Potential Indicator Value of Subfossil Gastropods in Assessing the Ecological Health of the Middle and Lower Reaches of the Yangtze River Floodplain System (China)


 ,
,
Abstract
1. Introduction
2. Materials and Methods
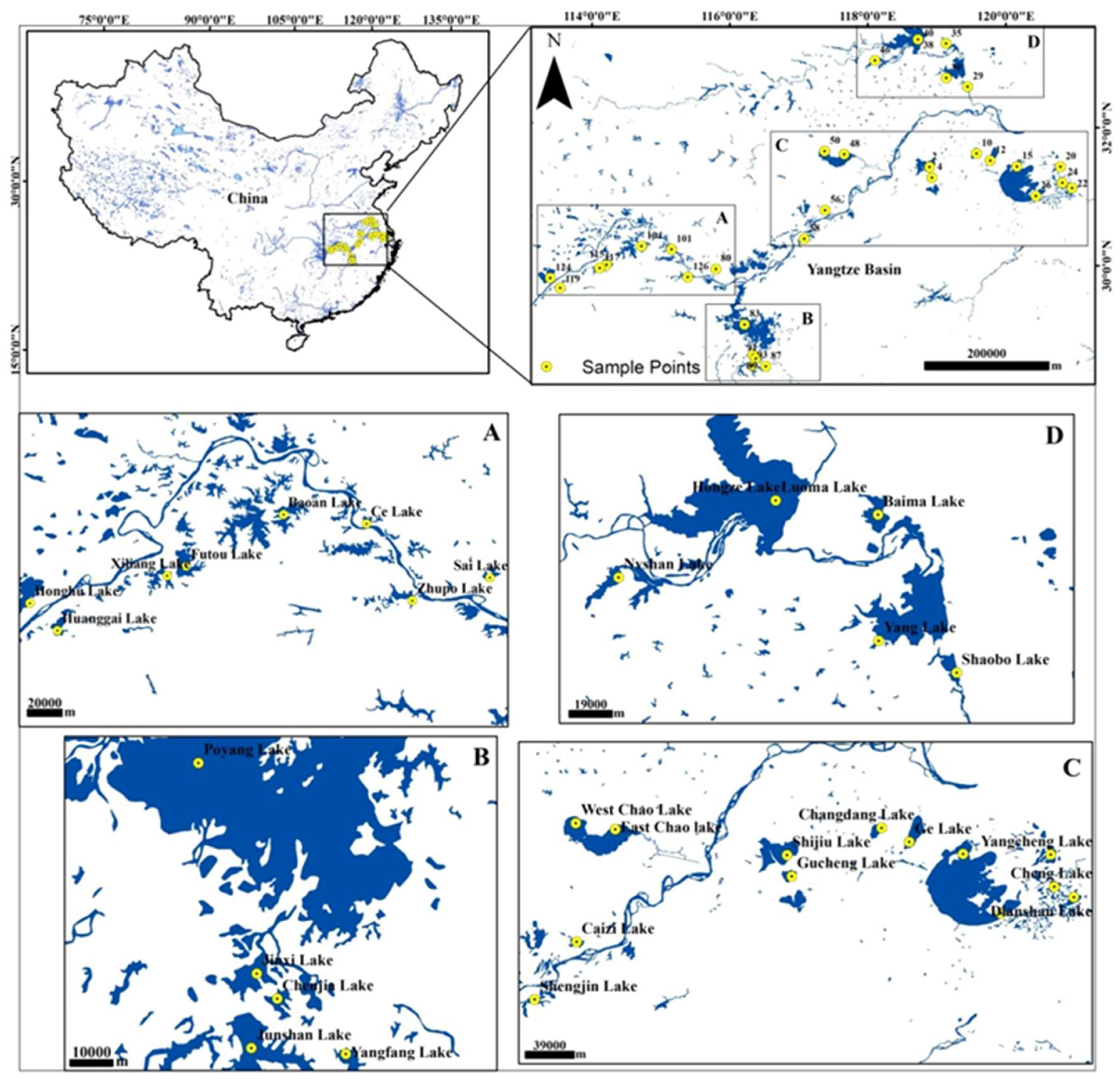
2.1. Study Area
2.2. Sediment Collection
2.3. Physico-Chemical Analysis of Water
2.4. Gastropod Subfossil Analysis
2.5. Numerical Assessment of Lake Ecosystems Health
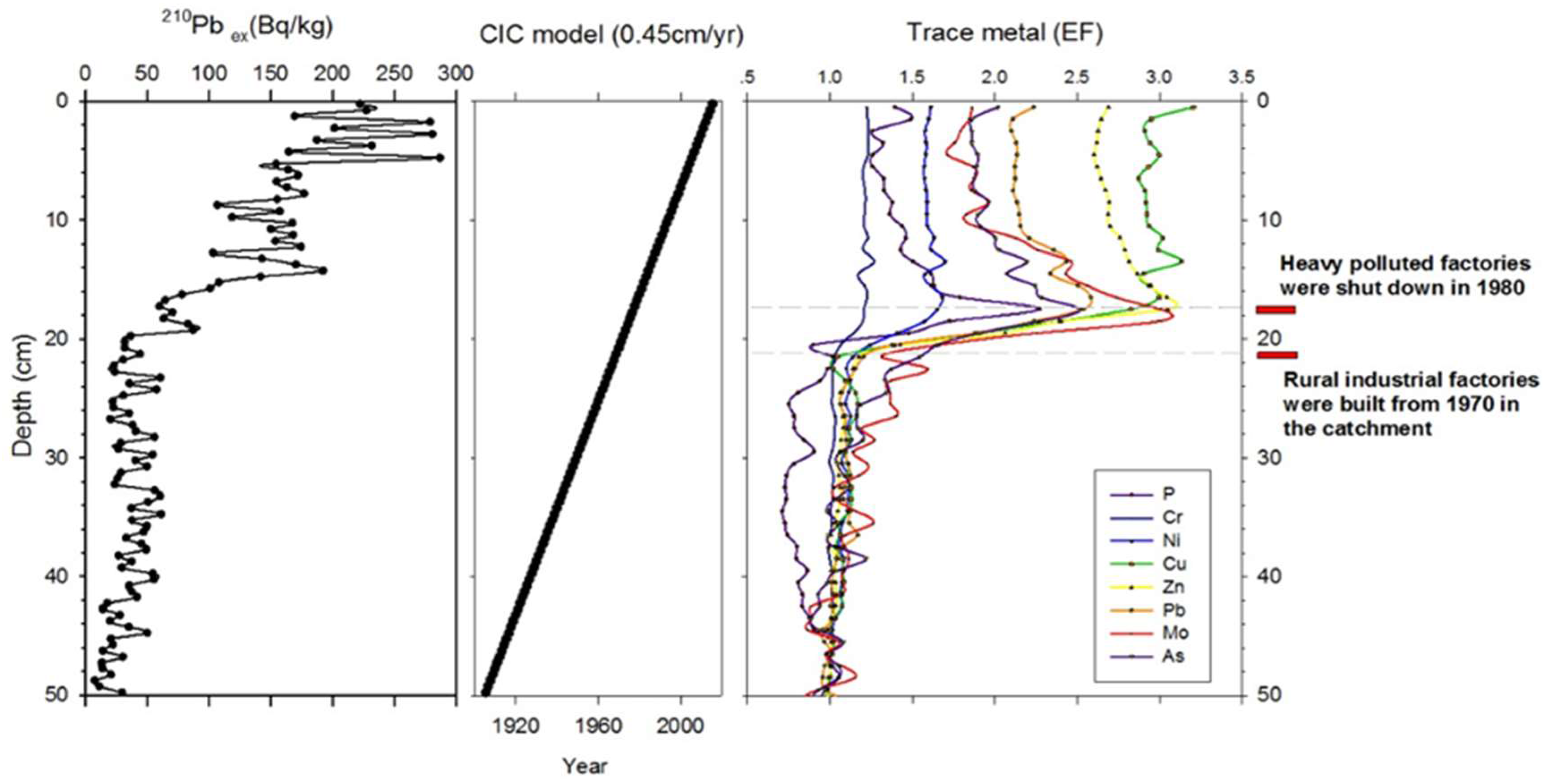
2.6. Development of Age-Depth Model
3. Results
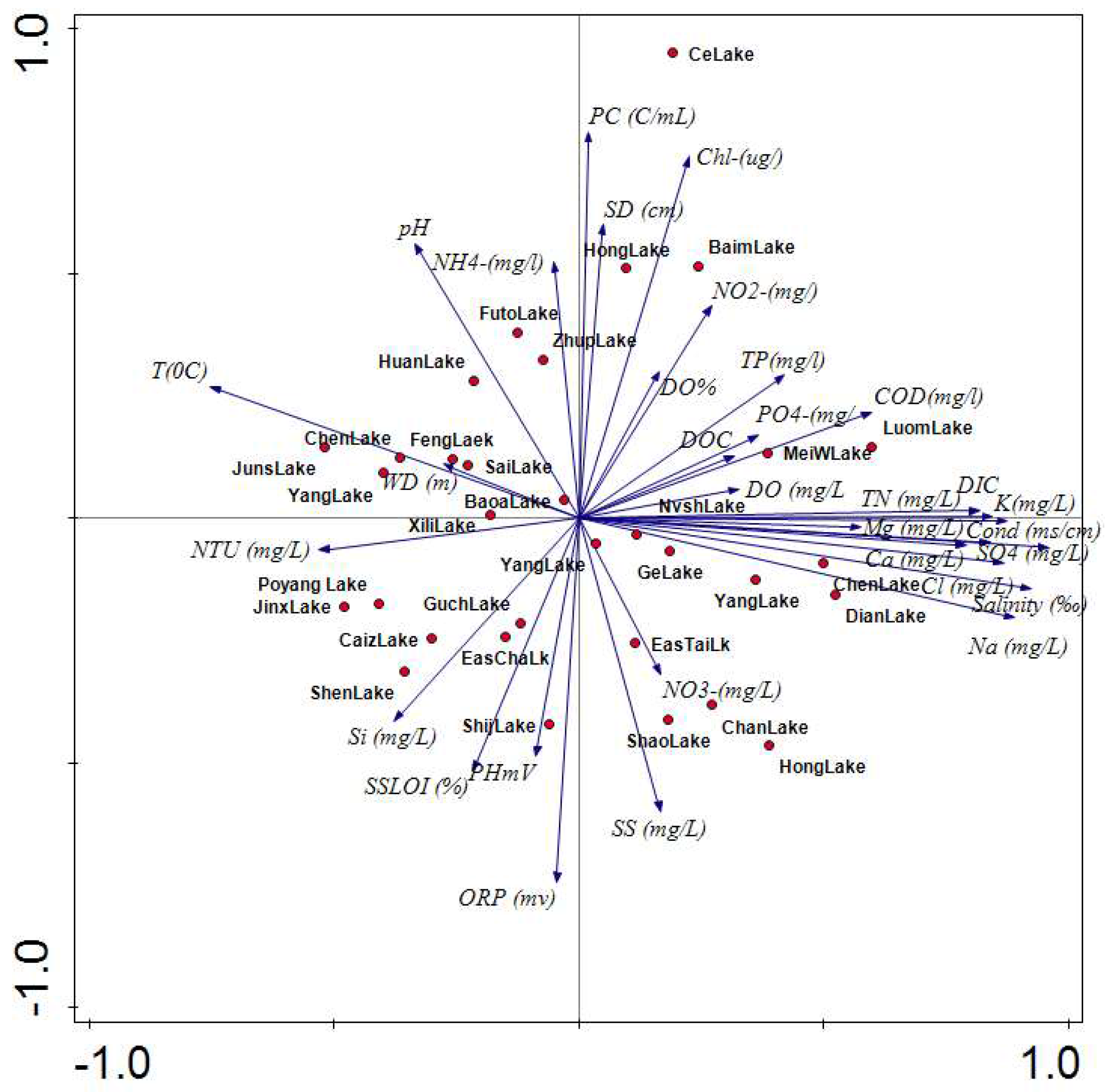
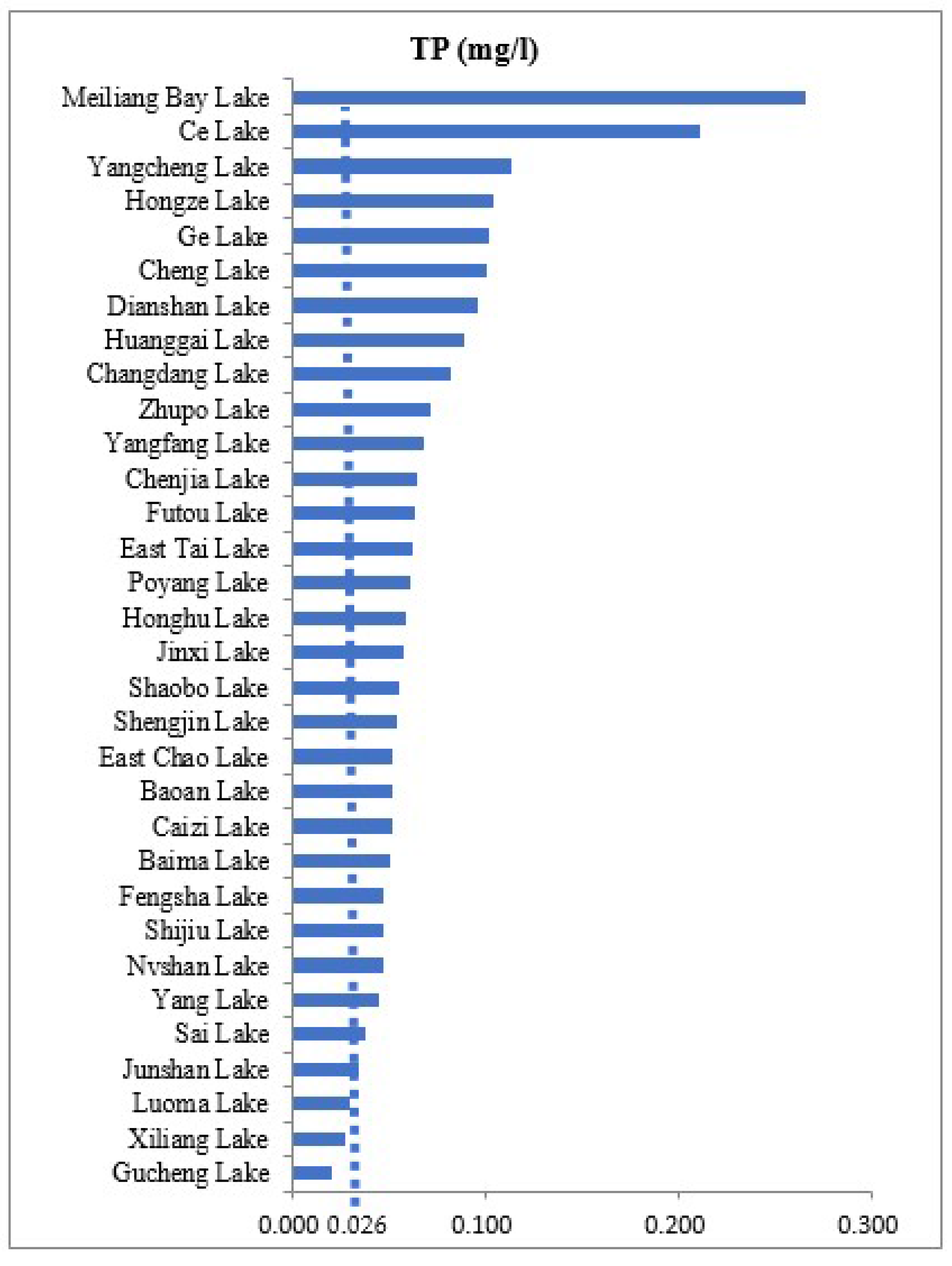
3.1. Environmental Gradients
3.2. Detrended Corespondence Analysis (DCA)
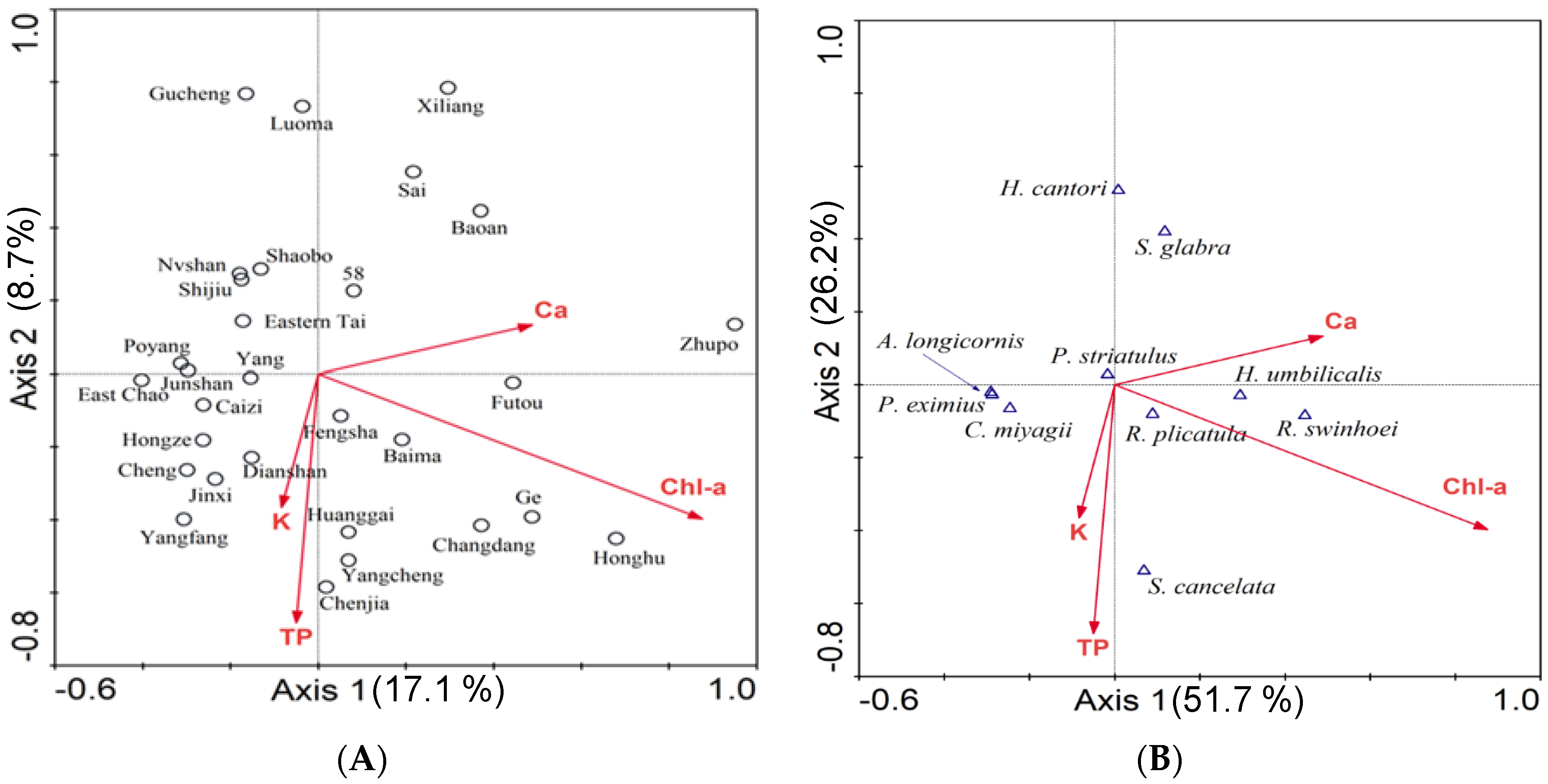
3.3. Canonical Correspondence Analysis (CCA)
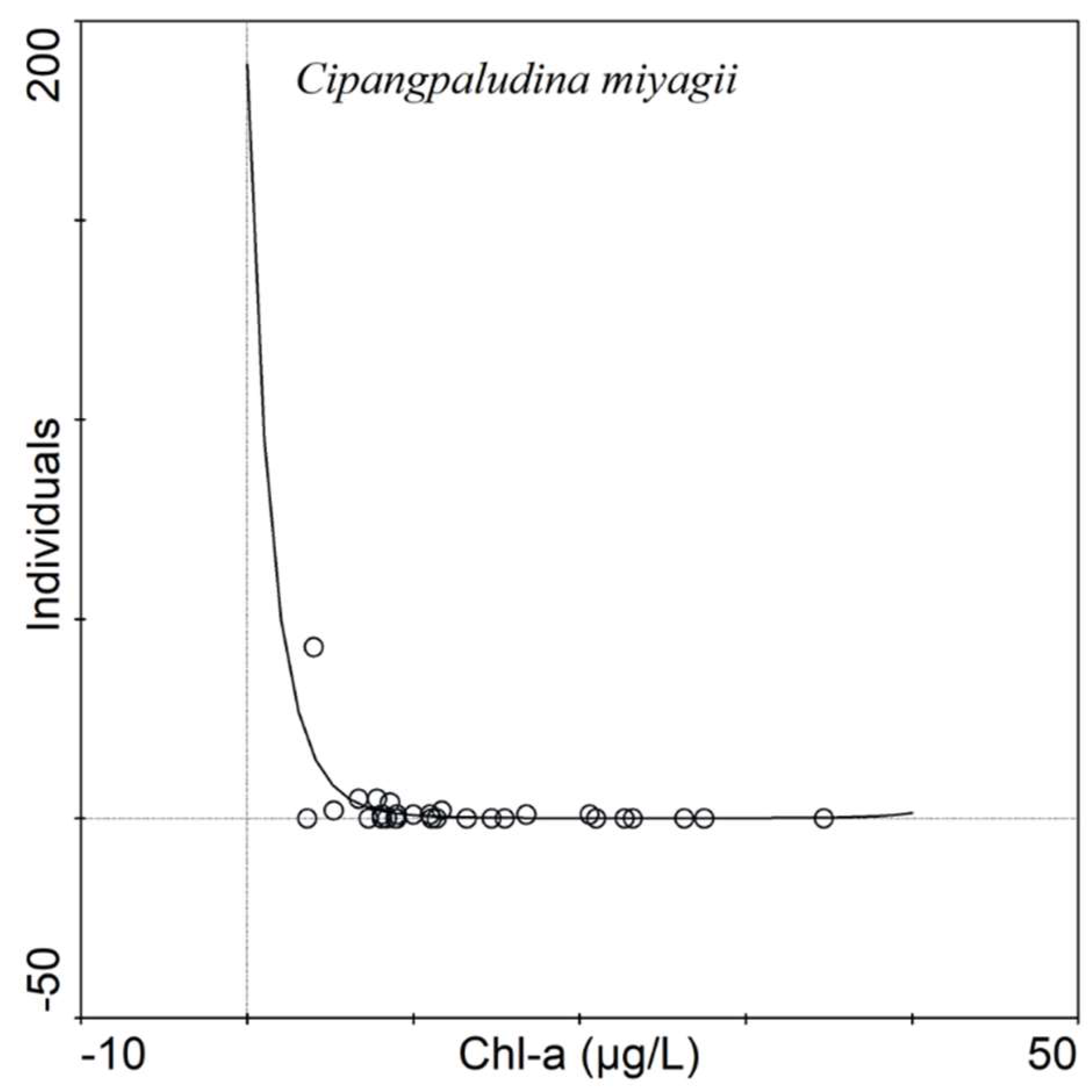
3.4. Generalized Linear Model (GLM)
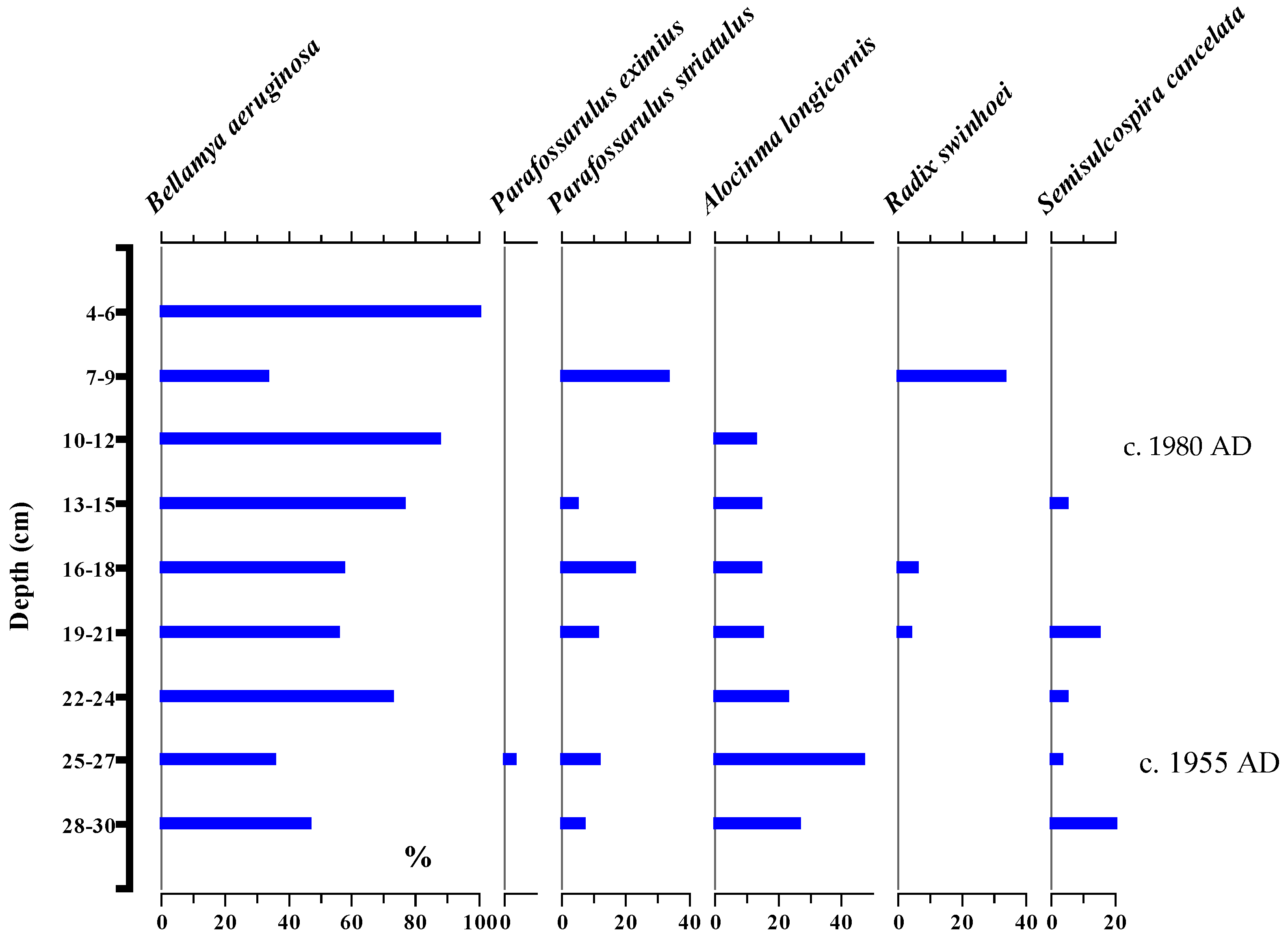
3.5. Subfossil Gastropod Stratigraphy
4. Discussion
4.1. Gastropods and Aquatic Ecosystem Health in the Middle and Lower Reaches of the Yangtze River Floodplain Lake System
4.2. Community Level Response to Lake Ecosystem Health
4.3. Population Level Response of Lake-Ecosystem Health
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costanza, R.; Mageau, M. What is a healthy ecosystem? Aquat. Ecol. 1999, 33, 105–115. [Google Scholar] [CrossRef]
- Yan, Y.; Zhao, C.; Wang, C.; Shan, P.; Zhang, Y.; Wu, G. Ecosystem health assessment of the Liao River basin upstream region based on ecosystem services. Acta Ecol. Sin. 2016, 36, 294–300. [Google Scholar] [CrossRef]
- Costanza, R. Ecosystem health and ecological engineering. Ecol. Eng. 2012, 45, 24–29. [Google Scholar] [CrossRef]
- Liu, X.-Q.; Gippel, C.J.; Wang, H.-Z.; Leigh, C.; Jiang, X.-H. Assessment of the ecological health of heavily utilized, large lowland rivers: Example of the lower Yellow River, China. Limnology 2016, 18, 17–29. [Google Scholar] [CrossRef]
- Cui, L.; Gao, C.; Zhao, X.; Ma, Q.; Zhang, M.; Li, W.; Song, H.; Wang, Y.; Li, S.; Zhang, Y. Dynamics of the lakes in the middle and lower reaches of the Yangtze River basin, China, since late nineteenth century. Environ. Monit. Assess. 2013, 185, 4005–4018. [Google Scholar] [CrossRef] [PubMed]
- Kattel, G.R.; Dong, X.; Yang, X. A century-scale, human-induced ecohydrological evolution of wetlands of two large River basins in Australia (Murray) and China (Yangtze). Hydrol. Earth Syst. Sci. 2016, 20, 2151–2168. [Google Scholar] [CrossRef]
- Hu, L.; Chao, Z.; Gu, M.; Li, F.; Chen, L.; Liu, B.; Li, X.; Huang, Z.; Li, Y.; Xing, B.; et al. Evidence for a neolithic age fire-irrigation paddy cultivation system in the lower Yangtze River delta, China. J. Archaeol. Sci. 2013, 40, 72–78. [Google Scholar] [CrossRef]
- Wu, J.; Zeng, H.; Yu, H.; Ma, L.; Xu, L.; Qin, B. Water and sediment quality in lakes along the middle and lower reaches of the Yangtze River, China. Water Resour. Manag. 2012, 26, 3601–3618. [Google Scholar] [CrossRef]
- Yang, S.L.; Xu, K.H.; Milliman, J.D.; Yang, H.F.; Wu, C.S. Decline of Yangtze River water and sediment discharge: Impact from natural and anthropogenic changes. Sci. Rep. 2015, 5, 12581. [Google Scholar] [CrossRef] [PubMed]
