
Ostracod Response to a Major Middle Jurassic Sea-Level Fall: A Case Study from Southern Tunisia (North Gondwana) with Implications on Regional Stratigraphy and Palaeoenvironmental Reconstruction

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
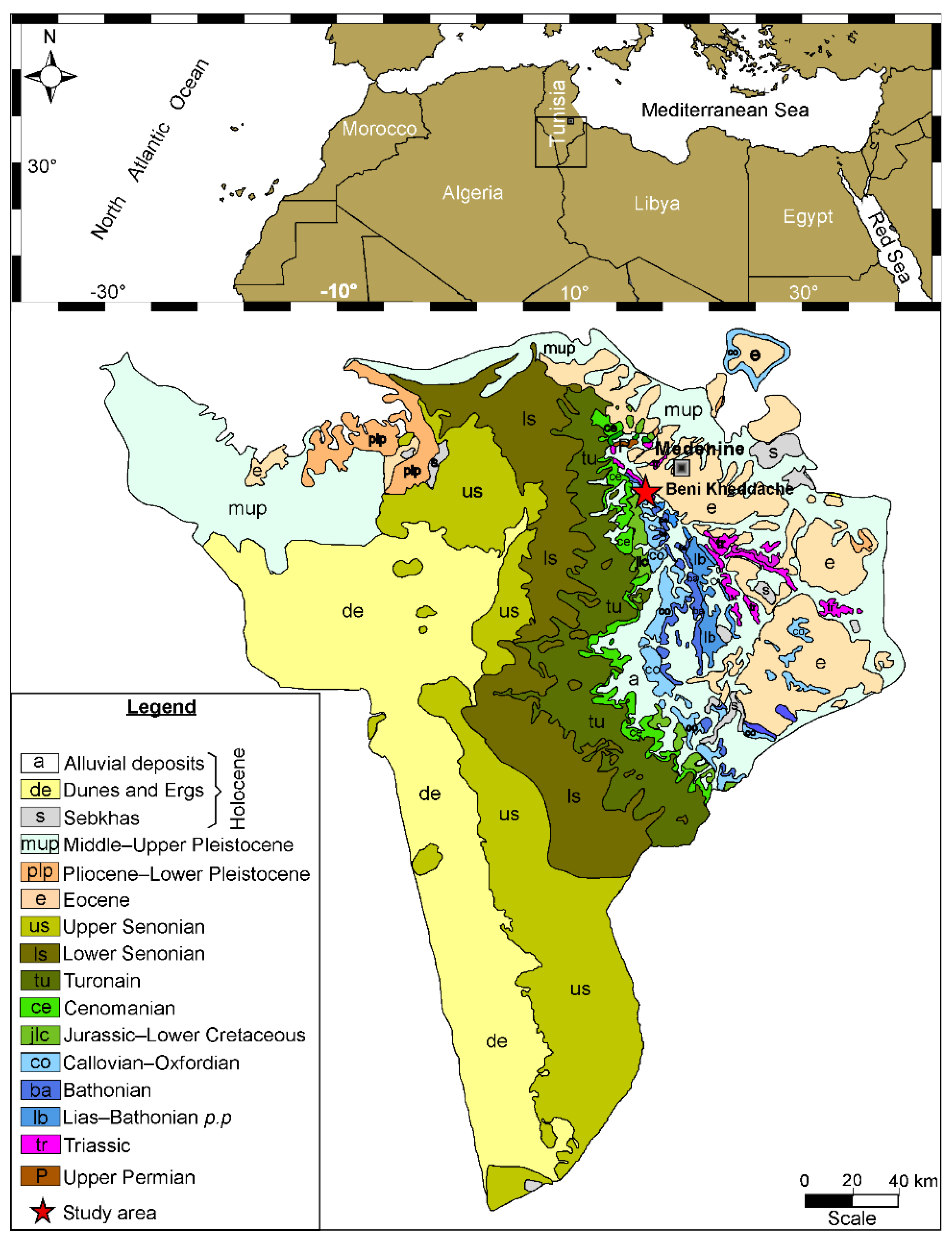
2. Geological Setting
3. Material and Methods
4. Result: Systematic Description
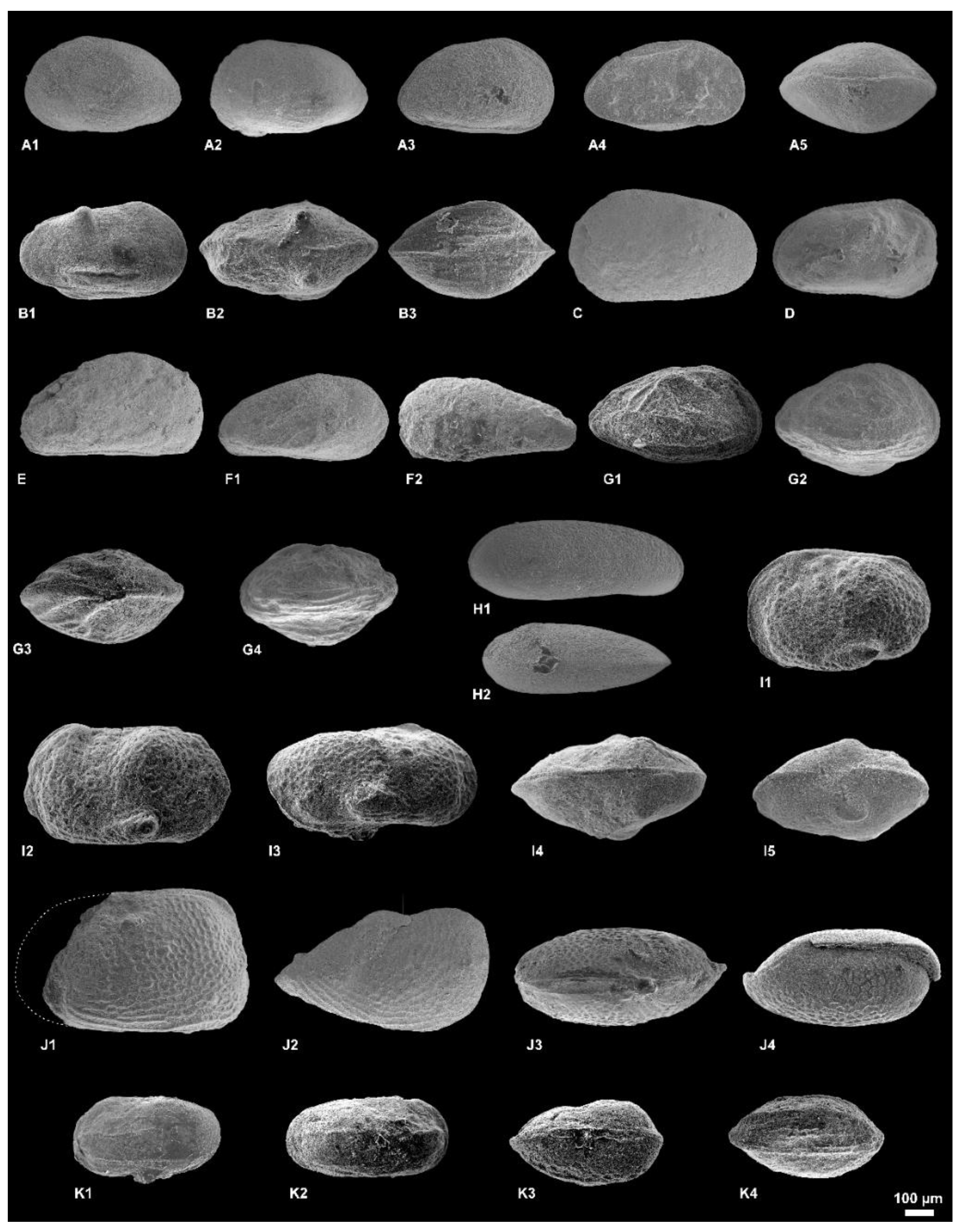
5. Discussion
5.1. Ostracod Assemblages and Depositional Palaeoenvironments
5.2. Biostratigraphy and Implication on Regional Stratigraphic Correlation
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andreu, B. Ostracodes du Bathonien Moyen et Supérieur des Grands-Causses, Sud de la France. Associations et Paléoenvironnements. Rev. Micropal. 1999, 42, 187–211. [Google Scholar] [CrossRef]
- Azerêdo, A.C.; Cabral, M.C.; Ramalho, M.M.; Pereira, R. Overview of Microfossil Assemblages and Paleoecological Signatures in the Middle-Upper Jurassic Transitional Successions from the Lusitanian Basin, Portugal. Comm. Inst. Geol. Min. 2002, 89, 155–178. [Google Scholar]
- Bate, R.H. Middle Jurassic Ostracoda from North Lincolnshire. Bull. Brit. Mus. Nat. Hist. Geol. 1963, 8, 173–219. [Google Scholar]
- Bate, R.H. Middle Jurassic Ostracoda from South Yorkshire. Bull. Brit. Mus. Nat. Hist. Geol. 1963, 9, 19–46. [Google Scholar]
- Bate, R.H. Freshwater ostracods from the Bathonian of Oxfordshire. Palaeontology 1965, 8, 749–756. [Google Scholar]
- Bate, R.H. Middle Jurassic Ostracoda from the Grey Limestone Series, Yorkshire. Bull. Brit. Mus. Nat. Hist. Geol. 1965, 11, 73–134. [Google Scholar]
- Bate, R.H. The Bathonian Upper Estuarine Series of eastern England Part 1: Ostracoda. Bull. Brit. Mus. Nat. Hist. Geol. 1967, 14, 21–66. [Google Scholar]
- Bate, R.H. Middle Jurassic (Aalenian–Bathonian). In Ostracods in British Stratigraphy; Whittaker, J.E., Hart, M.B., Eds.; Micropalaeontological Society, Special Publication, Geological Society: London, UK, 2009; pp. 100–223. [Google Scholar]
- Colin, J.P. Les Ostracodes Limniques du Jurassique Européen. In Biostratigraphie du Jurassique Ouest-Européen et Méditerranéen; Cariou, P., Hantzpergue, P., Eds.; Bulletin du Centre de Recherches Elf Exploration Production: Boussens, France, 1997; Volume 17, pp. 273–279. [Google Scholar]
