Abstract
Carentonosaurus soaresi was recently described in the uppermost middle Cenomanian (Upper Cretaceous) of Casais dos Carecos (Coimbra, western Portugal) based on a diverse set of new material (cervical and dorsal vertebrae) of the Pythonomorpha lizard. The main morphological characteristics observed in the vertebrae used for the diagnosis of this species are the presence of distinct lateral and subcentral foramina, highly laterally projected paradiapophyses beyond the prezygapophyses, a low subrectangular neural spine ornamented with longitudinal grooves, and dorsal vertebrae displaying a sagittal furrow along the ventral surface. Additional diagnostic details observed both in new material and the previously studied vertebrae are described herein, such as the presence of keels in the zygantrum and zygosphene. These and other important morphological characteristics present in the species soaresi are absent in the genotype Carentonosaurus mineaui and in other known Squamata, allowing for the definition of the new genus Segurasaurus.
1. Introduction
Recently, the first record of the marine lizard Carentonosaurus Rage and Néraudeau, 2004 [1] has been reported from the Cenomanian of Portugal. The type material consists of 10 vertebrae (cervical and dorsal vertebrae), whose features have allowed the description of the new species Carentonosaurus soaresi Pimentel, Berrocal-Casero, Audije-Gil and Callapez, 2023 [2]. Nevertheless, this taxon has significant morphological features that allow its differentiation from Carentonosaurus mineaui Rage and Néraudeau, 2004, the type species of this genus [1].
Before this new record, the genus Carentonosaurus was only known from the Cenomanian of France, e.g., [1,3,4,5] and Spain [6], and, additionally, as reworked bones from the Pleistocene [7,8]. After the submission of the work related to C. soaresi, a proposed species named Carentonosaurus algorensis Cabezuelo-Hérnandez and Pérez-García, 2023, has also been described with a single vertebra from the Cenomanian of Algora (central Spain) [9], showing some features close to those of C. soaresi.
As presented by [1], the diagnostic features of the genus Carentonosaurus are the same as the type species C. mineaui, as this genus was originally regarded as monospecific. These morphological features are not compatible with those of C. soaresi at a “generic level”, suggesting that a new genus should be defined to separate these Portuguese findings from the original French taxon.
The aims of this paper are thus to compare the main characteristics of C. soaresi with the original diagnostic features of the genus Carentonosaurus, in order to determine the taxonomic basis needed to create a new genus.
2. Location and Stratigraphic and Palaeoenvironmental Contexts
The type material of Carentonosaurus soaresi Pimentel, Berrocal-Casero, Audije-Gil and Callapez, 2023 [2] was sampled in the quarry of Beiraterra (40°15′10.9″ N, 8°34′13.3″ W) located close to the village of Casais dos Carecos in the Baixo Mondego ranges (Coimbra, western Portugal) (Figure 1A,B). It consists of 10 vertebrae found in situ [2] in the lower part of the Tentúgal Formation [10]. This unit locally records the West Portuguese carbonate platform [11]. The lower part of the succession (“Unit B”) [12] corresponds to the uppermost middle Cenomanian and consists of sandstones intercalated with thin layers of sandy limestones, mudstones, and marls with invertebrate assemblages containing oysters (Gyrostrea ouremensis), mussels (Septifer lineatus), and infaunal bivalves (Anisocardia orientalis) [13,14,15]. This shallow-water, tidal-flat sequence with mixed facies, rich in vertebrate remains, is overlaid by upper Cenomanian shelf limestones with a cephalopod assemblage dominated by the Tethyan ammonite species Neolobites vibrayeanus [12,13]. This sequence of nodular marly limestones and a massive limestone bed (“Unit C” and “Unit D”) [12] finishes upwards with several meters of nodular marly limestones and calcareous marls with Vascoceras ammonites (e.g., [13,14,15,16,17,18,19,20,21]) (Figure 1C).
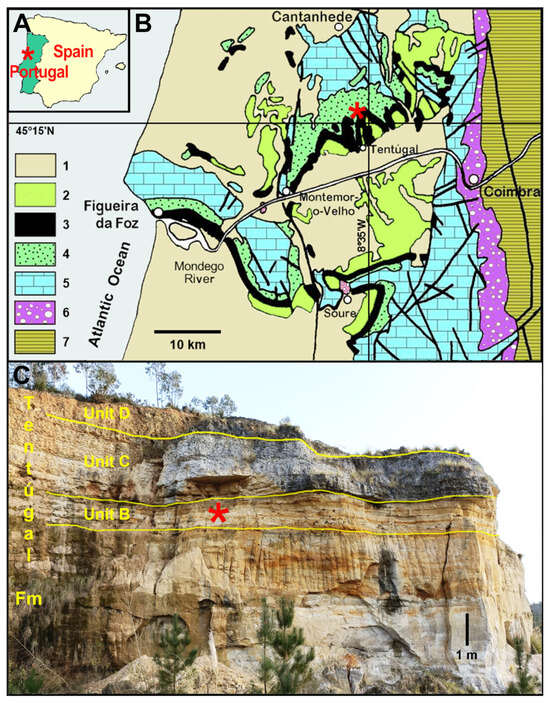
Figure 1.
