
Vertebrate Skeletal Remains as Paleohydrologic Proxies: Complex Hydrologic Setting in the Upper Cretaceous Kaiparowits Formation


Abstract
1. Introduction
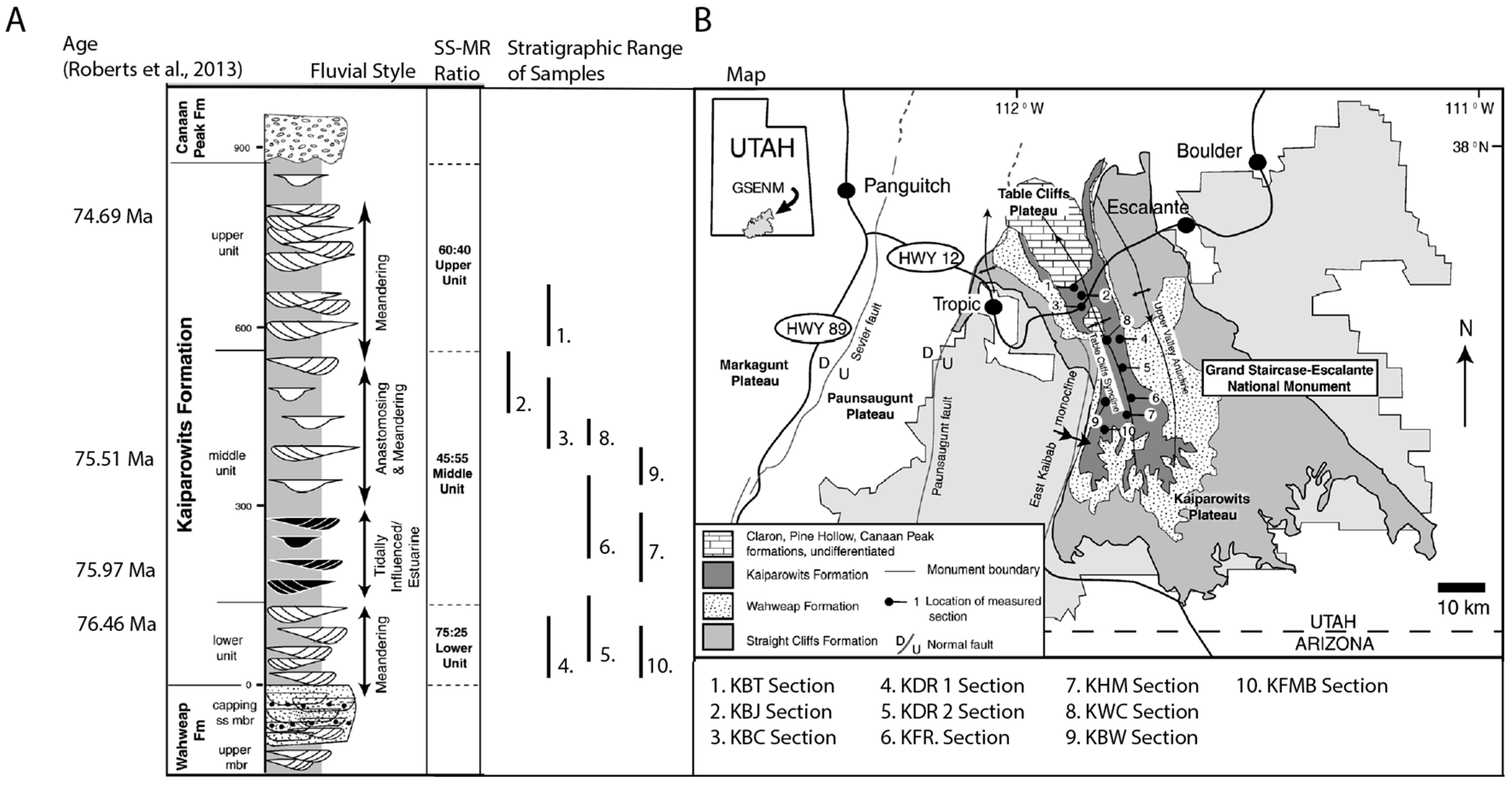
2. Geologic Setting
3. Materials and Methods
3.1. Materials
3.2. Stable Isotope Analysis
3.3. Statistical Analyses
4. Results
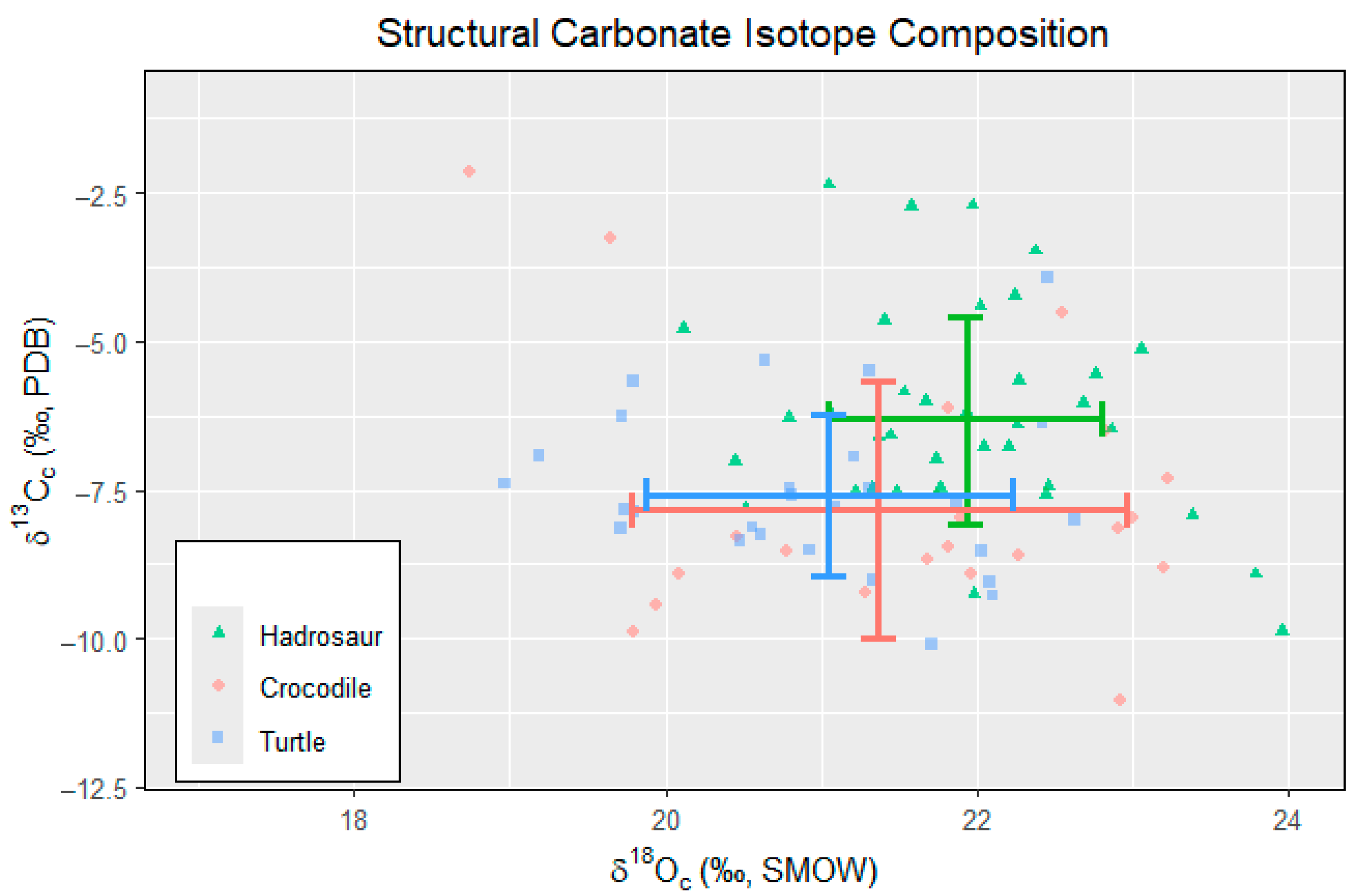
4.1. Carbon Isotope (δ13C)
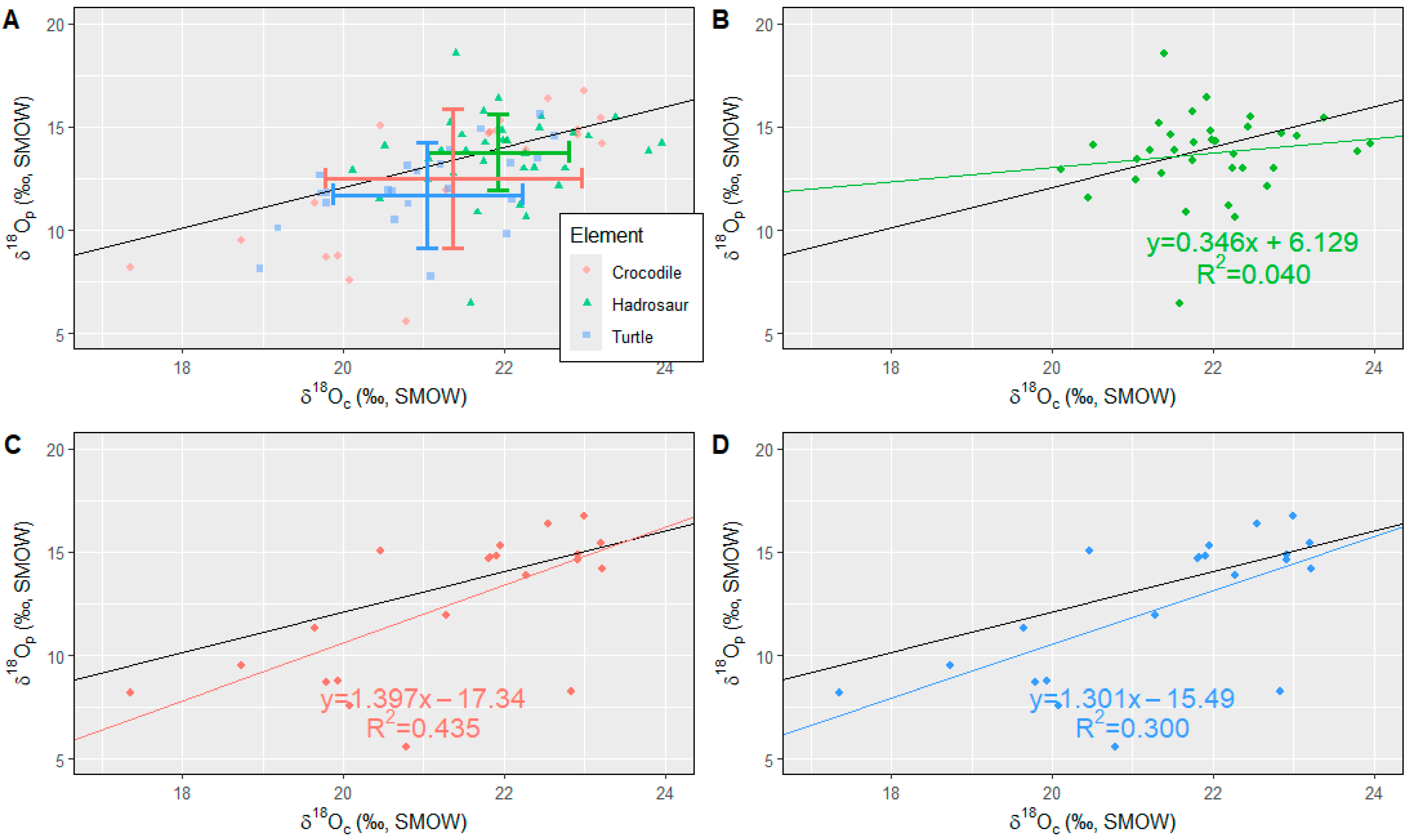
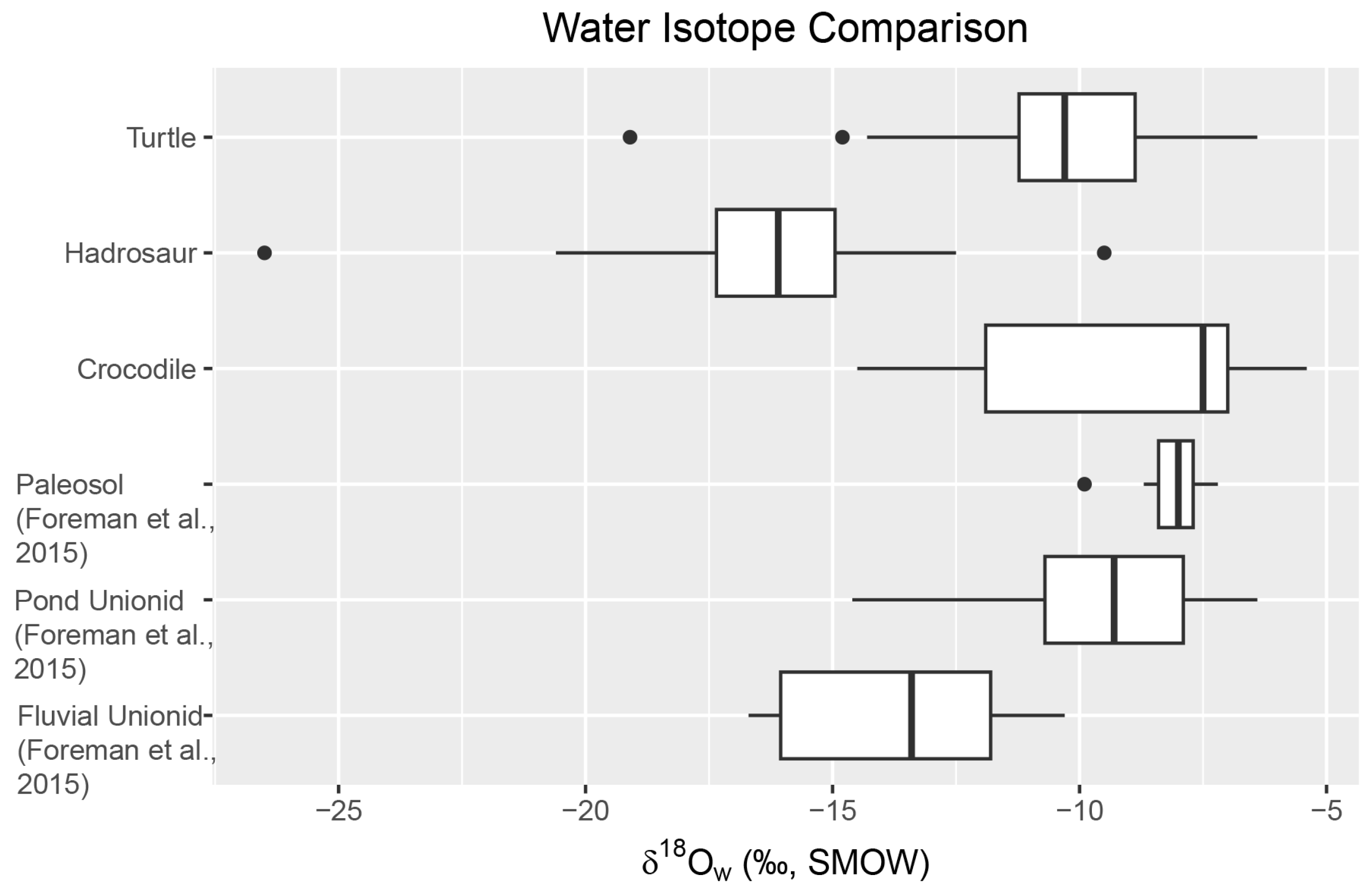
4.2. Oxygen Isotope (δ18Oc, δ18Op, and δ18Ow)
5. Discussion
5.1. Bone/Tooth Diagenesis
5.2. The δ13C as Diet Proxy
5.3. The Kaiparowits Paleoclimate
5.4. Campanian Paleoclimate Model
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| UMNH | Natural History Museum of Utah |
| UASIL | University of Arkansas Stable Isotope Laboratory |
| VPDB | Vienna Pee Dee Belemnite |
| VSMOW | Vienna Standard Mean Ocean Water |
References
- Kohn, M.J.; McKay, M.P.; Knight, J.L. Dining in the Pleistocene—Who’s on the Menu? Geology 2005, 33, 649–652. [Google Scholar] [CrossRef]
- Zazzo, A.; Balasse, M.; Passey, B.; Moloney, A.; Monahan, F.; Schmidt, O. The Isotope Record of Short-and Long-Term Dietary Changes in Sheep Tooth Enamel: Implications for Quantitative Reconstruction of Paleodiets. Geochim. Cosmochim. Acta 2010, 74, 3571–3586. [Google Scholar] [CrossRef]
- Barrick, R.E. Isotope Paleobiology of the Vertebrates: Ecology, Physiology, and Diagenesis. Paleontol. Soc. Pap. 1998, 4, 101–137. [Google Scholar] [CrossRef]
- Amiot, C.E.; Terry, D.J.; Jimmieson, N.L.; Callan, V.J. A Longitudinal Investigation of Coping Processes during a Merger: Implications for Job Satisfaction and Organizational Identification. J. Manag. 2006, 32, 552–574. [Google Scholar] [CrossRef]
- Fricke, H.C.; Foreman, B.Z.; Sewall, J.O. Integrated Climate Model-Oxygen Isotope Evidence for a North American Monsoon during the Late Cretaceous. Earth Planet. Sci. Lett. 2010, 289, 11–21. [Google Scholar] [CrossRef]
- Suarez, C.A.; Ludvigson, G.A.; Gonzalez, L.A.; Fiorillo, A.R.; Flaig, P.P.; McCarthy, P.J. Use of Multiple Oxygen Isotope Proxies for Elucidating Arctic Cretaceous Palaeo-Hydrology. Geol. Soc. 2013, 382, 185–202. [Google Scholar] [CrossRef]
