [CrossRef] [PubMed]
- Gnanaprakasam, J.N.; Wang, R. MYC in Regulating Immunity: Metabolism and Beyond. Genes (Basel) 2017, 8, 88. [Google Scholar] [CrossRef] [PubMed]
- Ehrke, M.J.; Porter, C.W.; Eppolito, C.; Mihich, E. Selective modulation by alpha-difluoromethylornithine of T-lymphocyte and antibody-mediated cytotoxic responses to mouse tumor allografts. Cancer Res. 1986, 46, 2798–2803. [Google Scholar] [PubMed]
- Bowlin, T.L.; McKown, B.J.; Sunkara, P.S. Increased ornithine decarboxylase activity and polyamine biosynthesis are required for optimal cytolytic T lymphocyte induction. Cell. Immunol. 1987, 105, 110–117. [Google Scholar] [CrossRef]
- Bowlin, T.L.; McKown, B.J.; Schroeder, K.K. Methyl-acetylenicputrescine (MAP), an inhibitor of polyamine biosynthesis, reduces the frequency and cytolytic activity of alloantigen-induced LyT 2.2 positive lymphocytes in vivo. Int. J. Immunopharmacol. 1989, 11, 259–265. [Google Scholar] [CrossRef]
- Bowlin, T.L.; Rosenberger, A.L.; McKown, B.J. α-difluoromethylornithine, an inhibitor of polyamine biosynthesis, augments cyclosporin A inhibition of cytolytic T lymphocyte induction. Clin. Exp. Immunol. 1989, 77, 151–156. [Google Scholar] [PubMed]
- Schall, R.P.; Sekar, J.; Tandon, P.M.; Susskind, B.M. Difluoromethylornithine (DFMO) arrests murine CTL development in the late, pre-effector stage. Immunopharmacology 1991, 21, 129–143. [Google Scholar] [CrossRef]
- Bowlin, T.L.; Davis, G.F.; McKown, B.J. Inhibition of alloantigen-induced cytolytic T lymphocytes in vitro with (2R,5R)-6-heptyne-2,5-diamine, an irreversible inhibitor of ornithine decarboxylase. Cell. Immunol. 1988, 111, 443–450. [Google Scholar] [CrossRef]
- Lampropoulou, V.; Sergushichev, A.; Bambouskova, M.; Nair, S.; Vincent, E.E.; Loginicheva, E.; Cervantes-Barragan, L.; Ma, X.; Huang, S.C.; Griss, T.; et al. Itaconate Links Inhibition of Succinate Dehydrogenase with Macrophage Metabolic Remodeling and Regulation of Inflammation. Cell Metab. 2016, 24, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Rathmell, J.; Pearce, E. SnapShot: Immunometabolism. Cell Metab. 2015, 22, 190.e1. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.; O‘Neill, L.A. Metabolic reprogramming in macrophages and dendritic cells in innate immunity. Cell Res. 2015, 25, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Youn, J.I.; Gabrilovich, D.I. Reciprocal relationship between myeloid-derived suppressor cells and T cells. J. Immunol. 2013, 191, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.I.; Collazo, M.; Shalova, I.N.; Biswas, S.K.; Gabrilovich, D.I. Characterization of the nature of granulocytic myeloid-derived suppressor cells in tumor-bearing mice. J. Leukoc. Biol. 2012, 91, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.C.; Ochoa, A.C.; Al-Khami, A.A. Arginine Metabolism in Myeloid Cells Shapes Innate and Adaptive Immunity. Front. Immunol. 2017, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.Y.; Iyo, A.; Piletz, J.E.; Regunathan, S. Expression of human arginine decarboxylase, the biosynthetic enzyme for agmatine. Biochim. Biophys. Acta 2004, 1670, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, J.; Thompson, P.R. Protein Arginine Methylation and Citrullination in Epigenetic Regulation. ACS Chem. Biol. 2016, 11, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Zanovello, P. Regulation of immune responses by l-arginine metabolism. Nat. Rev. Immunol. 2005, 5, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Geng, Z.; Dominguez, D.; Chen, S.; Fan, J.; Qin, L.; Long, A.; Zhang, Y.; Kuzel, T.M.; Zhang, B. Targeting Ornithine Decarboxylase by α-Difluoromethylornithine Inhibits Tumor Growth by Impairing Myeloid-Derived Suppressor Cells. J. Immunol. 2016, 196, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Ziv, Y.; Fazio, V.W.; Kitago, K.; Gupta, M.K.; Sawady, J.; Nishioka, K. Effect of tamoxifen on 1,2-dimethylhydrazine-HCl-induced colon carcinogenesis in rats. Anticancer Res. 1997, 17, 803–810. [Google Scholar] [PubMed]