(A) Location of Casais dos Carecos (Coimbra, western Portugal) within the context of the Iberian Peninsula. (B) Simplified geological map of the Baixo Mondego region, showing the West Portuguese Margin units, and the geographic position of the type locality. 1: Cenozoic; 2: Upper Cretaceous (lower Turonian to Maastrichtian) coarse alluvial siliciclastics and reddish lutites; 3: Upper Cretaceous (middle Cenomanian to lower Turonian) platform carbonates, including the Tentúgal Formation; 4: Lower to Upper Cretaceous (upper Aptian to middle Cenomanian) alluvial to marginal marine coarse siliciclastics; 5: Jurassic (except Hettangian) mostly marine carbonates; 6: Triassic and Hettangian red-beds and evaporites; 7: Pre-Cambrian and Palaeozoic Variscan basement. (C) Panoramic view of the Beiraterra quarry, type locality of Segurasaurus gen. nov., showing the middle to basal upper Cenomanian succession with the carbonate platform stratigraphic units B, C, and D of the Tentúgal Formation individualized marginal marine coarse sandstones. The red asterisks indicate the location of Casais dos Carecos and the levels that have provided the studied material.
For the local palaeogeography, the incidence of a rather diverse vertebrate fauna [2,22] found in mixed and carbonate facies with shallow marine invertebrate taxa can be interpreted as the local record of the inner shelf to tidal flat environments of a carbonate platform, which was mostly developed in a transgressive context, with retrogradation of extensive post-rift alluvial systems located eastward of the Baixo Mondego ranges, e.g., [11,15,16,17,20,21]. For more information on the general geological, stratigraphic, and palaeoenvironmental settings of the Casais dos Carecos fossil site and additional references, see [2].
3. Material and Methods
The additional vertebra studied for this work was obtained from the screen-washing of a 200 kg bulk sample of sediments. The vertebra was studied with a Leica© MZ 16A binocular stereomicroscope and photographs were taken with an Olympus© E-M5II camera. Measurements were taken using a digital caliper. The vertebra measurements (Table 1) and the anatomical abbreviations mainly follow [1,2,8,23]. The discussed specimens (DCT-CC-1, DCT-CC-2, DCT-CC-4, DCT-CC-5, DCT-CC-7, DCT-CC-8, DCT-CC-27, DCT-CC-28, DCT-CC-29, and DCT-CC-31; see Figures 5–7 in [2] and new material (Figure 2B), are part of the palaeontological collection of Casais dos Carecos housed at the Department of Earth Sciences (DCT-CC), University of Coimbra, Portugal.

Table 1.
Available measurements on the vertebra DCT-CC-35 of Segurasaurus soaresi from the uppermost middle Cenomanian of Casais dos Carecos, Coimbra (western Portugal). Measurements in mm.

Figure 2.
Segurasaurus soaresi (Pimentel, Berrocal-Casero, Audije-Gil and Callapez) [2] from the uppermost middle Cenomanian of Casais dos Carecos, Coimbra (western Portugal). (A) Dorsal vertebra DCT-CC-1, holotype of the genotype of Segurasaurus gen. nov. (B) Dorsal vertebra DCT-CC-35, new material. The numbers indicate: cranial (1); caudal (2); right lateral (3); left lateral (4); dorsal (5); and ventral (6) views. Scale bar = 1 cm.
3.1. Vertebrae Measurement Abbreviations
Centrum length, measured ventrally: cL; condyle height: cdH; condyle width: cdW; cotyle height: ctH; cotyle width: ctW; distance between the exterior edges of the postzygapophyses: ptzD; distance between the exterior edges of the prezygapophyses: przD; distance between the paradiapophyses: pdD; maximum height in lateral view: MH; maximum length measured ventrally: ML; neural arch width, measured at the maximum interzygapophyseal constriction: naW; neural spine length: nsL; zygosphene width: zsW.
3.2. Vertebrae Anatomical Abbreviations
Condyle: cd; cotyle: ct; epidiapophyseal ridge: er; interzygapophyseal ridge: ir; lateral foramina: lf; longitudinal grooves: lg; neural canal: nc; neural spine: ns; paracotylar foramen: pcf; paradiapophysis: pd; parazygosphenal foramen: pzf; postzygapophysis: ptz; prezygapophyseal facet: prf; prezygapophysis: prz; sagittal furrow: sf; subcentral foramen: sbf; zygantral foramina: zf; zygantral keel: ztk; zygantrum: zt; zygosphenal keel: zsk; zygosphene: zs.
4. Systematic Palaeontology (by Mélani Berrocal-Casero, Ricardo Pimentel, and Pedro M. Callapez)
Class: Reptilia Linnaeus, 1758 [24]
Order: Squamata Oppel, 1811 [25]
Unranked Pythonomorpha Cope, 1869 [26]
Genus: Segurasaurus gen. nov.
Type species: Carentonosaurus soaresi Pimentel, Berrocal-Casero, Audije-Gil and Callapez, 2023, in Figures 5–7 in [2].