- Kohn, M.J.; Schoeninger, M.J.; Valley, J.W. Herbivore Tooth Oxygen Isotope Compositions: Effects of Diet and Physiology. Geochim. Cosmochim. Acta 1996, 60, 3889–3896. [Google Scholar] [CrossRef]
- Kohn, M.J.; Cerling, T.E. Stable Isotope Compositions of Biological Apatite. Rev. Miner. Geochem. 2002, 48, 455–488. [Google Scholar] [CrossRef]
- Suarez, C.A.; You, H.-L.; Suarez, M.B.; Li, D.-Q.; Trieschmann, J. Stable Isotopes Reveal Rapid Enamel Elongation (Amelogenesis) Rates for the Early Cretaceous Iguanodontian Dinosaur Lanzhousaurus magnidens. Sci. Rep. 2017, 7, 15319. Available online: https://www.nature.com/articles/s41598-017-15653-6 (accessed on 30 April 2025). [CrossRef]
- Goedert, J.; Amiot, R.; Boudad, L.; Buffetaut, E.; Fourel, F.; Godefroit, P.; Kusuhashi, N.; Suteethorn, V.; Tong, H.; Watabe, M. Preliminary Investigation of Seasonal Patterns Recorded in the Oxygen Isotope Compositions of Theropod Dinosaur Tooth Enamel. Palaios 2016, 31, 10–19. [Google Scholar] [CrossRef]
- Yamamura, D.; Suarez, C.A.; Titus, A.L.; Manlove, H.M.; Jackson, T. Multiproxy Approaches to Investigating Palaeoecology and Palaeohydrology in the Upper Cretaceous Kaiparowits Formation, USA. Geol. Soc. 2021, 507, 293–311. [Google Scholar] [CrossRef]
- During, M.A.; Smit, J.; Voeten, D.F.; Berruyer, C.; Tafforeau, P.; Sanchez, S.; Stein, K.H.; Verdegaal-Warmerdam, S.J.; van der Lubbe, J.H. The Mesozoic Terminated in Boreal Spring. Nature 2022, 603, 91–94. Available online: https://www.nature.com/articles/s41586-022-04446-1 (accessed on 30 April 2025). [CrossRef] [PubMed]
- Dutton, A.; Carlson, A.E.; Long, A.J.; Milne, G.A.; Clark, P.U.; DeConto, R.; Horton, B.P.; Rahmstorf, S.; Raymo, M.E. Sea-Level Rise Due to Polar Ice-Sheet Mass Loss during Past Warm Periods. Science 2015, 349, aaa4019. [Google Scholar] [CrossRef]
- Dansgaard, W. Stable Isotopes in Precipitation. Tellus 1964, 16, 436–468. [Google Scholar] [CrossRef]