- Oertli, H.J. Faunes d’Ostracodes du Mésozoïque de France; Brill: Leiden, The Netherlands, 1963; p. 57. [Google Scholar]
- Dépêche, F. Les Ostracodes d’une Plateforme Continentale au Jurassique, Recherches sur le Bathonien du Bassin Parisien; Unpublished, these d’état; Université. Pierre-et-Marie Curie: Paris, France, 1984; Volume 84, p. 419. [Google Scholar]
- Dépêche, F. Lias superieur, Dogger, Malm. In Atlas des Ostracodes de France; Oertli, H.J., Ed.; Bulletin du Centre de Recherches Elf Exploration Production: Boussens, France, 1985; Volume 9, pp. 119–124. [Google Scholar]
- Mojon, P.O. Les Formations Mésozoïques à Charophytes (Jurassique moyen-Crétacé inférieur) de la Marge Téthysienne Nord-Occidentale (Sud-Est de la France, Suisse Occidentale, Nord-Est de l’Espagne). Sédimentologie, Micropaléontologie, Biostratigraphie. Ph.D. Thesis, Université Joseph-Fourier-Grenoble I, Saint-Martin-d’Estréaux, France, 2001. [Google Scholar]
- Oertli, H.J. Ostracodes Lacustres du Bathonian du Poitou (Bassin de Paris); Bernard, F., Bizon, J.J., Oertli, H.J., Eds.; Bulletin de la Société Géologique de France: Paris, France, 1957; Volume 6, pp. 753–770. [Google Scholar]
- Rohr, W.M. Mitteljurassische Ostracoden aus den Grands Causses Süd-Frankreichs. Ph.D Thesis, Freie Universität, Berlin, Germany, 1976; p. 166. [Google Scholar]
- Sheppard, L.M. Middle Jurassic Ostracoda from Southern England and Northern France. Ph.D Thesis, University of London, London, UK, 1981. [Google Scholar]
- Wakefield, M.I. Ostracoda and Palaeosalinity Fluctuations in the Middle Jurassic Lealt Shale Formation, Inner Hebrides, Scotland. Palaeontology 1995, 38, 583–617. [Google Scholar]
- Wienholz, E. Neue Ostracoden aus dem Norddeutschen Callov. Freib. Forsch. Paläontologie 1967, 213, 23–51. [Google Scholar]
- Mette, W. Ostracods from the Middle Jurassic of Southern Tunisia. Beringeria 1995, 16, 259–348. [Google Scholar]
- Mette, W. Palaeoecology and Palaeobiogeography of the Middle Jurassic Ostracods of Southern Tunisia. Palaeogeog. Palaeoclimat. Palaeoecol. 1997, 131, 65–111. [Google Scholar] [CrossRef]
- Kamoun, F.; Peybernès, B.; Montacer, M.; Ben Youssef, M. Sequence Stratigraphy, Micropalaeontology and Palaeogeography of the Jurassic Series of Southern Tunisia. In The Geology of Northwest Libya. Second Symposium on the Sedimentary Basins of Libya; Salem, M.J., Oun, K.K., Seddiq, H.M., Eds.; Earth Science Society of Libya (ESSL): Tripoli, Libya, 2000; pp. 79–98. [Google Scholar]