- Du, L.-N.; Li, Y.; Chen, X.-Y.; Yang, J.-X. Effect of eutrophication on molluscan community composition in the lake Dianchi (China, Yunnan). Limnologica 2011, 41, 213–219. [Google Scholar] [CrossRef]
- Bouetard, A.; Cote, J.; Besnard, A.L.; Collinet, M.; Coutellec, M.A. Environmental versus anthropogenic effects on population adaptive divergence in the freshwater snail Lymnaea stagnalis. PLoS ONE 2014, 9, e106670. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.; An, S.; Gu, B. Assessment of ecosystem health during the past 40 years for lake Taihu in the Yangtze River delta, China. Limnology 2010, 12, 47–53. [Google Scholar] [CrossRef]
- Cai, Y.; Jiang, J.; Zhang, L.; Chen, Y.; Gong, Z. Simplification of macrozoobenthic assemblages related to anthropogenic eutrophication and cyanobacterial blooms in two large shallow subtropical lakes in China. Aquat. Ecosyst. Health Manag. 2012, 15, 81–91. [Google Scholar] [CrossRef]
- Yang, X.; Anderson, N.J.; Dong, X.; Shen, J.I. Surface sediment diatom assemblages and epilimnetic total phosphorus in large, shallow lakes of the Yangtze floodplain: Their relationships and implications for assessing long-term eutrophication. Freshw. Biol. 2008, 53, 1273–1290. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Y.; Singh, V.P.; Zhu, J.; Jiang, L.; Zeng, D.; Liu, D.; Zeng, X.; Wu, J.; Wang, L.; et al. Ecological and health risk assessment of PAHS, OCPS, and PCBS in Taihu lake basin. Ecol. Indic. 2017, in press. [Google Scholar] [CrossRef]
- Guo, C.; Ye, S.; Lek, S.; Liu, J.; Zhang, T.; Yuan, J.; Li, Z. The need for improved fishery management in a shallow macrophytic lake in the Yangtze River basin: Evidence from the food web structure and ecosystem analysis. Ecol. Model. 2013, 267, 138–147. [Google Scholar] [CrossRef]
- Tong, Y.; Zhang, W.; Wang, X.; Couture, R.-M.; Larssen, T.; Zhao, Y.; Li, J.; Liang, H.; Liu, X.; Bu, X.; et al. Decline in Chinese lake phosphorus concentration accompanied by shift in sources since 2006. Nat. Geosci. 2017, 10, 507–511. [Google Scholar] [CrossRef]
- Zhou, Y.; Ma, J.; Zhang, Y.; Qin, B.; Jeppesen, E.; Shi, K.; Brookes, J.D.; Spencer, R.G.M.; Zhu, G.; Gao, G. Improving water quality in China: Environmental investment pays dividends. Water Res. 2017, 118, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-J.; Pan, B.-Z.; Liang, X.-M.; Wang, H.-Z. Gastropods on submersed macrophytes in Yangtze lakes: Community characteristics and empirical modelling. Int. Rev. Hydrobiol. 2006, 91, 521–538. [Google Scholar] [CrossRef]
- Cai, Y.; Zhang, Y.; Wu, Z.; Chen, Y.; Xu, J.; Gong, Z. Composition, diversity, and environmental correlates of benthic macroinvertebrate communities in the five largest freshwater lakes of China. Hydrobiologia 2016, 788, 85–98. [Google Scholar] [CrossRef]
- Gérard, C.; Carpentier, A.; Paillisson, J.-M. Long-term dynamics and community structure of freshwater gastropods exposed to parasitism and other environmental stressors. Freshw. Biol. 2008, 53, 470–484. [Google Scholar] [CrossRef]