- Amiot, R.; Lécuyer, C.; Buffetaut, E.; Fluteau, F.; Legendre, S.; Martineau, F. Latitudinal Temperature Gradient during the Cretaceous Upper Campanian–Middle Maastrichtian: Δ18O Record of Continental Vertebrates. Earth Planet. Sci. Lett. 2004, 226, 255–272. [Google Scholar] [CrossRef]
- Hubert, J.; Panish, P.; Chure, D.; Prostak, K. Chemistry, Microstructure, Petrology, and Diagenetic Model of Jurassic Dinosaur Bones, Dinosaur National Monument, Utah. J. Sediment. Res. 1996, 66, 531–547. [Google Scholar] [CrossRef]
- Trueman, C.N.; Tuross, N. Trace Elements in Recent and Fossil Bone Apatite. Rev. Miner. Geochem. 2002, 48, 489–521. [Google Scholar] [CrossRef]
- Wings, O. Authigenic Minerals in Fossil Bones from the Mesozoic of England: Poor Correlation with Depositional Environments. Palaeogeogr. Palaeoclim. Palaeoecol. 2004, 204, 15–32. [Google Scholar] [CrossRef]
- Iacumin, P.; Bocherens, H.; Mariotti, A.; Longinelli, A. Oxygen Isotope Analyses of Co-Existing Carbonate and Phosphate in Biogenic Apatite: A Way to Monitor Diagenetic Alteration of Bone Phosphate? Earth Planet. Sci. Lett. 1996, 142, 1–6. [Google Scholar] [CrossRef]
- Zazzo, A.; Lécuyer, C.; Mariotti, A. Experimentally-Controlled Carbon and Oxygen Isotope Exchange between Bioapatites and Water under Inorganic and Microbially-Mediated Conditions. Geochim. Cosmochim. Acta 2004, 68, 1–12. [Google Scholar] [CrossRef]
- Piga, G.; Santos-Cubedo, A.; Solà, S.M.; Brunetti, A.; Malgosa, A.; Enzo, S. An X-Ray Diffraction (XRD) and X-Ray Fluorescence (XRF) Investigation in Human and Animal Fossil Bones from Holocene to Middle Triassic. J. Archaeol. Sci. 2009, 36, 1857–1868. [Google Scholar] [CrossRef]
- Fricke, H.C.; Pearson, D.A. Stable Isotope Evidence for Changes in Dietary Niche Partitioning among Hadrosaurian and Ceratopsian Dinosaurs of the Hell Creek Formation, North Dakota. Paleobiology 2008, 34, 534–552. [Google Scholar] [CrossRef]
- Fricke, H.C.; Rogers, R.R.; Gates, T.A. Hadrosaurid Migration: Inferences Based on Stable Isotope Comparisons among Late Cretaceous Dinosaur Localities. Paleobiology 2009, 35, 270–288. [Google Scholar] [CrossRef]