- Busson, G. Le Mésozoïque Saharien. 1ère Partie: L’Extrême Sud Tunisien. Cent. Nat. Rech. Sci. Paris Géol. 1967, 8, 1–204. [Google Scholar]
- Walley, C.D. Depositional History of Southern Tunisia and Northwestern Libya in Mid and Late Jurassic Time. Geol. Magaz. 1985, 212, 233–247. [Google Scholar] [CrossRef]
- Kamoun, F. Le Jurassique du Sud Tunisien, Témoin de la Marge Africaine de la Téthys; Stratigraphie, Sédimentologie et Micropaléontologie. Ph.D. Thesis, Université de Toulouse, Toulouse, France, 1988; p. 330. [Google Scholar]
- Kamoun, F.; Ben Youssef, M.; Peybernès, B. Stratigraphie Séquentielle du Dogger et de la Base du Malm (Intervalle Aalénien–Kimméridgien) de l’Extrême-Sud de la Tunisie. Com. Rend. Acad. Sci. Paris 1992, 315, 1373–1379. [Google Scholar]
- Ben Ismail, M.H. Mixed Evaporitic Shallow Marine Carbonate and Siliciclastic Sedimentation of Jurassic Sequences in Southeast Tunisia. In Proceedings of the 8th IAS Regional Meeting of Sedimentology, Tunis, Tunisia, 1–3 April 1987; International Association of Sedimentologists: Algier, Algeria, 1987; pp. 80–82. [Google Scholar]
- Ben Ismail, M.H.; M’Rabet, A. Evaporite, Carbonate, and Siliciclastic Transitions in the Jurassic Sequences of Southeastern Tunisia. Sed. Geol. 1990, 66, 65–82. [Google Scholar] [CrossRef]
- Bouaziz, S. La Déformation Dans la Plate-Forme du Sud Tunisien (Dahar et Jeffara): Approche Multiscalaire et Pluridisciplinaire. Ph.D. Thesis, Université de Tunis, Tunis, Tunisia, 1986; p. 180. [Google Scholar]
- Peybernès, B.; Alméras, Y.; Ben Youssef, M.; Kamoun, F.; Mello, J.; Rey, J.; Zargouni, F. Nouveaux Eléments de Datation Dans le Jurassique du Sud-Tunisien (Plate-Forme Saharienne). Comp. Rend. Acad. Sci. Paris 1985, 300, 113–119. [Google Scholar]
- Ben Ismail, M.H.; Bouaziz, S.; Alméras, Y.; Clavel, B.; Donze, P.; Enay, R.; Ghanmi, M.; Tintant, H. Nouvelles donnée biostratigraphiques sur le Callovien et les faciès "purbecko-wealdiens" (Oxfordien à Vraconien) dans la region de Tataouine (Sud tunisien). Bull. Soc. Géolog. France 1989, 8, 353–360. [Google Scholar] [CrossRef]
