
Palynostratigraphy, Palynofacies, T-R Cycles and Paleoenvironments in the Middle Jurassic–Early Cretaceous Ramså Basin, Andøya, Northern Norway


Abstract
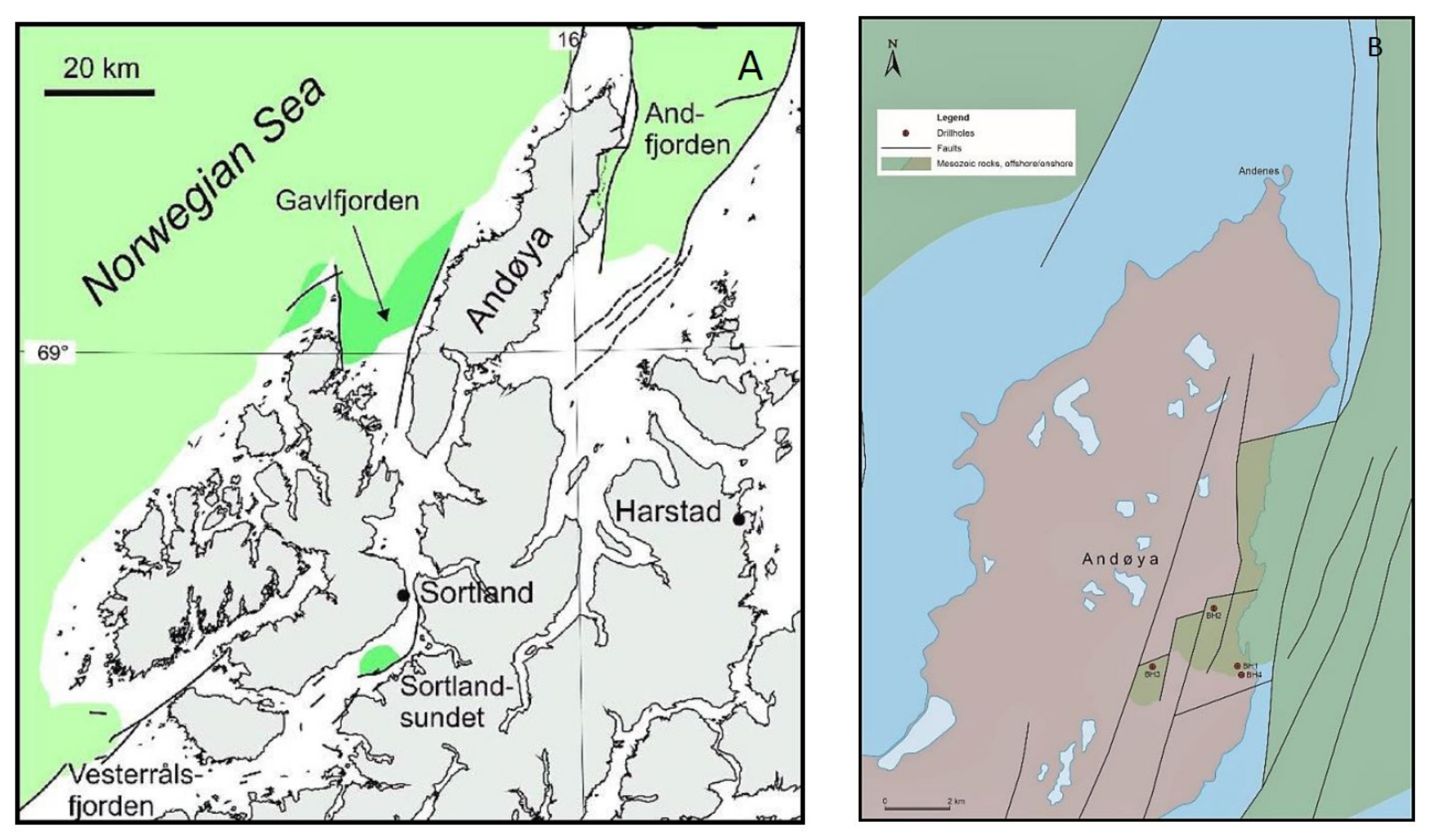
