
An Overview of Malaria Transmission Mechanisms, Control, and Modeling


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Inclusion and Exclusion Criteria for Paper Eligibility
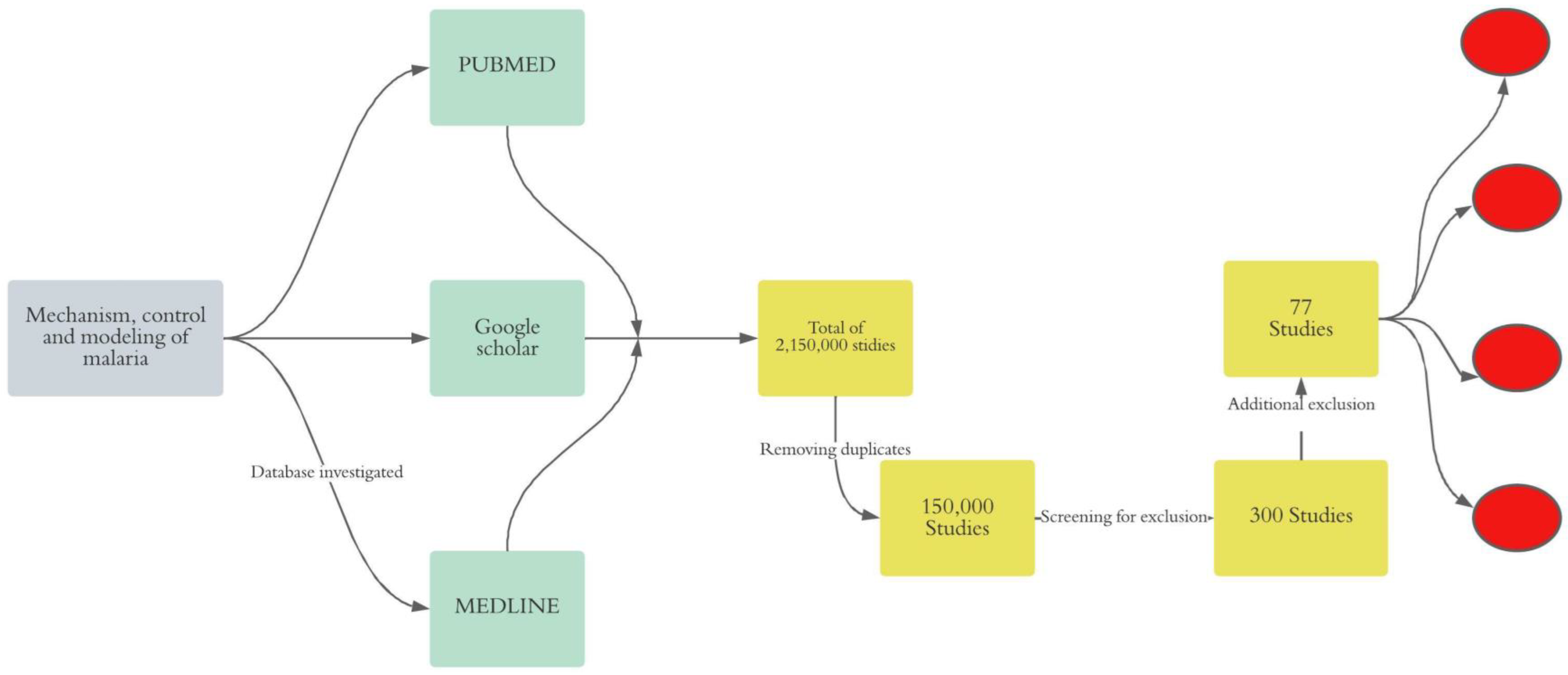
2.2. Search Strategy
2.3. Screening of Studies and Evidence Extraction
2.4. Analysis and Gaps in the Analysis
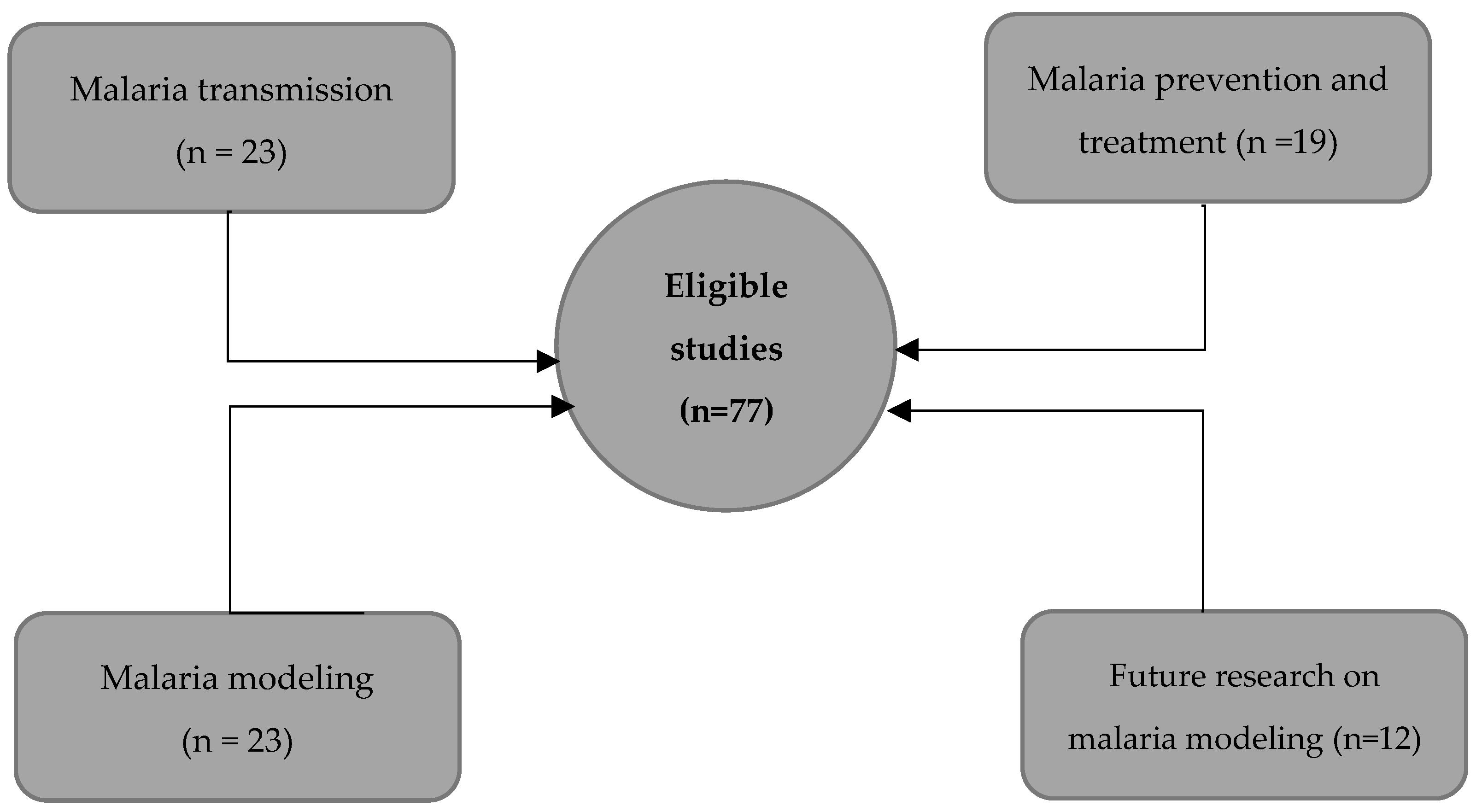
3. Results
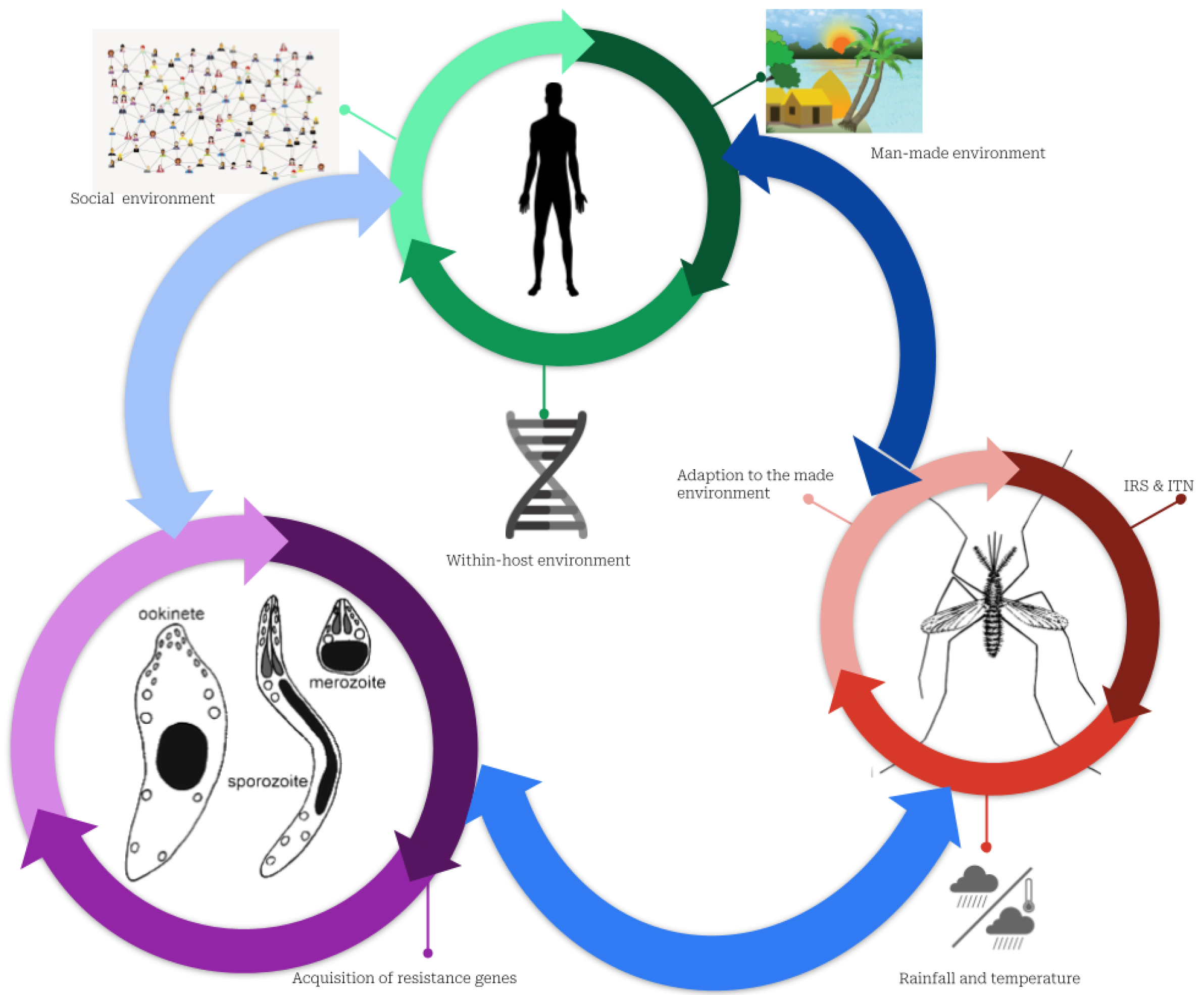
3.1. Malaria Transmission Mechanism
3.2. Malaria Prevention and Treatment
3.3. Mathematical Modeling of Malaria
3.4. Malaria Modeling: Present and Future
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACT | Artemisinin-based Combination Therapies |
| DDT | Dichlorodiphenyltrichloroethane |
| GMAP | Global Malaria Action Plan |
| GMEP | Global Malaria Eradication Program |
| IBM | Individual-based models |
| IRS | Indoor residual spraying |