- Cai, Y.; Gong, Z.; Qin, B. Benthic macroinvertebrate community structure in lake Taihu, China: Effects of trophic status, wind-induced disturbance and habitat complexity. J. Great Lakes Res. 2012, 38, 39–48. [Google Scholar] [CrossRef]
- Cao, Y.; Li, W.; Jeppesen, E. The response of two submerged macrophytes and periphyton to elevated temperatures in the presence and absence of snails: A microcosm approach. Hydrobiologia 2014, 738, 49–59. [Google Scholar] [CrossRef]
- Underwood, G.J.C.; Thomas, J.D.; Baker, J.H. An experimental investigation of interactions in snail-macrophyte-epiphyte systems. Oecologia 1992, 91, 587–595. [Google Scholar] [CrossRef] [PubMed]
- McCollum, E.W.; Crowder, L.B.; McCollum, S.A. Complex interactions of fish, snails, and littoral zone periphyton. Ecology 1998, 79, 1980–1994. [Google Scholar] [CrossRef]
- Donohue, I.; Irvine, K. Size-specific effects of increased sediment loads on gastropod communities in Lake Tanganyika, Africa. Hydrobiologia 2004, 522, 337–342. [Google Scholar] [CrossRef]
- Koopman, K.R.; Collas, F.P.L.; van der Velde, G.; Verberk, W.C.E.P. Oxygen can limit heat tolerance in freshwater gastropods: Differences between gill and lung breathers. Hydrobiologia 2015, 763, 301–312. [Google Scholar] [CrossRef]
- Zheng, S.; Zhou, Q. Intoxication and biochemical responses of freshwater snail Bellamya aeruginosa to ethylbenzene. Environ. Sci. Pollut. Res. 2017, 24, 189–198. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, P.B.; Mitchell, E.; France, K.; Rivers, A.; Hakizimna, P.; Cohen, A.S. Individual and assemblage level effects of anthropogenic sedimentation on snails in Lake Tanganyika. Conserv. Biol. 2005, 19, 171–181. [Google Scholar] [CrossRef]
- Lodge, D.M.; Kelly, P. Habitat disturbance and the stability of freshwater gastropod populations. Oecologia 1985, 68, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Gérard, C.; Brient, L.; Rouzic, B.L. Variation in the response of juvenile and adult gastropods (Lymnaea stagnalis) to cyanobacterial toxin (microcystin-LR). Environ. Toxicol. 2005, 20, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Lance, E.; Paty, C.; Bormans, M.; Brient, L.; Gerard, C. Interactions between cyanobacteria and gastropods ii. Impact of toxic Planktothrix agardhii on the life-history traits of Lymnaea stagnalis. Aquat. Toxicol. 2007, 81, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Horsak, M. Mollusc community patterns and species response curves along a mineral richness gradient: A case study in fens. J. Biogeogr. 2006, 33, 98–107. [Google Scholar] [CrossRef]
- Conley, D.J.; Carstensen, J.; Vaquer-Sunyer, R.; Duarte, C.M. Ecosystem thresholds with hypoxia. Hydrobiologia 2009, 629, 21–29. [Google Scholar] [CrossRef]
- Fink, P.; Von Elert, E. Physiological responses to stoichiometric constraints: Nutrient limitation and compensatory feeding in a freshwater snail. Oikos 2006, 115, 484–494. [Google Scholar] [CrossRef]