- Bowlin, T.L.; Hoeper, B.J.; Rosenberger, A.L.; Davis, G.F.; Sunkara, P.S. Effects of three irreversible inhibitors of ornithine decarboxylase on macrophage-mediated tumoricidal activity and antitumor activity in B16F1 tumor-bearing mice. Cancer Res. 1990, 50, 4510–4514. [Google Scholar] [PubMed]
- Hayes, C.S.; Shicora, A.C.; Keough, M.P.; Snook, A.E.; Burns, M.R.; Gilmour, S.K. Polyamine-blocking therapy reverses immunosuppression in the tumor microenvironment. Cancer Immunol. Res. 2014, 2, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Schrum, A.G.; Cho, H.I.; Celis, E.; Gabrilovich, D.I. Mechanism of T cell tolerance induced by myeloid-derived suppressor cells. J. Immunol. 2010, 184, 3106–3116. [Google Scholar] [CrossRef] [PubMed]
- Sunderkotter, C.; Nikolic, T.; Dillon, M.J.; Van Rooijen, N.; Stehling, M.; Drevets, D.A.; Leenen, P.J. Subpopulations of mouse blood monocytes differ in maturation stage and inflammatory response. J. Immunol. 2004, 172, 4410–4417. [Google Scholar] [CrossRef] [PubMed]
- Voisin, M.B.; Buzoni-Gatel, D.; Bout, D.; Velge-Roussel, F. Both expansion of regulatory GR1+ CD11b+ myeloid cells and anergy of T lymphocytes participate in hyporesponsiveness of the lung-associated immune system during acute toxoplasmosis. Infect. Immun. 2004, 72, 5487–5492. [Google Scholar] [CrossRef] [PubMed]
- Mencacci, A.; Montagnoli, C.; Bacci, A.; Cenci, E.; Pitzurra, L.; Spreca, A.; Kopf, M.; Sharpe, A.H.; Romani, L. CD80+Gr-1+ myeloid cells inhibit development of antifungal Th1 immunity in mice with candidiasis. J. Immunol. 2002, 169, 3180–3190. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Spector, S.A. HIV type 1 gp120-induced expansion of myeloid derived suppressor cells is dependent on interleukin 6 and suppresses immunity. J. Infect. Dis. 2014, 209, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Patel, S.; Tcyganov, E.; Gabrilovich, D.I. The Nature of Myeloid-Derived Suppressor Cells in the Tumor Microenvironment. Trends Immunol. 2016, 37, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Niino, D.; Komohara, Y.; Murayama, T.; Aoki, R.; Kimura, Y.; Hashikawa, K.; Kiyasu, J.; Takeuchi, M.; Suefuji, N.; Sugita, Y.; et al. Ratio of M2 macrophage expression is closely associated with poor prognosis for Angioimmunoblastic T-cell lymphoma (AITL). Pathol. Int. 2010, 60, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; He, F.; Feng, F.; Liu, X.W.; Dong, G.Y.; Qin, H.Y.; Hu, X.B.; Zheng, M.H.; Liang, L.; Feng, L.; et al. Notch signaling determines the M1 versus M2 polarization of macrophages in antitumor immune responses. Cancer Res. 2010, 70, 4840–4849. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Nelson, A.; Youn, J.I.; Cheng, P.; Quiceno, D.; Gabrilovich, D.I. Antigen-specific CD4(+) T cells regulate function of myeloid-derived suppressor cells in cancer via retrograde MHC class II signaling. Cancer Res. 2012, 72, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.L.; Kelly, B.; Logan, A.; Costa, A.S.H.; Varma, M.; Bryant, C.E.; Tourlomousis, P.; Dabritz, J.H.M.; Gottlieb, E.; Latorre, I.; et al. Succinate Dehydrogenase Supports Metabolic Repurposing of Mitochondria to Drive Inflammatory Macrophages. Cell 2016, 167, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, Q.; Du, Y.; Liu, F.; Zhang, Y.; Feng, Y.; Jin, F. l-arginine and docetaxel synergistically enhance anti-tumor immunity by modifying the immune status of tumor-bearing mice. Int. Immunopharmacol. 2016, 35, 7–14. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Lin, H.; Yuan, L.; Li, B. Combination therapy with l-arginine and α-PD-L1 antibody boosts immune response against osteosarcoma in immunocompetent mice. Cancer Biol. Ther. 2017, 18, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Mondanelli, G.; Bianchi, R.; Pallotta, M.T.; Orabona, C.; Albini, E.; Iacono, A.; Belladonna, M.L.; Vacca, C.; Fallarino, F.; Macchiarulo, A.; et al. A Relay Pathway between Arginine and Tryptophan Metabolism Confers Immunosuppressive Properties on Dendritic Cells. Immunity 2017, 46, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Rubin, R.L.; Burlingame, R.W. Drug-induced autoimmunity: A disorder at the interface between metabolism and immunity. Biochem. Soc. Trans. 1991, 19, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Rathmell, J.C. Apoptosis and B cell tolerance. Curr. Dir. Autoimmun. 2003, 6, 38–60. [Google Scholar] [PubMed]