| ITN | Insecticide-treated bed nets |
| R0 | reproductive number |
| SSA | Sub-Saharan Africa |
| SIR | Susceptible Infectious Removed model |
| WHO | World Health Organization |
References
- Ndugwa, R.P.; Ramroth, H.; Müller, O.; Jasseh, M.; Sié, A.; Kouyaté, B.; Greenwood, B.; Becher, H. Comparison of All-Cause and Malaria-Specific Mortality from Two West African Countries with Different Malaria Transmission Patterns. Malar. J. 2008, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Sachs, J.; Malaney, P. The Economic and Social Burden of Malaria. Nature 2002, 415, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Savi, M.K.; Callo-Concha, D.; Tonnang, H.E.Z.; Borgemeister, C. Emerging Properties of Malaria Transmission and Persistence in Urban Accra, Ghana: Evidence from a Participatory System Approach. Malar. J. 2021, 20, 321. [Google Scholar] [CrossRef] [PubMed]
- Asare, E.O.; Amekudzi, L.K. Assessing Climate Driven Malaria Variability in Ghana Using a Regional Scale Dynamical Model. Climate 2017, 5, 20. [Google Scholar] [CrossRef]
- Akazili, J.; Aikins, M.; Binka, F.N. Malaria Treatment in Northern Ghana: What Is the Treatment Cost per Case to Households? Afr. J. Health Sci. 2007, 14, 70–79. [Google Scholar] [CrossRef]
- Jowett, M.; Miller, N.J. The Financial Burden of Malaria in Tanzania: Implications for Future Government Policy. Int. J. Health Plann. Manag. 2005, 20, 67–84. [Google Scholar] [CrossRef]
- Wells, T.N.C.; Alonso, P.L.; Gutteridge, W.E. New Medicines to Improve Control and Contribute to the Eradication of Malaria. Nat. Rev. Drug. Discov. 2009, 8, 879–891. [Google Scholar] [CrossRef]
- Breman, J. Rolling Back Malaria: Action or Rhetoric? Available online: https://www.semanticscholar.org/author/J.-Breman/144538342 (accessed on 15 November 2022).