1. Introduction
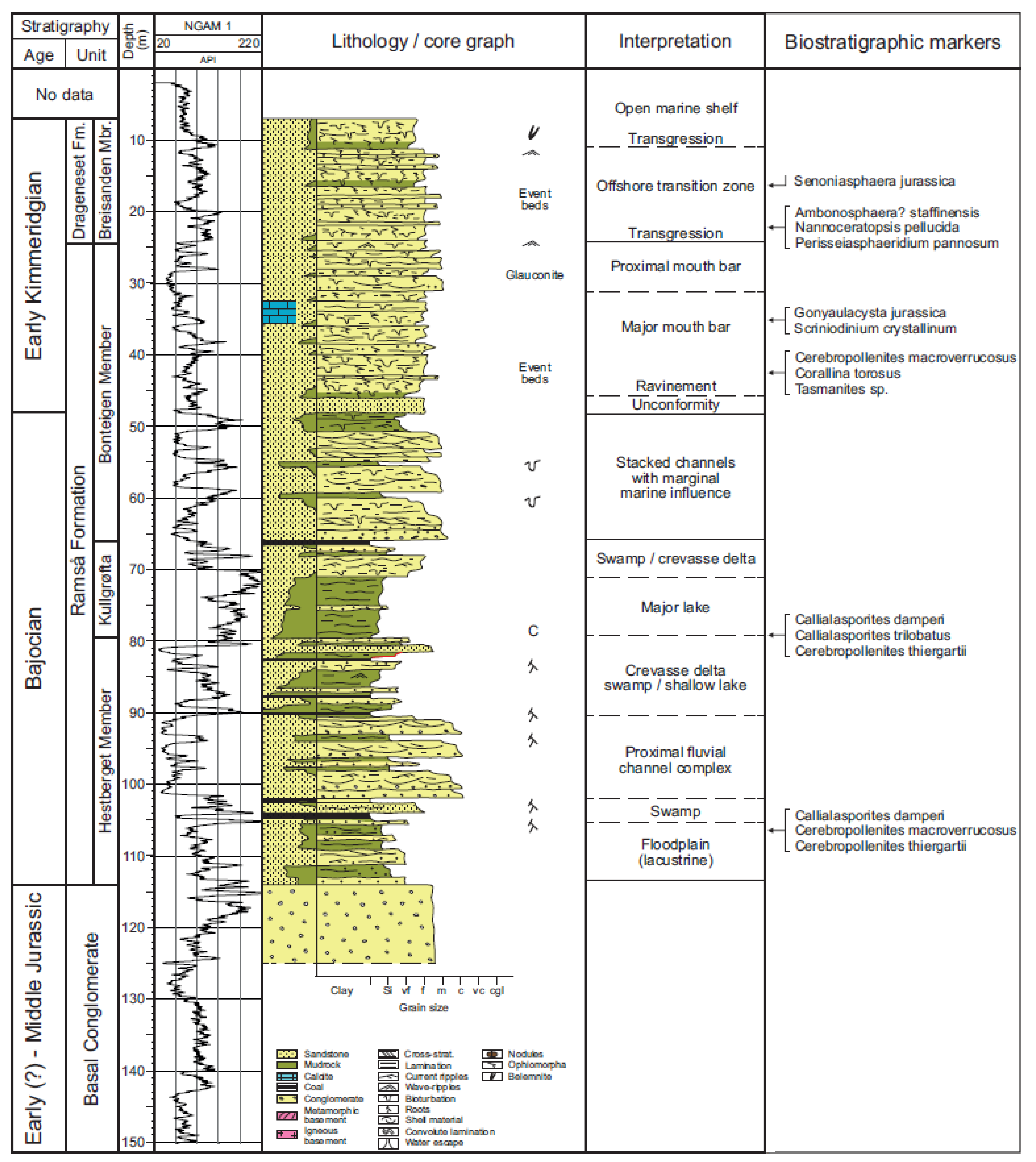
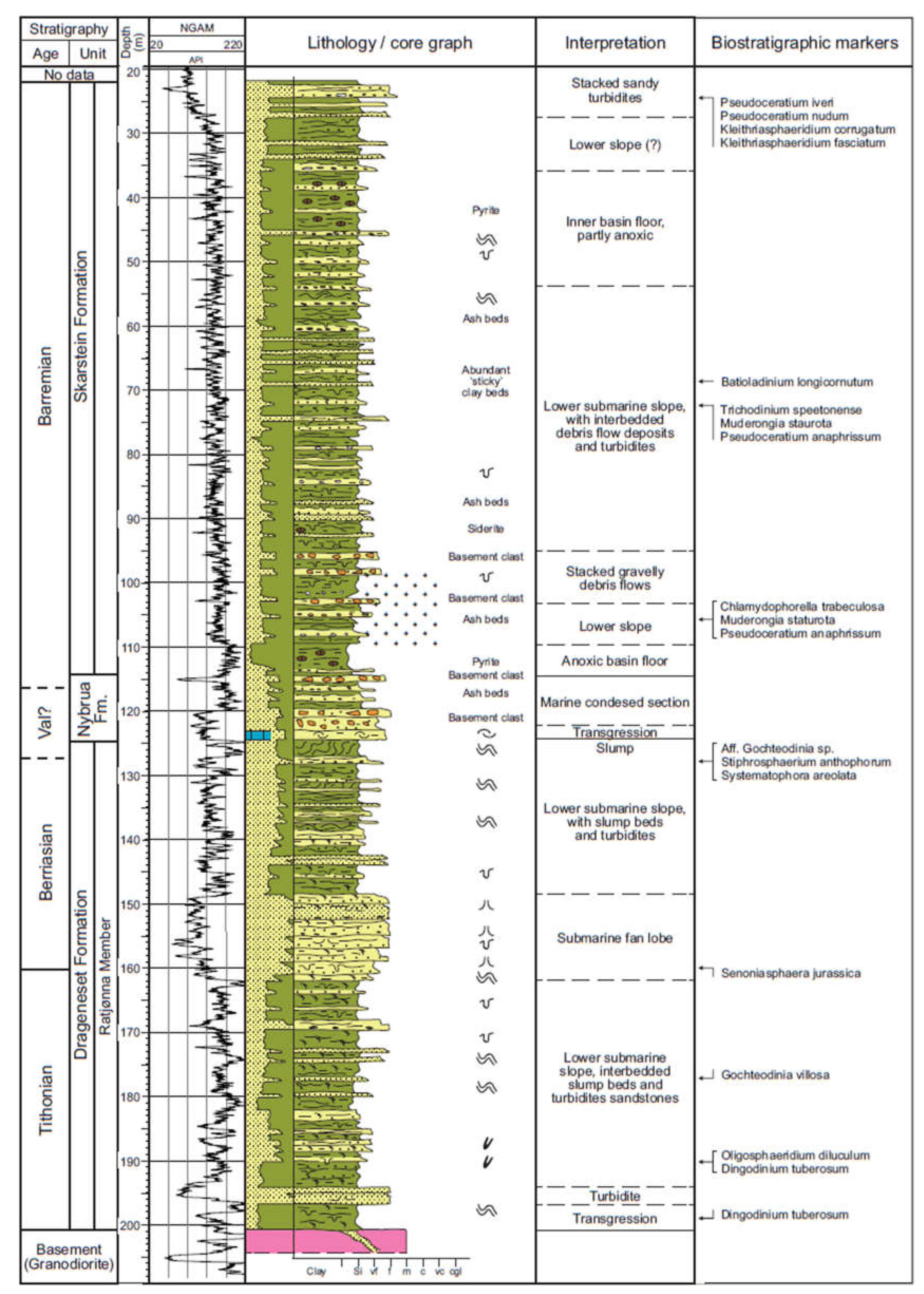