- Foreman, B.Z.; Roberts, E.M.; Tapanila, L.; Ratigan, D.; Sullivan, P. Stable Isotopic Insights into Paleoclimatic Conditions and Alluvial Depositional Processes in the Kaiparowits Formation (Campanian, South-Central Utah, USA). Cretac. Res. 2015, 56, 180–192. [Google Scholar] [CrossRef]
- Crystal, V.F.; Evans, E.S.; Fricke, H.; Miller, I.M.; Sertich, J.J. Late Cretaceous Fluvial Hydrology and Dinosaur Behavior in Southern Utah, USA: Insights from Stable Isotopes of Biogenic Carbonate. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 516, 152–165. [Google Scholar] [CrossRef]
- Burgener, L.; Hyland, E.; Huntington, K.W.; Kelson, J.R.; Sewall, J.O. Revisiting the Equable Climate Problem during the Late Cretaceous Greenhouse Using Paleosol Carbonate Clumped Isotope Temperatures from the Campanian of the Western Interior Basin, USA. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 516, 244–267. [Google Scholar] [CrossRef]
- Roberts, E.M. Facies Architecture and Depositional Environments of the Upper Cretaceous Kaiparowits Formation, Southern Utah. Sediment. Geol. 2007, 197, 207–233. [Google Scholar] [CrossRef]
- Roberts, E.M.; Sampson, S.D.; Deino, A.L.; Bowring, S.A.; Buchwaldt, R. The Kaiparowits Formation: A Remarkable Record of Late Cretaceous Terrestrial Environments, Ecosystems, and Evolution in Western North America; Indiana University Press: Bloomington, Indiana, 2013; ISBN 0253008832. [Google Scholar]
- Beveridge, T.L.; Roberts, E.M.; Titus, A.L. Volcaniclastic Member of the Richly Fossiliferous Kaiparowits Formation Reveals New Insights for Regional Correlation and Tectonics in Southern Utah during the Latest Campanian. Cretac. Res. 2020, 114, 104527. [Google Scholar] [CrossRef]
- Titus, A.L.; Powell, J.D.; Roberts, E.M.; Sampson, S.D.; Pollock, S.L.; Kirkland, J.I.; Albright, L.B. Late Cretaceous Stratigraphy, Depositional Environments, and Macrovertebrate Paleontology of the Kaiparowits Plateau, Grand Staircase–Escalante National Monument, Utah. Field Guides 2005, 6, 101–128. [Google Scholar] [CrossRef]