Type material: Holotype DCT-CC-1 (Figure 2A); paratypes: DCT-CC-2, DCT-CC-4, DCT-CC-5, DCT-CC-7, DCT-CC-8, DCT-CC-27, DCT-CC-28, DCT-CC-29, and DCT-CC-31 (see Figures 5–7 in [2]). New material: DCT-CC-35 (Figure 2B).
4.1. Diagnosis
The diagnosis of Segurasaurus is that of the type species S. soaresi [2], considering cervical and dorsal vertebrae until more anatomical material and additional species are known. Segurasaurus has pachyostotic mid-dorsal vertebrae, being the cervical and the most posterior dorsal vertebrae non or weakly pachyostotic.
The cervical and dorsal vertebrae of S. soaresi differ from those of Carentonosaurus mineaui [1], the type species of the genus Carentonosaurus, in having the following features: the paradiapophyses are highly laterally projected beyond the prezygapophyses; the epidiapophyseal and interzygapophyseal ridges are poorly defined and usually merged; and the neural spine is ornamented with longitudinal grooves. In addition, Zygosphenal and zygantral vertical keels (less conspicuous in cervical vertebrae) and lateral and subcentral foramina are present.
The dorsal vertebrae of S. soaresi differ from those of C. mineaui in the following features: zygantrum with articular facets; a zygosphene that is deeply notched, highly projected, exceeding the length of the centrum, being as long as the prezygapophyses in dorsal and ventral views; a low subrectangular neural spine ornamented with longitudinal grooves; the cotyle-condyle system is less oblique, being sometimes straight; the lateral borders of the centrum are concave in ventral view; and the presence of sagittal furrow on the ventral surface.
4.2. Distribution
The samples correspond to the uppermost middle Cenomanian, at the lower part of the Tentúgal Formation (Type horizon: “Unit B” with Gyrostrea ouremensis), just below the first nodular limestone beds, which presents a cephalopod assemblage with Neolobites vibrayeanus and Calycoceras naviculare, correlative to the standard Zone of Calycoceras guerangeri (considered the lowermost upper Cenomanian ammonite biozone).
4.3. Type Locality
Beiraterra quarry, Casais dos Carecos, Baixo Mondego ranges (Coimbra, western Portugal; coordinates: 40°15′07.0″ N 8°34′15.6″ W).
4.4. Etymology
Named after Emeritus Professor Manuel Segura, former director of the Department of Geology of the University of Alcalá (Alcalá de Henares, Spain) and renowned specialist on the Iberian Cretaceous.
4.5. Material Description
The studied material is composed of ten procoelous vertebrae, with diverse dimensions previously classified as Carentonosaurus soaresi in [2]. It also includes a new additional dorsal vertebra, DCT-CC-35 (Figure 2B and Table 1).
Description of the New Material (DCT-CC-35)
In anterior view, the vertebra DCT-CC-35 is strongly depressed and wide, with it showing a subrectangular shape (Figure 2(B1)). The neural canal is sub-triangular in shape and higher than it is wide (Figure 2(B1,B2)). The cotyle is concave, subcircular, and as wide as the zygosphene (Figure 2(B1)). The vertebra presents parazygosphenal and paracotylar foramina (Figure 2(B1,B2,B5)). A keel can be well differentiated in the middle of the zygantrum and the zygosphene, connecting with the base of the neural spine (Figure 2(B1,B2)). The zygantrum is formed by two fossae located below the neural spine. In posterior view, the right zygantral foramen and the zygantral facets are visible. The condyle is convex, oval, and slightly wider than it is high (Figure 2(B2)). In lateral view, the condyle exceeds the posterior margin of the neural spine, and the postzygapophyses are larger than the neural spine (Figure 2(B3,B4)).
The neural spine is partially broken but ranges across the entire length of the neural arch, although its posterior end is longer in the upper part and shorter close to the base (Figure 2(B3,B4)). In lateral view, the sub-rectangular and low profile of the neural spine is evident, with it being considerably longer than it is high and ornamented with longitudinal grooves. The epidiapophyseal and interzygapophyseal ridges are perceptible and seem to be fused together. The axis of the cotyle-condyle system is almost straight (Figure 2(B3,B4)).
The width of the prezygapophyses exceeds that of the postzygapophyses, and the paradiapophyses are highly projected laterally beyond the prezygapophyses (Figure 2(B5)). They are located anteriorly in the vertebra, behind the prezygapophyses. In dorsal view, the prezygapophyseal facets are large, and the centrum displays a marked posterior constriction of the neural arch (Figure 2(B5)). The zygosphene is well developed and deeply notched, with it being highly projected and exceeding the length of the centrum and being as long as the prezygapophyses in dorsal and ventral views (Figure 2(B5,B6)). The anterior margins of the zygosphenal articular facets are rounded and they are laterally delimited by concave borders (Figure 2(B5)). In ventral view, the centrum shows a subtriangular shape, with concave lateral borders, and the left subcentral foramen can be seen. A sagittal furrow is also evident, delimited by blunt ridges (Figure 2(B6)). The ventral edges of the paradiapophyses are prominent (Figure 2(B6)). The postzygapophyseal facets are comparatively big and subrounded (Figure 2(B6)).