- Elser, J.J.; Schampel, J.H.; Kyle, M.; Watts, J.; Carson, E.W.; Dowling, T.E.; Tang, C.; Roopnarine, P.D. Response of grazing snails to phosphorus enrichment of modern stromatolitic microbial communities. Freshw. Biol. 2005, 50, 1826–1835. [Google Scholar] [CrossRef]
- Liess, A.; Hillebrand, H. Role of nutrient supply in grazer–periphyton interactions: Reciprocal influences of periphyton and grazer nutrient stoichiometry. J. N. Am. Benthol. Soc. 2006, 25, 632–642. [Google Scholar] [CrossRef]
- Yin, G.; Zhou, Y.; Strid, A.; Zheng, Z.; Bignert, A.; Ma, T.; Athanassiadis, I.; Qiu, Y. Spatial distribution and bioaccumulation of polychlorinated biphenyls (PCBS) and polybrominated diphenyl ethers (PBDES) in snails (Bellamya aeruginosa) and sediments from Taihu lake area, China. Environ. Sci. Pollut. Res. 2017, 24, 7740–7751. [Google Scholar] [CrossRef] [PubMed]
- Niggebrugge, K.; Durance, I.; Watson, A.M.; Leuven, R.S.E.W.; Ormerod, S.J. Applying landscape ecology to conservation biology: Spatially explicit analysis reveals dispersal limits on threatened wetland gastropods. Biol. Conserv. 2007, 139, 286–296. [Google Scholar] [CrossRef]
- De Francesco, C.G.; Hassan, G.S. The significance of molluscs as paleoecological indicators of freshwater systems in central-western Argentina. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 274, 105–113. [Google Scholar] [CrossRef]
- Shi, W.; Yu, N.; Jiang, X.; Han, Z.; Wang, S.; Zhang, X.; Wei, S.; Giesy, J.P.; Yu, H. Influence of blooms of phytoplankton on concentrations of hydrophobic organic chemicals in sediments and snails in a hyper-eutrophic, freshwater lake. Water Res. 2017, 113, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Apolinarska, K.; Pełechaty, M.; Kossler, A. Within-sample variability of δ13C and δ18O values of freshwater gastropod shells and the optimum number of shells to measure per sediment layer in the Paddenluch palaeolacustrine sequence, Germany. J. Paleolimnol. 2015, 54, 305–323. [Google Scholar] [CrossRef]
- De Francesco, C.G.; Zárate, M.A.; Miquel, S.E. Late Pleistocene mollusc assemblages and inferred paleoenvironments from the Andean piedmont of Mendoza, Argentina. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 251, 461–469. [Google Scholar] [CrossRef]
- Chaves-Campos, J.; Coghill, L.M.; de Leon, F.J.G.; Johnson, S.G. The effect of aquatic plant abundance on shell crushing resistance in a freshwater snail. PLoS ONE 2012, 7, e44374. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, D.; Fagel, N.; Araneda, A.; Jana-Pinninghoff, P.; Keppens, E.; Urrutia, R. Late Holocene climate variability on the eastern flank of the Patagonian Andes (Chile): A δ18O record from mollusks in Lago Cisnes (47°S). Holocene 2015, 25, 1220–1230. [Google Scholar] [CrossRef]
- Yi, L.; Jiao, W.; Chen, X.; Chen, W. An overview of reclaimed water reuse in china. J. Environ. Sci. 2011, 23, 1585–1593. [Google Scholar] [CrossRef]
- Fang, J.; Wang, Z.; Zhao, S.; Li, Y.; Tang, Z.; Yu, D.; Ni, L.; Liu, H.; Xie, P.; Da, L.; et al. Biodiversity changes in the lakes of the central Yangtze. Front. Ecol. Environ. 2006, 4, 369–377. [Google Scholar] [CrossRef]