- Titus, A.L.; Roberts, E.M.; Albright, L.B., III. Geologic Overview. In At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah; Indiana University Press: Bloomington, Indiana, 2013; pp. 13–41. [Google Scholar]
- Goldstrand, P.M.; Trexler, J.; Kowallis, B.J.; Eaton, J.G.; Morales, M. Late Cretaceous to Early Tertiary Tectonostratigraphy of Southwestern Utah. In Aspects of Mesozoic Geology and Paleontology of the Colorado Plateau: Flagstaff, Arizona, Museum of Northern Arizona, Bulletin; Museum of Northern Arizona: Flagstaff, AZ, USA, 1993; Volume 59, pp. 181–188. [Google Scholar]
- Hutchison, J.H.; Knell, M.J.; Brinkman, D.B.; Titus, A.; Loewen, M. Turtles from the Kaiparowits Formation, Utah. In At the Top of the Grand Staircase, The Late Cretaceous of Southern Utah; Indiana University Press: Bloomington, Indiana, 2013; pp. 295–318. [Google Scholar]
- Irmis, R.B.; Hutchison, J.H.; Sertich, J.J.; Titus, A.L. Crocodyliforms from the Late Cretaceous of Grand Staircase-Escalante National Monument and Vicinity, Southern Utah, USA. In At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah; Indiana University Press: Bloomington, Indiana, 2013; pp. 424–444. [Google Scholar]
- Sampson, S.D.; Loewen, M.A.; Roberts, E.M.; Getty, M.A. A New Macrovertebrate Assemblage from the Late Cretaceous (Campanian) of Southern Utah; Indiana University Press: Bloomington, Indiana, 2013; ISBN 0253008832. [Google Scholar]
- Sampson, S.D.; Lund, E.K.; Loewen, M.A.; Farke, A.A.; Clayton, K.E. A Remarkable Short-Snouted Horned Dinosaur from the Late Cretaceous (Late Campanian) of Southern Laramidia. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131186. [Google Scholar] [CrossRef]
- Miller, I.; Johnson, K.; Kline, D.; Nichols, D.; Barclay, R. A Late Campanian Flora from the Kaiparowits Formation, Southern Utah, and a Brief Overview of the Widely Sampled but Little-Known Campanian Vegetation of the Western Interior of North America. In At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah; Indiana University Press: Bloomington, Indiana, 2013; pp. 107–131. [Google Scholar]