- Enay, R.; El Asmi, K.; Soussi, M.; Mangold, C.; Hantzpergue, P. Un Pachyerymnoceras Arabique Dans le Callovien Supérieur du Dahar (Sud Tunisien), Nouvel Elément de Datation du Membre Ghomrassène (Formation Tataouine); Corrélations Avec l’Arabie Saoudite et le Moyen-Orient. Comp. Rend. Geosci. 2002, 334, 1157–1167. [Google Scholar] [CrossRef]
- Alméras, Y.; Enay, R.; Mangold, C.; Soussi, M.; El Asmi, K.; Hantzpergue, P. Observations à la Note: Mise en Évidence d’une Flore Oxfordienne Dans le Sud-Est de la Tunisie: Intérêts Stratigraphique et Paléoécologique par M. Ouaja. Geobios 2004, 37, 89–97, Erratum in Geobios 2005, 38, 691–695. [Google Scholar]
- Tiss, L.; Trabelsi, K.; Kamoun, F.; Soussi, M.; Houla, Y.; Sames, B.; Martín-Closas, C. Middle Jurassic Charophytes from Southern Tunisia: Implications on Evolution and Paleobiogeography. Rev. Palaeob. Palynol. 2019, 263, 65–84. [Google Scholar] [CrossRef]
- Nötzold, T. Die Präparation von Gyrogoniten und Kalkigen Charophyten-Oogonien aus Festen Kalksteinen. Monat. Deut. Akad. Wiss. Berl. 1965, 7, 216–221. [Google Scholar]
- Trabelsi, K.; Touir, J.; Soulié-Märsche, I.; Martín-Closas, C.; Soussi, M.; Colin, J.P. Découverte des Charophytes de l’Albien Dans la Formation Kebar (Tunisie Centrale): Implications Paléoécologiques et Paléobiogéographiques. Ann. Paléontol. 2010, 96, 117–133. [Google Scholar] [CrossRef]
- Trabelsi, K.; Sames, B.; Salmouna, A.; Piovesan, E.K.; Ben Rouina, S.; Houla, Y.; Touir, J.; Soussi, M. Ostracods from the Marginal Coastal Lower Cretaceous (Aptian) of the Central Tunisian Atlas (North Africa): Paleoenvironment, Biostratigraphy and Paleobiogeography. Rev. Micropaléontol. 2015, 58, 309–331. [Google Scholar] [CrossRef]
- Trabelsi, K.; Soussi, M.; Touir, J.; Houla, Y.; Abbes, C.; Martín-Closas, C. Charophyte Biostratigraphy of the Non-Marine Lower Cretaceous in the Central Tunisian Atlas (North Africa): Palaeobiogeographic Implications. Cret. Res. 2016, 67, 66–83. [Google Scholar] [CrossRef]
- Trabelsi, K.; Sames, B.; Nasri, A.; Piovesan, E.K.; Ferhi, F.; Skanji, A.; Houla, Y.; Soussi, M.; Wagreich, M. Ostracods as Proxies for Marginal Marine to Non-Marine Intervals in the Mid-Cretaceous Carbonate Platform of the Central Tunisian Atlas (North Africa): Response to Major Short-Term Sea-Level Falls. Cret. Res. 2021, 117, 104581. [Google Scholar] [CrossRef]