- Le, C.; Zha, Y.; Li, Y.; Sun, D.; Lu, H.; Yin, B. Eutrophication of lake waters in China: Cost, causes, and control. Environ. Manag. 2010, 45, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Duan, H.; Hu, C.; Feng, X.; Li, A.; Ju, W.; Jiang, J.; Yang, G. A half-century of changes in China’s lakes: Global warming or human influence? Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef]
- Wang, S.; Jin, X.; Jiao, L.; Wu, F. Nitrogen fractions and release in the sediments from the shallow lakes in the middle and lower reaches of the Yangtze River area, China. Water Air Soil Pollut. 2007, 187, 5–14. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, L.; Kong, L.; Liu, E.; Wang, L.; Zhu, J. Spatial distribution, ecological risk assessment and source identification for heavy metals in surface sediments from Dongping Lake, Shandong, East China. Catena 2015, 125, 200–205. [Google Scholar] [CrossRef]
- Xu, M.; Dong, X.; Yang, X.; Wang, R.; Zhang, K.; Zhao, Y.; Davidson, T.A.; Jeppesen, E. Using palaeolimnological data and historical records to assess long-term dynamics of ecosystem services in typical Yangtze shallow lakes (China). Sci. Total Environ. 2017, 584–585, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Renberg, I. The HON-Kajak sediment corer. J. Paleolimnol. 1991, 6, 167–170. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Zhang, W.Z.; Wang, Y.X.; Wang, E.Y. Economic Fauna of China; Science Press: Beijing, China, 1979. (In Chinese) [Google Scholar]
- He, J.; Zhang, Z. The Freshwater Bivalves of China; Conch Books: Beijing, China, 2013. [Google Scholar]
- Ahmadi-Nedushan, B.; St-Hilaire, A.; Bérubé, M.; Robichaud, É.; Thiémonge, N.; Bobée, B. A review of statistical methods for the evaluation of aquatic habitat suitability for instream flow assessment. River Res. Appl. 2006, 22, 503–523. [Google Scholar] [CrossRef]
- Choudhary, P.; Routh, J.; Chakrapani, G.J. An environmental record of changes in sedimentary organic matter from Lake Sattal in Kumaun Himalayas, India. Sci. Total Environ. 2009, 407, 2783–2795. [Google Scholar] [CrossRef] [PubMed]
- Bos, D.G.; Cumming, B.F. Sedimentary cladoceran remains and their relationship to nutrients and other limnological variables in 53 lakes from British Columbia, Canada. Can. J. Fish. Aquat. Sci. 2003, 60, 1177–1189. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and Canodraw for Window’s User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Jones, J.I.; Sayer, C.D. Does the fish–invertebrate–periphyton cascade precipitate plant loss in shallow lakes? Ecology 2003, 84, 2155–2167. [Google Scholar] [CrossRef]
- Liess, A.; Hillebrand, H. Invited review: Direct and indirect effects in herbivore-periphyton interactions. Arch. Hydrobiol. 2004, 159, 433–453. [Google Scholar] [CrossRef]