- Roberts, E.M.; Chan, M.A. Variations in Iron Oxide, Iron Sulfide, and Carbonate Concretions and Their Distributions in Fluvio-Deltaic and Nearshore Sandstones: Cretaceous Examples from the Kaiparowits Plateau, Utah and San Juan Basin, New Mexico; Utah Geological Association: Salt Lake City, UT, USA, 2010. [Google Scholar]
- Koch, P.L.; Tuross, N.; Fogel, M.L. The Effects of Sample Treatment and Diagenesis on the Isotopic Integrity of Carbonate in Biogenic Hydroxylapatite. J. Archaeol. Sci. 1997, 24, 417–429. [Google Scholar] [CrossRef]
- Vennemann, T.W.; Fricke, H.C.; Blake, R.E.; O’Neil, J.R.; Colman, A. Oxygen Isotope Analysis of Phosphates: A Comparison of Techniques for Analysis of Ag3PO4. Chem. Geol. 2002, 185, 321–336. [Google Scholar] [CrossRef]
- Suarez, C.A.; González, L.A.; Ludvigson, G.A.; Cifelli, R.L.; Tremain, E. Water Utilization of the Cretaceous Mussentuchit Member Local Vertebrate Fauna, Cedar Mountain Formation, Utah, USA: Using Oxygen Isotopic Composition of Phosphate. Palaeogeogr. Palaeoclim. Palaeoecol. 2012, 313, 78–92. [Google Scholar] [CrossRef]
- Amiot, R.; Lécuyer, C.; Escarguel, G.; Billon-Bruyat, J.-P.; Buffetaut, E.; Langlois, C.; Martin, S.; Martineau, F.; Mazin, J.-M. Oxygen Isotope Fractionation between Crocodilian Phosphate and Water. Palaeogeogr. Palaeoclim. Palaeoecol. 2007, 243, 412–420. [Google Scholar] [CrossRef]
- Coulson, A.B.; Kohn, M.J.; Shirley, M.H.; Joyce, W.G.; Barrick, R.E. Phosphate–Oxygen Isotopes from Marine Turtle Bones: Ecologic and Paleoclimatic Applications. Palaeogeogr. Palaeoclim. Palaeoecol. 2008, 264, 78–84. [Google Scholar] [CrossRef]
- Lécuyer, C.; Bogey, C.; Garcia, J.-P.; Grandjean, P.; Barrat, J.-A.; Floquet, M.; Bardet, N.; Pereda-Superbiola, X. Stable Isotope Composition and Rare Earth Element Content of Vertebrate Remains from the Late Cretaceous of Northern Spain (Laño): Did the Environmental Record Survive? Palaeogeogr. Palaeoclim. Palaeoecol. 2003, 193, 457–471. [Google Scholar] [CrossRef]
- Lohmann, K.C. Geochemical Patterns of Meteoric Diagenetic Systems and Their Application to Studies of Paleokarst. In Paleokarst; Springer: Berlin/Heidelberg, Germany, 1988; pp. 58–80. [Google Scholar]