5. Discussion
5.1. Taxonomic Remarks
According to the diagnosis of the genus Carentonosaurus Rage and Néraudeau, 2004, which is the same as the type species Carentonosaurus mineaui Rage and Néraudeau, 2004 [1], this Pythonomorpha lizard is characterized by the association of pachyostotic mid- and posterior dorsal vertebrae and non-pachyostotic cervical, sacral, and caudal vertebrae. After [1], each vertebra is characterized by a combination of the following characteristics: “(1) presence of a zygosphene with well-shaped articular facets, and a zygantrum without articular facets; (2) the anterior border of the zygosphene is concave but not deeply notched; (3) frequent presence of paracotylar, parazygosphenal, and zygantral foramina, but absence of lateral and subcentral foramina; (4) the prezygapophyseal facets are somewhat larger than the postzygapophyseal facets; consequently, the vertebrae are wider across the prezygapophyses (paradiapophyses excluded) than across the postzygapophyses; (5) the precondylar constriction is absent. Moreover, the posterior cervical and dorsal vertebrae are very depressed. The axis of the cotyle-condyle system is weakly oblique, except in anterior cervical, sacral, and caudal vertebrae. Excluding the anterior cervical vertebrae, the paradiapophyses and epidiapophyseal ridges project strongly laterally and the interzygapophyseal ridges are approximately parallel to the vertebral axis. Lastly, the rib is weakly curved with a reduced pseudotuberculum”.
Considering the above-mentioned description of the genus Carentonosaurus, and the fact that it was based on the same material and characteristics used to define the type species C. mineaui, the substantial diagnostic differences observed in the dorsal vertebrae of C. soaresi (Figure 3A,B) suggest that this recently described new species can be considered as the type of a new genus. The previously mentioned main diagnostic characteristics of Carentonosaurus are different than those of Segurasaurus: (1) Carentonosaurus shows a zygantrum without articular facets, while Segurasaurus shows a zygantrum with articular facets; (2) the zygosphene is deeply notched in Segurasaurus but not in Carentonosaurus; and (3) the lateral and subcentral foramina are present in Segurasaurus, but not in Carentonosaurus. In this sense, three of five of the diagnostic features of the genus Carentonosaurus proposed by Rage and Néraudeau [1] are not present in of the specimens from Portugal.
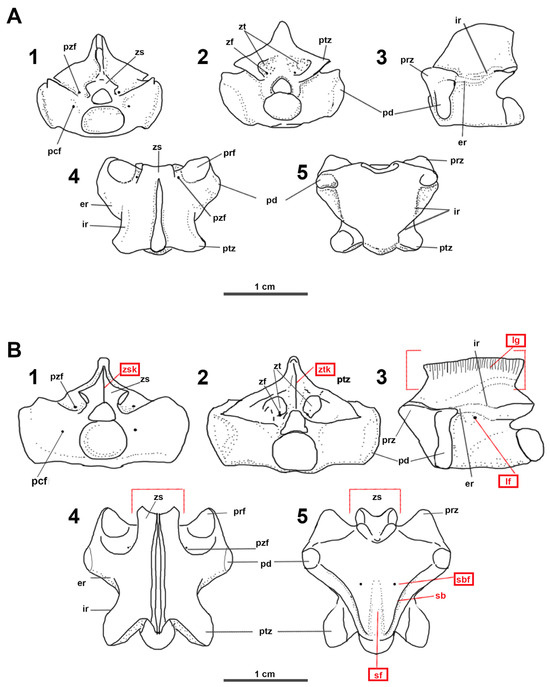
Figure 3.
Drawings showing the morphological features described in the text. (A) Carentonosaurus mineaui Rage and Néraudeau, 2004 [1] from the Cenomanian of Madame Island, France (modified from p. 1199, Figure 2 in [1]). (B) Segurasaurus soaresi (Pimentel, Berrocal-Casero, Audije-Gil and Callapez, 2023) [2] from the uppermost middle Cenomanian of Casais dos Carecos, Coimbra (Portugal). Cranial (1); caudal (2); lateral (3); dorsal (4); and ventral (5) views. Scale bars = 1 cm. The red letters and rectangles highlight the features present in Segurasaurus gen. nov. but are absent in Carentonosaurus Rage and Néraudeau, 2004 [1].
Moreover, Segurasaurus, the herein-proposed genus, differs from Carentonosaurus in the following features: the first presents seemly more depressed dorsal vertebrae (Figure 3(B1–B3) vs. Figure 3(A1–A3)); the existence of zygosphenal and zygantral vertical keels (Figure 3(B1,B2) vs. Figure 3(A1,A2); the presence of lateral and subcentral foramina, in addition to the paracotylar, parazygosphenal, and zygantral ones (Figure 3(B1–B4) vs. Figure 3(A1–A4)); and a low neural spine, which is also subrectangular and ornamented with longitudinal grooves (Figure 3(B3) vs. Figure 3(A3)). The interzygapophyseal and epidiapophyseal ridges are also different in Segurasaurus, with them being less evident and mainly merged compared to those from Carentonosaurus (Figure 3(B3) vs. Figure 3(A3)). The cotyle-condyle system is less oblique in Segurasaurus, with it being sometimes straight (Figure 3(B3) vs. Figure 3(A3)).