- Arifin, S.N.; Zhou, Y.; Davis, G.J.; Gentile, J.E.; Madey, G.R.; Collins, F.H. An Agent-Based Model of the Population Dynamics of Anopheles Gambiae. Malar. J. 2014, 13, 424. [Google Scholar] [CrossRef]
- Harrington, L.C.; Edman, J.D.; Scott, T.W. Why Do Female Aedes Aegypti (Diptera: Culicidae) Feed Preferentially and Frequently on Human Blood? J. Med. Entomol. 2001, 38, 411–422. [Google Scholar] [CrossRef]
- Li, Z.-X.; Pickett, J.A.; Field, L.M.; Zhou, J.-J. Identification and Expression of Odorant-Binding Proteins of the Malaria-Carrying Mosquitoes Anopheles Gambiae and Anopheles Arabiensis. Arch. Insect. Biochem. Physiol. 2005, 58, 175–189. [Google Scholar] [CrossRef]
- Tuno, N.; Kjaerandsen, J.; Badu, K.; Kruppa, T. Blood-Feeding Behavior of Anopheles Gambiae and Anopheles Melas in Ghana, Western Africa. J. Med. Entomol. 2010, 47, 28–31. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The Dominant Anopheles Vectors of Human Malaria in Africa, Europe and the Middle East: Occurrence Data, Distribution Maps and Bionomic Précis. Parasites Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [PubMed]
- Childs, L.M.; Abuelezam, N.N.; Dye, C.; Gupta, S.; Murray, M.B.; Williams, B.G.; Buckee, C.O. Modelling Challenges in Context: Lessons from Malaria, HIV, and Tuberculosis. Epidemics 2015, 10, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Eikenberry, S.E.; Gumel, A.B. Mathematical Modeling of Climate Change and Malaria Transmission Dynamics: A Historical Review. J. Math. Biol. 2018, 77, 857–933. [Google Scholar] [CrossRef]
- Adu-Prah, S.; Kofi Tetteh, E. Spatiotemporal Analysis of Climate Variability Impacts on Malaria Prevalence in Ghana. Appl. Geogr. 2015, 60, 266–273. [Google Scholar] [CrossRef]
- White, N.J.; Pukrittayakamee, S.; Hien, T.T.; Faiz, M.A.; Mokuolu, O.A.; Dondorp, A.M. Malaria. Lancet 2014, 383, 723–735. [Google Scholar] [CrossRef]
- Chaturvedi, H.K.; Mahanta, J.; Bajpai, R.C.; Pandey, A. Risk of Malaria among Febrile Patients: Retrospective Analysis of a Hospital-Based Study in an Endemic Area of Northeast India. Int. Health 2014, 6, 144–151. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Paaijmans, K.P.; Johnson, L.R.; Balzer, C.; Ben-Horin, T.; de Moor, E.; McNally, A.; Pawar, S.; Ryan, S.J.; Smith, T.C.; et al. Optimal Temperature for Malaria Transmission Is Dramatically Lower than Previously Predicted. Ecol. Lett. 2013, 16, 22–30. [Google Scholar] [CrossRef]
- Mwangangi, J.M.; Mbogo, C.M.; Orindi, B.O.; Muturi, E.J.; Midega, J.T.; Nzovu, J.; Gatakaa, H.; Githure, J.; Borgemeister, C.; Keating, J.; et al. Shifts in Malaria Vector Species Composition and Transmission Dynamics along the Kenyan Coast over the Past 20 Years. Malar. J. 2013, 12, 13. [Google Scholar] [CrossRef]
- Busula, A.O.; Takken, W.; de Boer, J.G.; Mukabana, W.R.; Verhulst, N.O. Variation in Host Preferences of Malaria Mosquitoes Is Mediated by Skin Bacterial Volatiles. Med. Vet. Entomol. 2017, 31, 320–326. [Google Scholar] [CrossRef]
- Verhulst, N.O.; Qiu, Y.T.; Beijleveld, H.; Maliepaard, C.; Knights, D.; Schulz, S.; Berg-Lyons, D.; Lauber, C.L.; Verduijn, W.; Haasnoot, G.W.; et al. Composition of Human Skin Microbiota Affects Attractiveness to Malaria Mosquitoes. PLoS ONE 2011, 6, e28991. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, R.; Mukabana, W.R.; Gouagna, L.C.; Koella, J.C. Malaria Infection Increases Attractiveness of Humans to Mosquitoes. PLoS Biol. 2005, 3, e298. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.; Schoone, G.; Schallig, H.; Verhage, D.; Telgt, D.; Eling, W.; Sauerwein, R. Quantification of Plasmodium Falciparum Gametocytes in Differential Stages of Development by Quantitative Nucleic Acid Sequence-Based Amplification. Mol. Biochem. Parasitol. 2004, 137, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Karnad, D.R.; Nor, M.B.M.; Richards, G.A.; Baker, T.; Amin, P. Council of the World Federation of Societies of Intensive and Critical Care Medicine Intensive Care in Severe Malaria: Report from the Task Force on Tropical Diseases by the World Federation of Societies of Intensive and Critical Care Medicine. J. Crit. Care 2018, 43, 356–360. [Google Scholar] [CrossRef]