- Wang, Y.; Cerling, T.E. A Model of Fossil Tooth and Bone Diagenesis: Implications for Paleodiet Reconstruction from Stable Isotopes. Palaeogeogr. Palaeoclim. Palaeoecol. 1994, 107, 281–289. [Google Scholar] [CrossRef]
- Eagle, R.A.; Tütken, T.; Martin, T.S.; Tripati, A.K.; Fricke, H.C.; Connely, M.; Cifelli, R.L.; Eiler, J.M. Dinosaur Body Temperatures Determined from Isotopic (13C-18O) Ordering in Fossil Biominerals. Science 2011, 333, 443–445. [Google Scholar] [CrossRef]
- Barrick, R.E.; Fischer, A.G.; Showers, W.J. Oxygen Isotopes from Turtle Bone: Applications for Terrestrial Paleoclimates? Palaios 1999, 14, 186–191. [Google Scholar] [CrossRef]
- Barrick, R.E.; Showers, W.J. Thermophysiology of Tyrannosaurus Rex: Evidence from Oxygen Isotopes. Science 1994, 265, 222–224. [Google Scholar] [CrossRef]
- Barrick, R.E.; Showers, W.J. Oxygen Isotope Variability in Juvenile Dinosaurs (Hypacrosaurus): Evidence for Thermoregulation. Paleobiology 1995, 21, 552–560. [Google Scholar] [CrossRef]
- Bocherens, H. Preservation of Isotopic Signals (13C, 15N) _in Pleistocene Mammals. In Biogeochemical Approaches to Paleodietary Analysis; Springer: Berlin/Heidelberg, Germany, 2000; pp. 65–88. [Google Scholar]
- Cullen, T.M.; Longstaffe, F.J.; Wortmann, U.G.; Huang, L.; Fanti, F.; Goodwin, M.B.; Ryan, M.J.; Evans, D.C. Large-Scale Stable Isotope Characterization of a Late Cretaceous Dinosaur-Dominated Ecosystem. Geology 2020, 48, 546–551. [Google Scholar] [CrossRef]
- Sewall, J.O.; Fricke, H.C. Andean-Scale Highlands in the Late Cretaceous Cordillera of the North American Western Margin. Earth Planet. Sci. Lett. 2013, 362, 88–98. [Google Scholar] [CrossRef]
- Jinnah, Z.A.; Roberts, E.M.; Deino, A.L.; Larsen, J.S.; Link, P.K.; Fanning, C.M. New 40Ar-39Ar and Detrital Zircon U-Pb Ages for the Upper Cretaceous Wahweap and Kaiparowits Formations on the Kaiparowits Plateau, Utah: Implications for Regional Correlation, Provenance, and Biostratigraphy. Cretac. Res. 2009, 30, 287–299. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| δ13C | δ18Oc | δ18Op | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Element | Mean | Range | SD | σ2 | Mean | Range | SD | σ2 | Mean | Range | SD | σ2 |