- Martin, J.W.; Davis, G.E. An Updated Classification of the Recent Crustacea. Nat. Hist. Mus. Los Angel. Cty. Sci. Ser. 2001, 39, 1–124. [Google Scholar]
- Meisch, C.; Smith, R.J.; Martens, K. A Subjective Global Checklist of the Extant Non-Marine Ostracoda (Crustacea). Eur. J. Taxon. 2019, 492, 1–135. [Google Scholar] [CrossRef] [Green Version]
- Horne, D.J. Ostracod Biostratigraphy and Palaeoecology of the Purbeck Limestone Group in Southern England. Spec. Pap. Palaeontol. 2002, 68, 1–18. [Google Scholar]
- Soulimane, C.; Reolid, M.; Marok, A. Ostracod Assemblages from the Uppermost Pliensbachian and Lower Toarcian of the Traras Mountains (Tlemcen Domain, North Algeria). Arab. J. Geosci. 2017, 10, 1–24. [Google Scholar] [CrossRef]
- Malz, H.; Hofmann, K.; Radke, G.; Cherchi, A. Biostratigraphy of the Middle Jurassic of NW Sardinia by Means of Ostracods. Senckenb. Leth. 1985, 66, 299–345. [Google Scholar]
- Dépêche, F. Etude Micropaleontologique du Bathonien des Causses du Quercy (Region de Cajaro). Rev. Micropaleontol. 1968, 11, 210–216. [Google Scholar]
- Sames, B. Early Cretaceous Theriosynoecum Branson 1936 in North America and Europe. Micropaleontology 2011, 57, 291–344. [Google Scholar]
- Sames, B. Glossary of Morphologic Terms of Late Mesozoic Non-Marine Ostracoda, Relevant to Theriosynoecum Branson 1936 and Cypridea Bosquet 1852. Micropaleontology 2011, 57, 433–454. [Google Scholar]
- Sames, B. Combined References for Taxonomic Studies in Early Cretaceous Non-Marine Ostracoda of North America. Micropaleontology 2011, 57, 455–465. [Google Scholar]
- Martens, K.; Rossetti, G.; Horne, D.J. How Ancient Are Ancient Asexuals? Proc. R. Soc. Lond. Biol. Sci. 2003, 270, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Fürsich, F.T.; Freytag, S.; Röhl, J.; Schmid, A. Palaeoecology of Benthic Associations in Salinity-Controlled Marginal Marine Environments: Examples from the Lower Bathonian (Jurassic) of the Causses (Southern France). Palaeogeogr. Palaeoclim. Palaeoecol. 1995, 113, 135–172. [Google Scholar] [CrossRef]
- Neale, J.W. Ostracods and Palaeosalinity Reconstruction. In Ostracoda in the Earth Sciences; Dedeckker, P., Colin, J.P., Peypouquet, J.P., Eds.; Elsevier: Amsterdam, The Netherlands, 1988; pp. 125–155. [Google Scholar]