In dorsal view, the paradiapophyses are projected laterally beyond the prezygapophyses in both vertebrae; however, these structures, the prezygapophyses and the postzygapophyses, are longer and much more laterally projected (Figure 3(B4) vs. Figure 3(A4)); the zygosphene is deeply notched in Segurasaurus, with it being highly projected, exceeding the length of the centrum, and being as long as the prezygapophyses in dorsal and ventral views (Figure 3(B4,B5) vs. Figure 3(A4,A5)).
In ventral view, both taxa are defined as presenting a “subtriangular shape”; nonetheless, these shapes are notably different from one another, with them showing lateral concave borders in Segurasaurus. Moreover, the ventral surface of the dorsal vertebrae of Segurasaurus presents a sagittal furrow (Figure 3(B5) vs. Figure 3(A5)).
Considering the cervical vertebrae of Segurasaurus, some diagnostic characteristics are present in both dorsal and cervical vertebrae: the prezygapophyses exceed those of the postzygapophyses; the paradiapophyses are highly laterally projected beyond the prezygapophyses; and the epidiapophyseal and interzygapophyseal ridges are poorly defined and usually merged. Although the neural spine in cervical vertebrae is taller posteriorly, the ornamentation with longitudinal grooves is also present (Figure 6(A3,A4) in [2]). A zygantral vertical narrow keel and the paracotylar and zygantral foramina are also distinguished in cervical vertebrae (Figure 6(A2–B2) in [2]). Moreover, lateral and subcentral foramina can also be differentiated in Figure 6(B4,B6) in [2].
Comparing the cervical vertebrae of Carentonosaurus and Segurasaurus, those attributed herein to Segurasaurus show longitudinal grooves on the neural spine, while it is smooth in Carentonosaurus (Figure 3F–H in [1]). In the cervical vertebrae of Segurasaurus, a vertical keel is distinguished in the zygantrum, with it being absent in Carentonosaurus [1].
Following the discussion set above, Segurasaurus gen. nov. is proposed herein and includes the species S. soaresi.
An emended diagnosis of the genus Carentonosaurus [9], considering the common features of the dorsal vertebra found in Algora (central Spain) and the dorsal vertebrae of C. mineaui from Madame Island (western France), describes: “neural arch characteristically wider anteriorly (across the prezygapophyses, the paradiapophyses being excluded) than posteriorly (across the postzygapophyses); paradiapophyses strongly extending laterally beyond the prezygapophyses; width across the paradiapophyses notably exceeding that across the postzygapophyses (in dorsal view); extreme pachyosteosclerotic neural arch, with a non-pachyostotic neural spine; prezygapophyseal facets larger than the postzygapophyseal facets; prezygapophyses standing out weakly against the bulk of the paradiapophyses; the condyle clearly exceeding the posterior margin of the neural spine (in lateral view); interzygapophyseal constriction located further back from the mid-length of the neural arch; small, subtriangular neural canal; cotyle wider than the zygosphene; and ‘V’-shaped centrum (in ventral view)”. This emended diagnosis also supports the proposal that the specimens from Portugal cannot be included in the genus Carentonoaurus as some features are not common in the Cenomanian specimens from the Iberian Peninsula, or at least in the specimens from Portugal. For example, the paradiapophyses are much more laterally projected beyond the prezygapophyses in the Cenomanian specimen from Portugal in comparison with Careontonosaurus mineaui (Figure 3(B4) vs. Figure 3(A4)). In some of the specimens from Portugal, the condyle does not surpass the posterior margin of the neural spine in lateral view (Figure 6(C3) in [2]).
Regarding the cotyle being wider than the zygosphene, this is a feature unusually present in the specimens from Portugal, in which the cotyle is as wide as the zygosphene in most of the specimens. In fact, in the vertebra DCT-CC-1 (holotype of C. soaresi, considered as type species of the genus Segurasaurus), the cotyle is as wide as the zygosphene (p. 9, Figure 5A in [2]), as occurs in the paratypes DCT-CC-31, DCT-CC-7, DCT-CC-27, DCT-CC-2, and DCT-CC-29 (p. 9, Figures 6(C1,D1,E1,F1) and 7A1 in [2]) and in the new material studied herein (Figure 2B). Only in the specimen DCT-CC-8 (p. 10, Figure 7B1 in [2]) is the cotyle slightly wider than the zygosphene, while in DCT-CC-28, the zygosphene is slightly wider than the cotyle. Regarding the shape of the neural canal, it presents a subtriangular to subelliptical shape in the dorsal vertebrae DCT-CC-31 and DCT-CC-38, with it being subelliptical in DCT-CC-7 (pp. 9–10, Figures 6(C1,D1) and 7C1 in [2]) and even circular in DCT-CC-8 (p. 10, Figure 7B2 in [2]). Regarding the ‘V’-shaped centrum in ventral view, this “V” is clearly different in the specimens from France and in those from Portugal, as previously discussed (Figure 3(A5) vs. Figure 3(B5)). Moreover, not only the centrum shows a different shape but all of the vertebra in ventral view shows a dissimilar shape regarding the length of the zygosphene (not visible in ventral view of Carentonosaurus), and more projected prezygapophyses, paradiapophyses, and postzygapophyses (Figure 3(A5) vs. Figure 3(B5)). For this, and all of the differences previously mentioned, it is considered herein that the dorsal vertebrae of the specimens from Portugal show too many differences to be considered as the genus Carentonosaurus.