- Kafy, H.T.; Ismail, B.A.; Mnzava, A.P.; Lines, J.; Abdin, M.S.E.; Eltaher, J.S.; Banaga, A.O.; West, P.; Bradley, J.; Cook, J.; et al. Impact of Insecticide Resistance in Anopheles Arabiensis on Malaria Incidence and Prevalence in Sudan and the Costs of Mitigation. Proc. Natl. Acad. Sci. USA 2017, 114, E11267–E11275. [Google Scholar] [CrossRef]
- Institute of Medicine (US) Committee for the Study on Malaria Prevention and Control. Malaria: Obstacles and Opportunities; Oaks, S.C., Mitchell, V.S., Pearson, G.W., Carpenter, C.C.J., Eds.; National Academies Press (US): Washington, DC, USA, 1991; ISBN 978-0-309-04527-8. [Google Scholar]
- Yam, X.Y.; Preiser, P.R. Host Immune Evasion Strategies of Malaria Blood Stage Parasite. Mol. BioSyst. 2017, 13, 2498–2508. [Google Scholar] [CrossRef]
- Talman, A.M.; Blagborough, A.M.; Sinden, R.E. A Plasmodium Falciparum Strain Expressing GFP throughout the Parasite’s Life-Cycle. PLoS ONE 2010, 5, e9156. [Google Scholar] [CrossRef]
- Bousema, T.; Drakeley, C. Determinants of Malaria Transmission at the Population Level. Cold. Spring. Harb. Perspect. Med. 2017, 7, a025510. [Google Scholar] [CrossRef]
- Sturrock, H.J.W.; Hsiang, M.S.; Cohen, J.M.; Smith, D.L.; Greenhouse, B.; Bousema, T.; Gosling, R.D. Targeting Asymptomatic Malaria Infections: Active Surveillance in Control and Elimination. PLoS Med. 2013, 10, e1001467. [Google Scholar] [CrossRef]
- Tusting, L.S.; Thwing, J.; Sinclair, D.; Fillinger, U.; Gimnig, J.; Bonner, K.E.; Bottomley, C.; Lindsay, S.W. Mosquito Larval Source Management for Controlling Malaria. Cochrane. Database. Syst. Rev. 2013, 8, CD008923. [Google Scholar] [CrossRef]
- Fillinger, U.; Lindsay, S.W. Larval Source Management for Malaria Control in Africa: Myths and Reality. Malar. J. 2011, 10, 353. [Google Scholar] [CrossRef] [PubMed]
- Liu, N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Roll Back Malaria Partnership. The Global Malaria Action Plan. In For a Free Malaria World; WHO: Geneva, Switzerland, 2008; p. 274. [Google Scholar]
- Aregawi, M.; Malm, K.L.; Wahjib, M.; Kofi, O.; Allotey, N.-K.; Yaw, P.N.; Abba-Baffoe, W.; Segbaya, S.; Owusu-Antwi, F.; Kharchi, A.T.; et al. Effect of Anti-Malarial Interventions on Trends of Malaria Cases, Hospital Admissions and Deaths, 2005–2015, Ghana. Malar. J. 2017, 16, 177. [Google Scholar] [CrossRef] [PubMed]
- Kudom, A.A.; Anane, L.N.; Afoakwah, R.; Adokoh, C.K. Relating High Insecticide Residues in Larval Breeding Habitats in Urban Residential Areas to the Selection of Pyrethroid Resistance in Anopheles Gambiae s.l. (Diptera: Culicidae) in Akim Oda, Ghana. J. Med. Entomol. 2018, 55, 490–495. [Google Scholar] [CrossRef]
- Moshi, I.R.; Ngowo, H.; Dillip, A.; Msellemu, D.; Madumla, E.P.; Okumu, F.O.; Coetzee, M.; Mnyone, L.L.; Manderson, L. Community Perceptions on Outdoor Malaria Transmission in Kilombero Valley, Southern Tanzania. Malar. J. 2017, 16, 274. [Google Scholar] [CrossRef]
- Eastman, R.T.; Fidock, D.A. Artemisinin-Based Combination Therapies: A Vital Tool in Efforts to Eliminate Malaria. Nat. Rev. Microbiol. 2009, 7, 864–874. [Google Scholar] [CrossRef]
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin Resistance in Plasmodium Falciparum Malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef]
- Yasuoka, J.; Jimba, M.; Levins, R. Application of Loop Analysis for Evaluation of Malaria Control Interventions. Malar. J. 2014, 13, 140. [Google Scholar] [CrossRef]
- Kaur, H.; Allan, E.L.; Mamadu, I.; Hall, Z.; Green, M.D.; Swamidos, I.; Dwivedi, P.; Culzoni, M.J.; Fernandez, F.M.; Garcia, G.; et al. Prevalence of Substandard and Falsified Artemisinin-Based Combination Antimalarial Medicines on Bioko Island, Equatorial Guinea. BMJ Glob. Health 2017, 2, e000409. [Google Scholar] [CrossRef]
- Briand, V.; Cottrell, G.; Massougbodji, A.; Cot, M. Intermittent Preventive Treatment for the Prevention of Malaria during Pregnancy in High Transmission Areas. Malar. J. 2007, 6, 160. [Google Scholar] [CrossRef]