- Teti, D.; Visalli, M.; McNair, H. Analysis of polyamines as markers of (patho)physiological conditions. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2002, 781, 107–149. [Google Scholar] [CrossRef]
- Karouzakis, E.; Gay, R.E.; Gay, S.; Neidhart, M. Increased recycling of polyamines is associated with global DNA hypomethylation in rheumatoid arthritis synovial fibroblasts. Arthritis Rheumatol. 2012, 64, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Pignata, S.; Di Luccia, A.; Lamanda, R.; Menchise, A.; D‘Agostino, L. Interaction of putrescine with nuclear oligopeptides in the enterocyte-like Caco-2 cells. Digestion 1999, 60, 255–261. [Google Scholar] [CrossRef] [PubMed]
- D‘Agostino, L.; Di Luccia, A. Polyamines interact with DNA as molecular aggregates. Eur. J. Biochem. 2002, 269, 4317–4325. [Google Scholar] [CrossRef] [PubMed]
- D‘Agostino, L.; di Pietro, M.; Di Luccia, A. Nuclear aggregates of polyamines are supramolecular structures that play a crucial role in genomic DNA protection and conformation. FEBS J. 2005, 272, 3777–3787. [Google Scholar] [CrossRef] [PubMed]
- Riboldi, P.; Gerosa, M.; Moroni, G.; Radice, A.; Allegri, F.; Sinico, A.; Tincani, A.; Meroni, P.L. Anti-DNA antibodies: A diagnostic and prognostic tool for systemic lupus erythematosus? Autoimmunity 2005, 38, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Fineschi, S.; Borghi, M.O.; Riboldi, P.; Gariglio, M.; Buzio, C.; Landolfo, S.; Cebecauer, L.; Tuchynova, A.; Rovensky, J.; Meroni, P.L. Prevalence of autoantibodies against structure specific recognition protein 1 in systemic lupus erythematosus. Lupus 2004, 13, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Casero, R.A., Jr. Say what? The activity of the polyamine biosynthesis inhibitor difluoromethylornithine in chemoprevention is a result of reduced thymidine pools? Cancer Discov. 2013, 3, 975–977. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ruan, H.; Hill, J.R.; Fatemie-Nainie, S.; Morris, D.R. Cell-specific translational regulation of S-adenosylmethionine decarboxylase mRNA. Influence of the structure of the 5′ transcript leader on regulation by the upstream open reading frame. J. Biol. Chem. 1994, 269, 17905–17910. [Google Scholar] [PubMed]
- Ruan, H.; Shantz, L.M.; Pegg, A.E.; Morris, D.R. The upstream open reading frame of the mRNA encoding S-adenosylmethionine decarboxylase is a polyamine-responsive translational control element. J. Biol. Chem. 1996, 271, 29576–29582. [Google Scholar] [CrossRef] [PubMed]
- Bale, S.; Lopez, M.M.; Makhatadze, G.I.; Fang, Q.; Pegg, A.E.; Ealick, S.E. Structural basis for putrescine activation of human S-adenosylmethionine decarboxylase. Biochemistry 2008, 47, 13404–13417. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hesterberg, R.S.; Cleveland, J.L.; Epling-Burnette, P.K. Role of Polyamines in Immune Cell Functions. Med. Sci. 2018, 6, 22. https://doi.org/10.3390/medsci6010022
Hesterberg RS, Cleveland JL, Epling-Burnette PK. Role of Polyamines in Immune Cell Functions. Medical Sciences. 2018; 6(1):22. https://doi.org/10.3390/medsci6010022
Chicago/Turabian StyleHesterberg, Rebecca S., John L. Cleveland, and Pearlie K. Epling-Burnette. 2018. "Role of Polyamines in Immune Cell Functions" Medical Sciences 6, no. 1: 22. https://doi.org/10.3390/medsci6010022
APA StyleHesterberg, R. S., Cleveland, J. L., & Epling-Burnette, P. K. (2018). Role of Polyamines in Immune Cell Functions. Medical Sciences, 6(1), 22. https://doi.org/10.3390/medsci6010022