Comparing Segurasaurus soaresi with Carentonosaurus algorensis, they share certain morphological characteristics, but there are several relevant differences. In [2], no reference is made to the vertical narrow ridge or vertical keel existing in the middle part of the zygantrum, named zygantral lamina for the C. algorensis holotype (Figures 3(D1–D3) and 5R in [9]). However, this morphological structure is also visible in dorsal vertebrae specimens of S. soaresi, e.g., Figure 5B in [2]. Furthermore, a morphologically similar structure is noticeable in the zygosphene (Figure 3(B1,B2) and Figure 5A in [2]). Within Squamata, descriptions of the vertebral column are scarce, and this scarcity of morphological information might in part be due to a limited nomenclature of vertebral features, generating a greater difficulty in elaborating concise, detailed descriptions [27].
Both types (of S. soaresi and C. algorensis) display a subtriangular centrum with concave subcentral borders (Figure 5F in [2] and Figure 3(B1–B3) in [9]); marked posterior constriction of the neural arch (Figure 5E in [2] and Figure 3(A1–A3) in [9]); a prezygapophyseal width slightly larger than the postzygapophyseal width (Figure 5E in [2] and Figure 3(A1–A3) in [9]); highly laterally projected paradiapophyses, which go beyond the prezygapophyses (Figure 5E in [2] and Figure 3(A1–A3) in [9]); a subrectangular and low neural spine, ornamented with longitudinal grooves (Figure 5C in [2] and Figures 3(E1–E3) and 5O–S in [9]); a deeply notched zygosphene, highly projected towards the front and presenting concave lateral borders (Figure 5E in [2] and Figure 3(A1–A3) in [9]); the presence of a zygosphenal vertical keel (Figure 5A in [2] and Figure 3(C1,C2) in [9]); and the presence of a zygantral vertical keel and zygantral facets (Figure 5B in [2] and Figures 3(D1–D3) and 5N–R in [9]). However, according to the diagnosis of C. algorensis published in [9] (p. 9), there are some differences between both materials: the epidiapophyseal and interzygapophyseal ridges were absent in the studied vertebra from Algora (central Spain); no lateral, parazygosphenal, paracotylar, and zygantral foramina are present either, and no sagittal furrow is mentioned in the ventral surface of the dorsal vertebrae published in [9]. Due to the similarities and differences between these Cenomanian specimens from Spain and Portugal, it is not clear whether they could be the same species. The material from Algora, a single vertebra, is, for the moment, too scarce to confirm this. In addition, no cervical vertebrae are known from this site.
5.2. Intracolumnar Position of the New Material
Within the Squamata, the new vertebra described herein is interpreted as a dorsal vertebra due to the lack of pleurapophyses, which are characteristically present in sacral and proximal caudal vertebrae, and due to the absence of haemapophyses and a chevron-shaped haemal arch, typically present in caudal vertebrae, e.g., [28,29]. Moreover, it should occupy a further posterior location in the dorsal section of the spinal column, as it is extremely dorsoventrally depressed and displays a lesser extent of or non-pachyostotic morphology, mainly visible through the paradiapophyses ventral protuberances [2,30].
6. Conclusions
The obvious differences between the morphological characteristics of the dorsal vertebrae of Carentonosaurus soaresi and those described in the diagnosis made by Rage and Néraudeau [1] for the genotype Carentonosaurus mineaui justify the establishment of a new genus for those specimens of marine Pythonomorpha from the uppermost middle Cenomanian of Portugal.
The new genus Segurasaurus (type species S. soaresi) is diagnosable by the presence of cervical and dorsal vertebrae with paradiapophyses highly laterally projected beyond the prezygapophyses. The epi-diapophyseal and interzygapophyseal ridges are poorly defined and usually merged. The neural spine (characteristically low and subrectangular in dorsal vertebrae) is ornamented with longitudinal grooves. They show lateral and subcentral foramina, in addition to paracotylar, parazygosphenal, and zygantral ones. The zygosphenal and zygantral longitudinal keels are also diagnostic of this new genus.
Segurasaurus is also diagnosable by the presence of pachyostotic mid- to less posterior dorsal vertebrae which show a subtriangular shape in ventral view, with lateral concave borders and a sagittal furrow. The zygosphene, which is deeply notched and highly projected, with it exceeding the length of the centrum and being as long as the prezygapophyses in dorsal and ventral views, is also characteristic of Segurasaurus.