| Hadrosaur teeth (n = 36) | −6.32 | −9.25 to −2.35 | 1.752 | 3.070 | 21.9 | 20.1 to 24.0 | 0.872 | 0.760 | 13.8 | 6.4 to 15.3 | 1.852 | 3.430 |
| Crocodile teeth (n = 22) | −7.84 | −11.03 to −2.15 | 2.156 | 4.648 | 21.4 | 17.4 to 23.2 | 1.589 | 2.525 | 12.5 | 5.6 to 16.8 | 3.365 | 11.323 |
| Turtle shell (n = 28) | −7.58 | −10.10 to −3.93 | 1.354 | 1.832 | 21.0 | 19.0 to 24.1 | 1.178 | 1.387 | 11.7 | 3.8 to 15.6 | 2.550 | 6.501 |
| δ13C | δ18Oc | δ18Op | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diff. | p-Value | Significance | Diff. | p-Value | Significance | Diff. | p-Value | Significance | Test | |
| Hadrosaur teeth vs. Crocodile teeth | 1.57 | 0.0007 | very strong | 0.55 | 0.4195 | no | 1.27 | 0.672 | no | Wilcoxon |
| Hadrosaur teeth vs. Turtle shell | 1.26 | 0.0017 | very strong | 0.90 | 0.00110 | very strong | 2.10 | 0.00188 | very strong | t-test |
| Crocodile vs. Turtle | 0.26 | 0.140 | no | 0.40 | 0.229 | no | 0.8 | 0.228 | no | Wilcoxon |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamura, D.; Suarez, C. Vertebrate Skeletal Remains as Paleohydrologic Proxies: Complex Hydrologic Setting in the Upper Cretaceous Kaiparowits Formation. Geosciences 2025, 15, 262. https://doi.org/10.3390/geosciences15070262
Yamamura D, Suarez C. Vertebrate Skeletal Remains as Paleohydrologic Proxies: Complex Hydrologic Setting in the Upper Cretaceous Kaiparowits Formation. Geosciences. 2025; 15(7):262. https://doi.org/10.3390/geosciences15070262
Chicago/Turabian StyleYamamura, Daigo, and Celina Suarez. 2025. "Vertebrate Skeletal Remains as Paleohydrologic Proxies: Complex Hydrologic Setting in the Upper Cretaceous Kaiparowits Formation" Geosciences 15, no. 7: 262. https://doi.org/10.3390/geosciences15070262
APA StyleYamamura, D., & Suarez, C. (2025). Vertebrate Skeletal Remains as Paleohydrologic Proxies: Complex Hydrologic Setting in the Upper Cretaceous Kaiparowits Formation. Geosciences, 15(7), 262. https://doi.org/10.3390/geosciences15070262