- Dicko, A.; Roh, M.E.; Diawara, H.; Mahamar, A.; Soumare, H.M.; Lanke, K.; Bradley, J.; Sanogo, K.; Kone, D.T.; Diarra, K.; et al. Efficacy and Safety of Primaquine and Methylene Blue for Prevention of Plasmodium Falciparum Transmission in Mali: A Phase 2, Single-Blind, Randomised Controlled Trial. Lancet. Infect. Dis. 2018, 18, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Otieno, L.; Oneko, M.; Otieno, W.; Abuodha, J.; Owino, E.; Odero, C.; Mendoza, Y.; Andagalu, B.; Awino, N.; Ivinson, K.; et al. Safety and Immunogenicity of RTS,S/AS01 Malaria Vaccine in Infants and Children with WHO Stage 1 or 2 HIV Disease: A Randomised, Double-Blind, Controlled Trial. Lancet. Infect. Dis. 2016, 16, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, B.; Dicko, A.; Sagara, I.; Zongo, I.; Tinto, H.; Cairns, M.; Kuepfer, I.; Milligan, P.; Ouedraogo, J.-B.; Doumbo, O.; et al. Seasonal Vaccination against Malaria: A Potential Use for an Imperfect Malaria Vaccine. Malar. J. 2017, 16, 182. [Google Scholar] [CrossRef] [PubMed]
- Cheuka, P.M.; Mayoka, G.; Mutai, P.; Chibale, K. The Role of Natural Products in Drug Discovery and Development against Neglected Tropical Diseases. Molecules 2016, 22, 58. [Google Scholar] [CrossRef] [PubMed]
- Tarkang, P.A.; Appiah-Opong, R.; Ofori, M.F.; Ayong, L.S.; Nyarko, A.K. Application of Multi-Target Phytotherapeutic Concept in Malaria Drug Discovery: A Systems Biology Approach in Biomarker Identification. Biomark. Res. 2016, 4, 25. [Google Scholar] [CrossRef]
- Vlietinck, A.J.; Pieters, L.; Apers, S.; Cimanga, K.; Mesia, K.; Tona, L. The Value of Central-African Traditional Medicine for Lead Finding: Some Case Studies. J. Ethnopharmacol. 2015, 174, 607–617. [Google Scholar] [CrossRef]
- Ginsburg, H.; Deharo, E. A Call for Using Natural Compounds in the Development of New Antimalarial Treatments—An Introduction. Malar. J. 2011, 10, S1. [Google Scholar] [CrossRef]
- Ross, R. The Prevention of Malaria; E.P. Dutton & Company: New York, NY, USA, 1910. [Google Scholar]
- Ross, R. Some Quantitative Studies in Epidemiology. Nature 1911, 87, 466–467. [Google Scholar] [CrossRef]
- Macdonald, G. The Analysis of Equilibrium in Malaria. Trop. Dis. Bull. 1952, 49, 813–829. [Google Scholar]
- Nájera, J.A.; González-Silva, M.; Alonso, P.L. Some Lessons for the Future from the Global Malaria Eradication Programme (1955-1969). PLoS Med. 2011, 8, e1000412. [Google Scholar] [CrossRef]
- World Health Assembly. 22 Re-Examination of the Global Strategy of Malaria Eradication: Report of the Director-General; World Health Organization: Geneva, Switzerland, 1969.
- Smith, T.A.; Chitnis, N.; Penny, M.; Tanner, M. Malaria Modeling in the Era of Eradication. Cold. Spring. Harb. Perspect. Med. 2017, 7, a025460. [Google Scholar] [CrossRef]
- May, R.M. Stability and Complexity in Model Ecosystems; Princeton Landmarks in Biology; Princeton University Press: Princeton, NJ, USA, 2001; ISBN 978-0-691-08861-7. [Google Scholar]
- Colizza, V.; Barthélemy, M.; Barrat, A.; Vespignani, A. Epidemic Modeling in Complex Realities. CR Biol. 2007, 330, 364–374. [Google Scholar] [CrossRef]
- Kermack, W.O.; McKendrick, A.G. Contributions to the Mathematical Theory of Epidemics—I. Bltn. Mathcal. Biol. 1991, 53, 33–55. [Google Scholar] [CrossRef]
- Aikins, M.K.; Pickering, H.; Greenwood, B.M. Attitudes to Malaria, Traditional Practices and Bednets (Mosquito Nets) as Vector Control Measures: A Comparative Study in Five West African Countries. J. Trop. Med. Hyg. 1994, 97, 81–86. [Google Scholar]
- Smith, D.L.; Hay, S.I.; Noor, A.M.; Snow, R.W. Predicting Changing Malaria Risk after Expanded Insecticide-Treated Net Coverage in Africa. Trends Parasitol. 2009, 25, 511–516. [Google Scholar] [CrossRef]
- Smith, D.L.; Dushoff, J.; Snow, R.W.; Hay, S.I. The Entomological Inoculation Rate and Plasmodium Falciparum Infection in African Children. Nature 2005, 438, 492–495. [Google Scholar] [CrossRef]