Author Contributions
Conceptualization, M.B.-C., R.P. and P.M.C.; data curation, M.B.-C., R.P. and P.M.C.; funding acquisition, M.B.-C., P.M.C., F.B.-B. and S.O.d.J.; investigation M.B.-C., R.P. and P.M.C.; methodology, M.B.-C., R.P. and P.M.C.; resources, M.B.-C., R.P., P.M.C., F.B.-B. and S.O.d.J.; software, M.B.-C. and P.M.C.; supervision, M.B.-C., R.P. and P.M.C.; writing—original draft, M.B.-C., R.P. and P.M.C.; writing—review and editing, M.B.-C., R.P., P.M.C., F.B.-B. and S.O.d.J. All authors have read and agreed to the published version of the manuscript.
Funding
Contracts Margarita Salas UCM CT31/21 (M.B.-C.) from the Complutense University of Madrid (Spain) and UI/BD/150971/2021 (S.O.d.J.) through the Earth and Space Research Center of the University of Coimbra (CITEUC) of the Foundation for Science and Technology (FCT, Portugal), Strategic Project UID/Multi/00611/2020 of the CITEUC, and Research Project SBPLY/21/180501/000242 of the Junta de Castilla-La Mancha and the University of Alcalá (Spain).
Data Availability Statement
The new genus described in this paper has been registered in Zoobank LSIDurn:lsid:zoobank.org:act:E02ABED7-5278-4BFB-B958-79DC9711CB86.
Acknowledgments
We give thanks to Manuel Segura of the University of Alcalá (UAH, Spain) and Julia Audije-Gil of the Autonomous University of Madrid (UAM, Spain). We thank the anonymous reviewers and the editors of Geosciences. The authors are thankful to the CITEUC (strategic project UID/Multi/00611/2020) and the FEDER—European Regional Development Fund through the COMPETE 2020—Competitiveness and Internationalization Operational Program (project POCI-01-0145-FEDER-006922).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rage, J.C.; Néraudeau, D. A new pachyostotic squamate reptile from the Cenomanian of France. Palaeontology 2004, 47, 1195–1210. [Google Scholar] [CrossRef]
- Pimentel, R.J.; Audije-Gil, J.; Barroso-Barcenilla, F.; Berrocal-Casero, M.; Callapez, P.M.; Ozkaya de Juanas, S.; Segura, M. On the occurrence of Carentonosaurus Rage and Néraudeau, 2004 (Squamata, Pythonomorpha) in the Iberian Cretaceous, with description of a new species from the uppermost middle Cenomanian of West Portugal. Cretac. Res. 2023, 151, 105469. [Google Scholar] [CrossRef]
- Bardet, N.; Houssaye, A.; Rage, J.C.; Pereda Suberbiola, X. The Cenomanian-Turonian (late Cretaceous) radiation of marine squamates (Reptilia): The role of the Mediterranean Tethys. Bull. Soc. Géol. Fr. 2008, 179, 605–622. [Google Scholar] [CrossRef]
- Vullo, R.; Néraudeau, D. Cenomanian vertebrate assemblages from southwestern France: A new insight into the European mid-Cretaceous continental fauna. Cret. Res. 2008, 29, 935–940. [Google Scholar] [CrossRef]
- Vullo, R.; Rage, J.C.; Néraudeau, D. Anuran and Squamate remains from the Cenomanian (Late Cretaceous) of Charentes, Western France. J. Vert. Paleont. 2011, 31, 279–291. [Google Scholar] [CrossRef]
- Vullo, R.; Bernárdez, E.; Buscalioni, A. Vertebrates from the middle?-late Cenomanian La Cabaña Formation (Asturias, northern Spain): Palaeoenvironmental and palaeobiogeographic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 276, 120–129. [Google Scholar] [CrossRef]
- Hontecillas, D.; Knoll, F.; Arsuaga, J.L.; Laplana, C.; Pérez-González, A.; Baquedano, E. Reworked remains of mosasauroids (Pythonomorpha, Squamata) in the Pleistocene of Pinilla del Valle (Madrid, Spain). In Proceedings of the 10th Annual Meeting of the European Association of Vertebrate Palaeontologists, Teruel, Spain, 19–24 June 2012; Royo-Torres, R., Gascó, F., Alcalá, L., Eds.; Conjunto Paleontológico de Teruel—Dinópolis: Teruel, Spain, 2012; p. 113. [Google Scholar]
- Hontecillas, D.; Houssaye, A.; Laplana, C.; Sevilla, P.; Arsuaga, J.L.; Pérez-González, A.; Baquedano, E.; Knoll, F. Reworked marine pythonomorph (Reptilia, Squamata) remains in Late Pleistocene cave deposits in central Spain. Cret. Res. 2015, 54, 188–202. [Google Scholar] [CrossRef]
- Cabezuelo-Hernández, A.; Pérez-García, A. A new species of the Pythonomorph Carentonosaurus from the Cenomanian of Algora (Guadalajara, Central Spain). Animals 2023, 13, 1197. [Google Scholar] [CrossRef]