- Colin, J.P.; Dépêche, F. Faunes d’Ostracodes Lacustres des Bassins Intracratoniques d’âge Albo-Aptien en Afrique de l’Ouest (Cameroun, Tchad) et au Brésil: Considérations d’Ordre Paléoécologique et Paléobiogéographique. Afri. Geosci. Rev. 1997, 4, 431–450. [Google Scholar]
- Nye, E.; Feist-Burkhardt, S.; Horne, D.J.; Ross, A.J.; Whittaker, J.E. The Palaeoenvironment Associated with a Partial Iguanodon Skeleton from the Upper Weald Clay (Barremian, Early Cretaceous) at Smokejacks Brickworks (Ockley, Surrey, UK), Based on Palynomorphs and Ostracods. Cret. Res. 2008, 29, 417–444. [Google Scholar] [CrossRef]
- Ayress, M.A.; Whatley, R.C. Early Cretaceous non-Marine Ostracoda from the North Falkland Basin, South Atlantic. Palaeontology 2014, 57, 1143–1175. [Google Scholar] [CrossRef]
- Oertli, H.J. The Aspect of Ostracods Fauna: A possible new tool in petroleum sedimentology. In Paléoécologie des Ostracodes; Oertli, H.J., Ed.; Bulletin du Centre de Recherches: Paris, France, 1971; pp. 137–151. [Google Scholar]
- Weiß, M. Stratigraphie und Mikrofauna im Kimmeridge SE-Niedersachsens unter Besonderer Berücksichtigung der Ostracoden. Ph.D. Thesis, Technische Universität Clausthal, Clausthal-Zellerfeld, Germany, 1995. Volume 48. pp. 1–274. [Google Scholar]
- Boomer, I.; Horne, D.J.; Slipper, I.J. The Use of Ostracods in Palaeoenvironmental Studies, or What Can You Do with an Ostracod Shell. In Bridging the Gap-Trends in the Ostracode Biological and Geological Sciences; Park, L.E., Smith, A.J., Eds.; The Paleontological Society Papers: Cambridge, UK, 2003; Volume 9, pp. 153–179. [Google Scholar]
- Whatley, R.C. The application of Ostracoda to paleoenvironmental analysis. In Applications of Ostracoda, Proceedings of the Eighth International Symposium on Ostracoda, Houston, TX, USA, 26–29 July; Department of Geosciences, University of Houston: Houston, TX, USA, 1983; pp. 51–77. [Google Scholar]
- Browers, E.M. Sediment transport detected from the analysis of ostracod population structure: An example from the Alaskan continental shelf. In Ostracoda in Earth Sciences; Dedeckker, P., Colin, J.P., Peypouquet, J.P., Eds.; Elsevier: Amsterdam, The Netherlands, 1988; pp. 231–244. [Google Scholar]
- Crasquin-Soleau, S.; Vaslet, D.; Le Nindre, Y.M. Ostracods as Markers of the Permian/Triassic boundary in the Khuff Formation of Saudi Arabia. Palaeontoloy 2005, 48, 853–868. [Google Scholar] [CrossRef]