- Smith, D.L.; McKenzie, F.E.; Snow, R.W.; Hay, S.I. Revisiting the Basic Reproductive Number for Malaria and Its Implications for Malaria Control. PLoS Biol. 2007, 5, e42. [Google Scholar] [CrossRef]
- Heesterbeek, H.; Anderson, R.M.; Andreasen, V.; Bansal, S.; de Angelis, D.; Dye, C.; Eames, K.T.D.; Edmunds, W.J.; Frost, S.D.W.; Funk, S.; et al. Modeling Infectious Disease Dynamics in the Complex Landscape of Global Health. Science 2015, 347, aaa4339. [Google Scholar] [CrossRef]
- Cohen, J.M.; Le Menach, A.; Pothin, E.; Eisele, T.P.; Gething, P.W.; Eckhoff, P.A.; Moonen, B.; Schapira, A.; Smith, D.L. Mapping Multiple Components of Malaria Risk for Improved Targeting of Elimination Interventions. Malar. J. 2017, 16, 459. [Google Scholar] [CrossRef]
- Agusto, F.B.; Teboh-Ewungkem, M.I.; Gumel, A.B. Mathematical Assessment of the Effect of Traditional Beliefs and Customs on the Transmission Dynamics of the 2014 Ebola Outbreaks. BMC Med. 2015, 13, 96. [Google Scholar] [CrossRef]
- Mushayabasa, S.; Bhunu, C.P.; Webb, C.; Dhlamini, M. A Mathematical Model for Assessing the Impact of Poverty on Yaws Eradication. Appl. Math. Model. 2012, 36, 1653–1667. [Google Scholar] [CrossRef]
- Ross, R. Mosquito Brigades and How to Organise Them/by Ronald Ross; Longmans, Green: London, UK, 1902. [Google Scholar]
- Hethcote, H.W. A Thousand and one epidemic models. In Proceedings of the Frontiers in Mathematical Biology; Levin, S.A., Ed.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 504–515. [Google Scholar]
- Xia, S.; Zhou, X.-N.; Liu, J. Systems Thinking in Combating Infectious Diseases. Infect. Dis. Poverty 2017, 6, 144. [Google Scholar] [CrossRef] [PubMed]
- Mecoli, M.; de Angelis, V.; Brailsford, S.C. Using System Dynamics to Evaluate Control Strategies for Mosquito-Borne Diseases Spread by Human Travel. Comput. Oper. Res. 2013, 40, 2219–2228. [Google Scholar] [CrossRef]
- Mandal, S.; Sarkar, R.R.; Sinha, S. Mathematical Models of Malaria—A Review. Malar. J. 2011, 10, 202. [Google Scholar] [CrossRef] [PubMed]
- Agusto, F.B.; del Valle, S.Y.; Blayneh, K.W.; Ngonghala, C.N.; Goncalves, M.J.; Li, N.; Zhao, R.; Gong, H. The Impact of Bed-Net Use on Malaria Prevalence. J. Biol. 2013, 320, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Ngonghala, C.N.; Del Valle, S.Y.; Zhao, R.; Mohammed-Awel, J. Quantifying the Impact of Decay in Bed-Net Efficacy on Malaria Transmission. J. Biol. 2014, 363, 247–261. [Google Scholar] [CrossRef]
- Le, P.V.V.; Kumar, P.; Ruiz, M.O. Stochastic Lattice-Based Modelling of Malaria Dynamics. Malar. J. 2018, 17, 250. [Google Scholar] [CrossRef] [PubMed]
- Dangerfield, C.E.; Ross, J.V.; Keeling, M.J. Integrating Stochasticity and Network Structure into an Epidemic Model. J. R Soc. Interface 2009, 6, 761–774. [Google Scholar] [CrossRef]
- Parham, P.E.; Michael, E. Outbreak Properties of Epidemic Models: The Roles of Temporal Forcing and Stochasticity on Pathogen Invasion Dynamics. J. Biol. 2011, 271, 1–9. [Google Scholar] [CrossRef]
- Smith, N.R.; Trauer, J.M.; Gambhir, M.; Richards, J.S.; Maude, R.J.; Keith, J.M.; Flegg, J.A. Agent-Based Models of Malaria Transmission: A Systematic Review. Malar. J. 2018, 17, 299. [Google Scholar] [CrossRef]
- Maude, R.J.; Pontavornpinyo, W.; Saralamba, S.; Aguas, R.; Yeung, S.; Dondorp, A.M.; Day, N.P.; White, N.J.; White, L.J. The Last Man Standing Is the Most Resistant: Eliminating Artemisinin-Resistant Malaria in Cambodia. Malar J. 2009, 8, 31. [Google Scholar] [CrossRef]
- Acevedo, M.A.; Prosper, O.; Lopiano, K.; Ruktanonchai, N.; Caughlin, T.T.; Martcheva, M.; Osenberg, C.W.; Smith, D.L. Spatial Heterogeneity, Host Movement and Mosquito-Borne Disease Transmission. PLoS ONE 2015, 10, e0127552. [Google Scholar] [CrossRef] [PubMed]
- Okell, L.C.; Paintain, L.S.; Webster, J.; Hanson, K.; Lines, J. From Intervention to Impact: Modelling the Potential Mortality Impact Achievable by Different Long-Lasting, Insecticide-Treated Net Delivery Strategies. Malar. J. 2012, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Johnston, G.L.; Smith, D.L.; Fidock, D.A. Malaria’s Missing Number: Calculating the Human Component of R0 by a Within-Host Mechanistic Model of Plasmodium Falciparum Infection and Transmission. PLoS Comput. Biol. 2013, 9, e1003025. [Google Scholar] [CrossRef] [PubMed]