- Barbosa, B.; Soares, A.F.; Rocha, R.B.; Manuppella, G.; Henriques, M. Carta Geológica de Portugal, Escala 1:50 000. Notícia Explicativa da Folha 19A—Cantanhede; Serviços Geológicos de Portugal: Lisboa, Portugal, 1988; pp. 1–46. [Google Scholar]
- Callapez, P.M. Palaeobiogeographic evolution and marine faunas of the Mid-Cretaceous Western Portuguese Carbonate Platform. Thalassas 2008, 24, 29–52. [Google Scholar]
- Choffat, P.L. Recueil de Monographies Stratigraphiques sur le Système Crétacique du Portugal—Deuxième Étude—Le Crétacé Supérieur au Nord du Tage; Direction des Services Géologiques du Portugal: Lisbonne, Portugal, 1900; pp. 1–287. [Google Scholar]
- Soares, A.F. Estudo das formações pós-jurássicas das regiões de entre Sargento-Mor e Montemor-o-Velho (margem direita do Rio Mondego). Mem. Notíc. 1966, 62, 1–343. [Google Scholar]
- Soares, A.F. Contribuição para o estudo do Cretácico em Portugal (o Cretácico superior da Costa de Arnes). Mem. Notíc. 1972, 74, 1–56. [Google Scholar]
- Soares, A.F. A “Formação Carbonatada” na região do Baixo-Mondego. Com. Serv. Geol. Port. 1980, 66, 99–109. [Google Scholar]
- Callapez, P.M. Estratigrafia e Paleobiologia do Cenomaniano-Turoniano. O significado do eixo da Nazaré-Leiria-Pombal. Ph.D. Thesis, University of Coimbra, Coimbra, Portugal, 1998. [Google Scholar]
- Callapez, P.M. The Cenomanian-Turonian central West Portuguese carbonate platform. In Cretaceous and Cenozoic Events in West Iberia Margins Field Trip Guidebook 2; Dinis, J., Cunha, P.P., Eds.; FCTUC: Coimbra, Portugal, 2004; pp. 39–51. [Google Scholar]
- Barroso-Barcenilla, F.; Callapez, P.M.; Soares, A.F.; Segura, M. Cephalopod assemblages and depositional sequences from the upper Cenomanian and lower Turonian of the Iberian Peninsula (Spain and Portugal). J. Iber. Geol. 2011, 37, 9–28. [Google Scholar]
- Segura, M.; Barroso-Barcenilla, F.; Callapez, P.M.; García-Hidalgo, J.F.; Gil-Gil, J. Depositional sequences and cephalopod assemblages in the upper Cenomanian-lower Santonian of the Iberian Peninsula (Spain and Portugal). Geol. Acta 2014, 12, 19–27. [Google Scholar]
- Berthou, P.Y. Albian-Turonian stage boundaries and subdivisions in the Western Portuguese Basin, with special emphasis on the Cenomanian-Turonian boundary in the Ammonite Facies and Rudist Facies. Bull. Geol. Soc. Denmark 1984, 33, 41–45. [Google Scholar] [CrossRef]
- Lauverjat, J. Le Crétacé Supérieur dans le Nord du Bassin Occidental Portugais. Ph.D. Thesis, Université Pierre et Marie Curie, Paris, France, 1982. [Google Scholar]
- Mateus, O.; Puértolas-Pascual, E.; Callapez, P.M. A new eusuchian crocodylomorph from the Cenomanian (Late Cretaceous) of Portugal reveals novel implications on the origin of Crocodylia. Zool. J. Linn. Soc. 2018, 186, 521–528. [Google Scholar] [CrossRef]
- Auffenberg, W. The fossil snakes of Florida. Tulane Stud. Zool. 1963, 10, 131–216. [Google Scholar]
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis, 10th ed.; Laurentius Salvius: Holmiae, Sweden, 1758; Volume 1, pp. 1–824. [Google Scholar]
- Oppel, M. Suite du 1er mémoire sur la classification des reptiles. Ord. II. Squammata mihi. Sect. II. Ophidii. Ord. III. Ophidii, Brongniart. Ann. Mus. Hist. Nat. 1811, 16, 376–393. [Google Scholar]
- Cope, E.D. On the reptilian orders Pythonomorpha and Streptosauria. Proc. Boston Soc. Nat. Hist. 1869, 12, 251–266. [Google Scholar]
- Tschopp, E. Nomenclature of vertebral laminae in lizards, with comments on ontogenetic and serial variation in Lacertini (Squamata, Lacertidae). PLoS ONE 2016, 11, e0149445. [Google Scholar] [CrossRef] [PubMed]
- Čerňanský, A.; Yaryhin, O.; Ciceková, J.; Werneburg, I.; Hain, M.; Klembara, J. Vertebral comparative anatomy and morphological differences in Anguine Lizards with a special reference to Pseudopus apodus. Anat. Rec. 2019, 302, 232–257. [Google Scholar] [CrossRef] [PubMed]
- Georgalis, G.L.; Scheyer, T.M. Lizards and snakes from the earliest Miocene of Saint-Gérand-le-Puy, France: An anatomical and histological approach of some of the oldest Neogene squamates from Europe. BMC Ecol. Evol. 2021, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Houssaye, A. A new aquatic pythonomorph (Reptilia, Squamata) from the Turonian (Late Cretaceous) of France. Comptes Rendus Palevol 2010, 9, 39–45. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).