- Malzahn, A.M.; Hantzsche, F.; Schoo, K.L.; Boersma, M.; Aberle, N. Differential effects of nutrient-limited primary production on primary, secondary or tertiary consumers. Oecologia 2010, 162, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, P.; Guo, L.; Zheng, L.; Ni, L. Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and -RR in a freshwater snail (Bellamya aeruginosa) from a large shallow, eutrophic lake of the subtropical China. Environ. Pollut. 2005, 134, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Xie, P.; Liu, Y.; Qiu, T. Transfer, distribution and bioaccumulation of microcystins in the aquatic food web in Lake Taihu, China, with potential risks to human health. Sci. Total Environ. 2009, 407, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Tang, T.; Xie, Z.; Ye, L.; Li, D.; Cai, Q. Distribution of the macroinvertebrate communities in the Xiangxi River system and relationships with environmental factors. J. Freshw. Ecol. 2005, 20, 233–238. [Google Scholar] [CrossRef]
- Scheifhacken, N.; Fiek, C.; Rothhaupt, K.O. Complex spatial and temporal patterns of littoral benthic communities interacting with water level fluctuations and wind exposure in the littoral zone of a large lake. Fundam. Appl. Limnol. 2007, 169, 115–129. [Google Scholar] [CrossRef]
- Dos Santos, A.M.; Thomaz, S. Aquatic macrophytes diversity in lagoons of a tropical floodplain: The role of connectivity and water level. Austral Ecol. 2007, 32, 177–190. [Google Scholar] [CrossRef]
- White, M.S.; Xenopoulos, M.A.; Metcalfe, R.A.; Somers, K.M. On the role of natural water level fluctuation in structuring littoral benthic macroinvertebrate community composition in lakes. Limnol. Oceanogr. 2010, 55, 2275–2284. [Google Scholar] [CrossRef]
- Yongjiu, C.; Xuea, Q.; Xub, J.; Zhanga, L.; Gonga, Z.; Achraya, K. Widespread natural intraspecific variation in tissue stoichiometryof two freshwater molluscs: Effect of nutrient enrichment. Ecol. Indic. 2016, 66, 583–591. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Latitude (North) | Longitude (East) | Mean Lake Depth (m) | SD (cm) | Temp. (°C) | Cond (ms/cm) | pH | NTU (mg/L) | DO % | Salinity (‰) | TN (mg/L) | TP (mg/L) | Si (mg/L) | Chl-a (ug/L) | SS (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shijiu Lake | 31°25′57.20″ | 118°53′47.50″ | 1.92 | 12.23 | 17.19 | 0.25 | 7.84 | 3.4 | 74.2 | 0.12 | 1.84 | 0.04 | 2.04 | 7.3 | 24.09 |
| Gucheng Lake | 31°16′48.70″ | 118°55′48.55″ | 3.15 | 40.6 | 16.96 | 0.26 | 8.18 | 5.46 | 322.0 | 0.14 | 1.31 | 0.02 | 2.62 | 3.6 | 9.08 |
| Changdang Lake | 31°37′42.11″ | 119°34′35.07″ | 1.57 | 6.60 | 16.69 | 0.44 | 7.90 | 15.3 | 316.17 | 0.22 | 1.92 | 0.08 | 2.57 | 26.3 | 42.72 |
| Ge Lake | 31°31′38.35″ | 119°46′34.24″ | 1.80 | 12.63 | 17.19 | 0.46 | 8.24 | 87.63 | 321.85 | 0.22 | 2.85 | 0.10 | 2.59 | 22.7 | 31.06 |
| Meiliang Bay Lake | 31°26′30.46″ | 120°09′46.56″ | 2.82 | 15.36 | 17.65 | 0.45 | 8.53 | 31.06 | 343.45 | 0.22 | 4.81 | 0.26 | 2.50 | 33.6 | 36.37 |
| Yangcheng Lake | 31°26′8.89″ | 120°47′40.40″ | 1.70 | 26.45 | 16.84 | 0.50 | 7.95 | 25.36 | 323.37 | 0.24 | 2.41 | 0.11 | 1.82 | 15.5 | 14.57 |
| Dianshan Lake | 31°07′48.55″ | 120°57′39.48″ | 2.50 | 26.66 | 16.96 | 0.58 | 8.15 | 19.36 | 318.2 | 0.28 | 2.62 | 0.09 | 2.10 | 11.7 | 21.95 |
| Cheng Lake | 31°12′18.43″ | 120°49′15.68″ | 2.17 | 31.32 | 17.09 | 0.53 | 8.20 | 13.23 | 320.6 | 0.26 | 3.09 | 0.10 | 2.21 | 8.6 | 9.62 |