- Frost, S.D.W.; Pybus, O.G.; Gog, J.R.; Viboud, C.; Bonhoeffer, S.; Bedford, T. Eight Challenges in Phylodynamic Inference. Epidemics 2015, 10, 88–92. [Google Scholar] [CrossRef]
- Buckee, C.O.; Wesolowski, A.; Eagle, N.; Hansen, E.; Snow, R.W. Mobile Phones and Malaria: Modeling Human and Parasite Travel. Travel. Med. Infect. Dis. 2013, 11, 15–22. [Google Scholar] [CrossRef]
- Ruktanonchai, N.W.; De Leenheer, P.; Tatem, A.J.; Alegana, V.A.; Caughlin, T.T.; Zu Erbach-Schoenberg, E.; Lourenço, C.; Ruktanonchai, C.W.; Smith, D.L. Identifying Malaria Transmission Foci for Elimination Using Human Mobility Data. PLoS Comput. Biol. 2016, 12, e1004846. [Google Scholar] [CrossRef]
- Ruktanonchai, N.W.; Bhavnani, D.; Sorichetta, A.; Bengtsson, L.; Carter, K.H.; Córdoba, R.C.; Le Menach, A.; Lu, X.; Wetter, E.; zu Erbach-Schoenberg, E.; et al. Census-Derived Migration Data as a Tool for Informing Malaria Elimination Policy. Malar. J. 2016, 15, 273. [Google Scholar] [CrossRef]
- Santos-Lozada, A.R.; Howard, J.T.; Verdery, A.M. How Differential Privacy Will Affect Our Understanding of Health Disparities in the United States. Proc. Natl. Acad. Sci. USA 2020, 117, 13405–13412. [Google Scholar] [CrossRef]
- Marino, A.; Bivona, D.A.; Bonacci, P. Updates in Central Nervous System Malaria: Literature Review and Considerations. Curr. Opin. Infect. Dis. 2022, 35, 255–261. [Google Scholar] [CrossRef]
- Siddiqui, A.J.; Bhardwaj, J.; Goyal, M.; Prakash, K.; Adnan, M.; Alreshidi, M.M.; Patel, M.; Soni, A.; Redman, W. Immune Responses in Liver and Spleen against Plasmodium Yoelii Pre-Erythrocytic Stages in Swiss Mice Model. J. Adv. Res. 2020, 24, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.K.; Kong, T.F.; Ng, C.S.; Chen, L.; Huang, Y.; Bhagat, A.A.S.; Nguyen, N.-T.; Preiser, P.R.; Han, J. Micromagnetic Resonance Relaxometry for Rapid Label-Free Malaria Diagnosis. Nat. Med. 2014, 20, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.K.; Ng, T.-T.; Loh, T.P. Machine Learning Assistive Rapid, Label-Free Molecular Phenotyping of Blood with Two-Dimensional NMR Correlational Spectroscopy. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, E.; Ferrauto, G.; Schwarzer, E.; Gianolio, E.; Valente, E.; Ulliers, D.; Aime, S.; Skorokhod, O. Relaxometric Studies of Erythrocyte Suspensions Infected by Plasmodium Falciparum: A Tool for Staging Infection and Testing Anti-Malarial Drugs. Magn. Reson. Med. 2020, 84, 3366–3378. [Google Scholar] [CrossRef]
- Arndt, L.; Koleala, T.; Orbán, Á.; Ibam, C.; Lufele, E.; Timinao, L.; Lorry, L.; Butykai, Á.; Kaman, P.; Molnár, A.P.; et al. Magneto-Optical Diagnosis of Symptomatic Malaria in Papua New Guinea. Nat. Commun. 2021, 12, 969. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Hashimoto, M.; Nagatomi, K.; Nogami, T.; Sofue, Y.; Hayashi, T.; Ido, Y.; Yatsushiro, S.; Abe, K.; Kajimoto, K.; et al. Field Evaluation of a Quantitative, and Rapid Malaria Diagnostic System Using a Fluorescent Blue-Ray Optical Device. BioRxiv 2019. [Google Scholar] [CrossRef]
- Chen, N.; Li, A.; Talluri, K. Reviews and Self-Selection Bias with Operational Implications. Manag. Sci. 2021, 67, 7472–7492. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savi, M.K. An Overview of Malaria Transmission Mechanisms, Control, and Modeling. Med. Sci. 2023, 11, 3. https://doi.org/10.3390/medsci11010003
Savi MK. An Overview of Malaria Transmission Mechanisms, Control, and Modeling. Medical Sciences. 2023; 11(1):3. https://doi.org/10.3390/medsci11010003
Chicago/Turabian StyleSavi, Merveille Koissi. 2023. "An Overview of Malaria Transmission Mechanisms, Control, and Modeling" Medical Sciences 11, no. 1: 3. https://doi.org/10.3390/medsci11010003
APA StyleSavi, M. K. (2023). An Overview of Malaria Transmission Mechanisms, Control, and Modeling. Medical Sciences, 11(1), 3. https://doi.org/10.3390/medsci11010003