- Haq, B.U. Jurassic Sea-Level Variations: A Reappraisal. GSA Today 2017, 28, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Mojon, P.O.; Haddoumi, H.; Charriére, A. Nouvelles Données sur les Charophytes et Ostracodes du Jurassique Moyen-Supérieur-Crétacé Inférieur de l’Atlas Marocain; Carnets De Géologie/Notebooks on Geology: Brest, France, 2009; pp. 1–39. [Google Scholar]
- Dercourt, J.; Fourcade, E.; Cecca, F.; Azéma, J.; Enay, R.; Bassoullet, J.P.; Cottereau, N. Palaeoenvironment of the Jurassic System in the Western and Central Tethys (Toarcian, Callovian, Kimmeridgian, Tithonian): An Overview. Geobios 1994, 17, 625–644. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trabelsi, K.; Tiss, L.; Sames, B.; Houla, Y.; Hanini, A.; Elferhi, F.; Skanji, A.; Kamoun, F.; Zagrarni, M.F.; Wagreich, M. Ostracod Response to a Major Middle Jurassic Sea-Level Fall: A Case Study from Southern Tunisia (North Gondwana) with Implications on Regional Stratigraphy and Palaeoenvironmental Reconstruction. Geosciences 2022, 12, 93. https://doi.org/10.3390/geosciences12020093
Trabelsi K, Tiss L, Sames B, Houla Y, Hanini A, Elferhi F, Skanji A, Kamoun F, Zagrarni MF, Wagreich M. Ostracod Response to a Major Middle Jurassic Sea-Level Fall: A Case Study from Southern Tunisia (North Gondwana) with Implications on Regional Stratigraphy and Palaeoenvironmental Reconstruction. Geosciences. 2022; 12(2):93. https://doi.org/10.3390/geosciences12020093
Chicago/Turabian StyleTrabelsi, Khaled, Lassad Tiss, Benjamin Sames, Yassine Houla, Amine Hanini, Faycel Elferhi, Ahmed Skanji, Fekri Kamoun, Mohamed Faouzi Zagrarni, and Michael Wagreich. 2022. "Ostracod Response to a Major Middle Jurassic Sea-Level Fall: A Case Study from Southern Tunisia (North Gondwana) with Implications on Regional Stratigraphy and Palaeoenvironmental Reconstruction" Geosciences 12, no. 2: 93. https://doi.org/10.3390/geosciences12020093
APA StyleTrabelsi, K., Tiss, L., Sames, B., Houla, Y., Hanini, A., Elferhi, F., Skanji, A., Kamoun, F., Zagrarni, M. F., & Wagreich, M. (2022). Ostracod Response to a Major Middle Jurassic Sea-Level Fall: A Case Study from Southern Tunisia (North Gondwana) with Implications on Regional Stratigraphy and Palaeoenvironmental Reconstruction. Geosciences, 12(2), 93. https://doi.org/10.3390/geosciences12020093