| East Tai Lake | 31°00′45.71″ | 120°26′8.74″ | 1.62 | 16.94 | 16.99 | 0.35 | 8.12 | 29.63 | 323.77 | 0.17 | 1.34 | 0.06 | 1.26 | 8.1 | 48.07 |
| Shaobo Lake | 32°35′51.34″ | 119°26′49.06″ | 1.87 | 15.08 | 15.87 | 0.40 | 7.98 | 46.82 | 320.72 | 0.19 | 1.76 | 0.05 | 2.73 | 7.8 | 63.09 |
| Yang Lake | 32°43′29.77″ | 119°08′13.86″ | 2.22 | 22.44 | 14.31 | 0.24 | 8.25 | 29.03 | 314.52 | 0.13 | 1.21 | 0.04 | 1.93 | 13.2 | 27.86 |
| Baima Lake | 33°13′36.34″ | 119°08′3.59″ | 1.65 | 50.6 | 16.49 | 0.43 | 9.08 | 24.05 | 335.55 | 0.10 | 1.76 | 0.05 | 2.35 | 23.2 | 22.00 |
| Hongze Lake | 33°17′0.93″ | 118°43′37.38″ | 2.82 | 11.33 | 15.24 | 0.50 | 8.17 | 79.7 | 320.17 | 0.24 | 2.13 | 0.10 | 3.16 | 5.2 | 105.15 |
| Luoma Lake | 33°17′0.93″ | 118°43′37.38″ | 3.87 | 48.66 | 16.02 | 0.69 | 8.45 | 27.1 | 323.27 | 0.34 | 1.14 | 0.03 | 1.06 | 8.4 | 80.00 |
| Nvshan Lake | 32°58′38.22″ | 118°06′7.25″ | 3.70 | 27.39 | 16.23 | 0.35 | 8.45 | 19.2 | 319.37 | 0.17 | 1.36 | 0.04 | 1.53 | 8.1 | 16.73 |
| East Chao lake | 31°37′2.30″ | 117°39′30.30″ | 3.77 | 12.28 | 18.49 | 0.27 | 8.35 | 42.47 | 322.92 | 0.13 | 1.43 | 0.05 | 3.16 | 6.7 | 23.48 |
| West Chao Lake | 31°39′45.51″ | 117°22′29.42″ | 3.12 | 36.51 | 19.21 | 0.21 | 8.70 | 22.37 | 327.9 | 0.10 | 1.32 | 0.04 | 2.83 | 16.8 | 15.23 |
| Caizi Lake | 30°48′29.22″ | 117°22‘33.11″ | 2.53 | 9.03 | 18.29 | 0.13 | 8.42 | 69.07 | 320.3 | 0.06 | 1.53 | 0.05 | 2.50 | 8.9 | 37.82 |
| Shengjin Lake | 30°23′32.34″ | 117°04′43.72″ | 2.18 | 8.73 | 19.76 | 0.17 | 8.37 | 94.32 | 311.35 | 0.09 | 1.21 | 0.05 | 2.24 | 9.0 | 38.81 |
| Sai Lake | 29°57′18.30″ | 115°47′49.05″ | 3.60 | 37.37 | 19.17 | 0.261 | 8.68 | 18.2 | 330.5 | 0.12 | 0.85 | 0.03 | 2.47 | 11.1 | 13.27 |
| Poyang Lake | 29°09′0.18″ | 116°12′47.72″ | 3.43 | 9.38 | 21.09 | 0.14 | 9.09 | 27.8 | 114.96 | 0.06 | 1.54 | 0.06 | 4.16 | 4.0 | 33.07 |
| Yangfang Lake | 28°32′27.43″ | 116°31′18.41″ | 1.92 | 35.15 | 21.74 | 0.14 | 8.55 | 119.3 | 330.45 | 0.07 | 1.48 | 0.06 | 1.91 | 10.0 | 18.66 |
| Jinxi Lake | 28°42′33.21″ | 116°20′6.90″ | 2.67 | 11.31 | 19.82 | 0.12 | 8.56 | 149.92 | 326.4 | 0.06 | 1.26 | 0.05 | 4.12 | 11.0 | 23.52 |
| Chenjia Lake | 28°39′23.40″ | 116°22′42.08″ | 3.10 | 18.05 | 19.56 | 0.18 | 8.30 | 151.25 | 335.7 | 0.09 | 0.76 | 0.06 | 1.67 | 20.6 | 12.32 |
| Junshan Lake | 28°33′11.38″ | 116°19′26.20″ | 4.45 | 59.37 | 20.51 | 0.08 | 8.56 | 137.37 | 330.85 | 0.04 | 0.63 | 0.03 | 2.41 | 9.0 | 6.55 |
| Ce Lake | 30°14′14.40″ | 115°9′9.52″ | 2.55 | 33.73 | 18.65 | 0.29 | 8.53 | 21.57 | 311.77 | 0.14 | 2.11 | 0.21 | 1.12 | 51.9 | 12.38 |
| Baoan Lake | 30°17′5.62″ | 114°43′11.59″ | 3.30 | 20.45 | 19.76 | 0.29 | 8.07 | 13.02 | 333.42 | 0.14 | 0.94 | 0.05 | 1.77 | 15.5 | 15.62 |
| Futou Lake | 30°00′52.05″ | 114°12′40.47″ | 2.42 | 25.62 | 19.51 | 0.27 | 8.78 | 68.75 | 332.05 | 0.13 | 1.94 | 0.06 | 2.20 | 21.0 | 14.33 |
| Xiliang Lake | 29°57′57.32″ | 114°06′40.92″ | 1.97 | 27.12 | 19.25 | 0.25 | 8.00 | 44.95 | 314.55 | 0.12 | 1.24 | 0.02 | 2.59 | 11.4 | 8.76 |
| Huanggai Lake | 29°40′44.75″ | 113°32′21.37″ | 2.60 | 21.82 | 18.82 | 0.16 | 8.57 | 28.7 | 320.82 | 0.08 | 2.18 | 0.08 | 3.00 | 14.7 | 10.39 |
| Honghu Lake | 29°49′22.86″ | 113°23′46.55″ | 1.52 | 34.36 | 18.74 | 0.34 | 8.56 | 30.32 | 338.27 | 0.16 | 1.33 | 0.06 | 1.54 | 34.7 | 15.85 |
| Zhupo Lake | 29°50′9.56″ | 115°23′28.83″ | 3.95 | 46.32 | 18.14 | 0.32 | 8.42 | 19.12 | 326.1 | 0.16 | 0.92 | 0.07 | 1.24 | 27.5 | 10.56 |
| Gastropod (Snail) Species | Mean | Min | Max |
|---|---|---|---|
| Cipangopaludinamiyagii (Kuroda) | 5.30 ± 11.42 | 1 | 43 |
| Parafossarulus eximius (Frauenfeld) | 1.00 ± 0 | 1 | 1 |
| Parafossarulus striatulus (Benson) | 3.41 ± 3.96 | 1 | 12 |
| Alocinma longicornis (Benson) | 4.00 ± 1.41 | 3 | 5 |
| Radix swinhoei (Adams) | 2.00 ± 1.15 | 1 | 4 |
| Radix plicatula (Benson) | 3.18 ± 2.08 | 1 | 6 |
| Stenothyra glabra (Adams) | 1.00 ± 0 | 1 | 1 |
| Bradybaena ravida (Benson) | 1.00 ± 0 | 1 | 1 |
| Semisulcospira cancelata (Benson) | 1.00 ± 0 | 1 | 1 |
| Hippeutis umbilicalis (Benson) | 1.00 ± 0 | 1 | 1 |
| Hippeutis cantori (Benson) | 1.00 ± 0 | 1 | 1 |
| Pulmonate | 6.18 ± 12.29 | 1 | 69 |
| Nonpulmonate | 1.96 ± 1.42 | 1 | 7 |
| DCA Axes | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Eigenvalues (λ) | 0.635 | 0.267 | 0.116 | 0.073 |
| Length of gradient | 4.255 | 3.006 | 2.479 | 2.107 |
| Cumulative percentage variance of species data | 26.0 | 36.9 | 41.6 | 44.6 |
| Sum of all unconstrained eigenvalues = 2.455 | ||||
| DCA axes | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Species-environment correlations | 0.850 | 0.637 | 0.703 | 0.492 |
| Cumulative percentage variance of species data (%) | 17.1 | 25.8 | 30.6 | 33.1 |
| Cumulative percentage variance of species-environment (%) | 51.7 | 77.9 | 92.3 | 100 |
| Sum of all eigenvalues = 2.415 | ||||
| Sum of all canonical eigen values = 0.800 | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kattel, G.; Cai, Y.; Yang, X.; Zhang, K.; Hao, X.; Wang, R.; Dong, X. Potential Indicator Value of Subfossil Gastropods in Assessing the Ecological Health of the Middle and Lower Reaches of the Yangtze River Floodplain System (China). Geosciences 2018, 8, 222. https://doi.org/10.3390/geosciences8060222
Kattel G, Cai Y, Yang X, Zhang K, Hao X, Wang R, Dong X. Potential Indicator Value of Subfossil Gastropods in Assessing the Ecological Health of the Middle and Lower Reaches of the Yangtze River Floodplain System (China). Geosciences. 2018; 8(6):222. https://doi.org/10.3390/geosciences8060222
Chicago/Turabian StyleKattel, Giri, Yongjiu Cai, Xiangdong Yang, Ke Zhang, Xu Hao, Rong Wang, and Xuhui Dong. 2018. "Potential Indicator Value of Subfossil Gastropods in Assessing the Ecological Health of the Middle and Lower Reaches of the Yangtze River Floodplain System (China)" Geosciences 8, no. 6: 222. https://doi.org/10.3390/geosciences8060222
APA StyleKattel, G., Cai, Y., Yang, X., Zhang, K., Hao, X., Wang, R., & Dong, X. (2018). Potential Indicator Value of Subfossil Gastropods in Assessing the Ecological Health of the Middle and Lower Reaches of the Yangtze River Floodplain System (China). Geosciences, 8(6), 222. https://doi.org/10.3390/geosciences8060222