2. Lithostratigraphy
3. Material and Methods
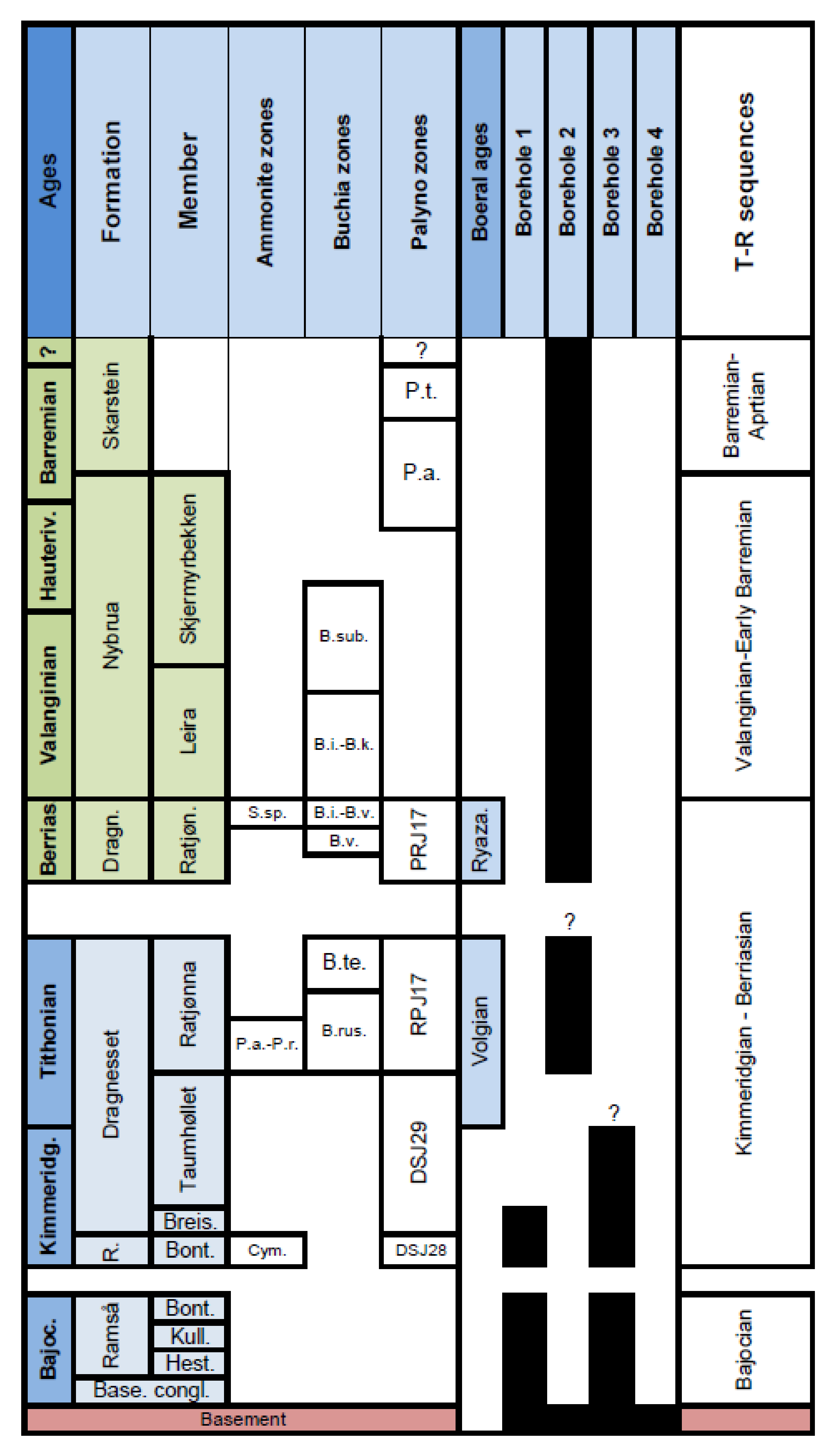
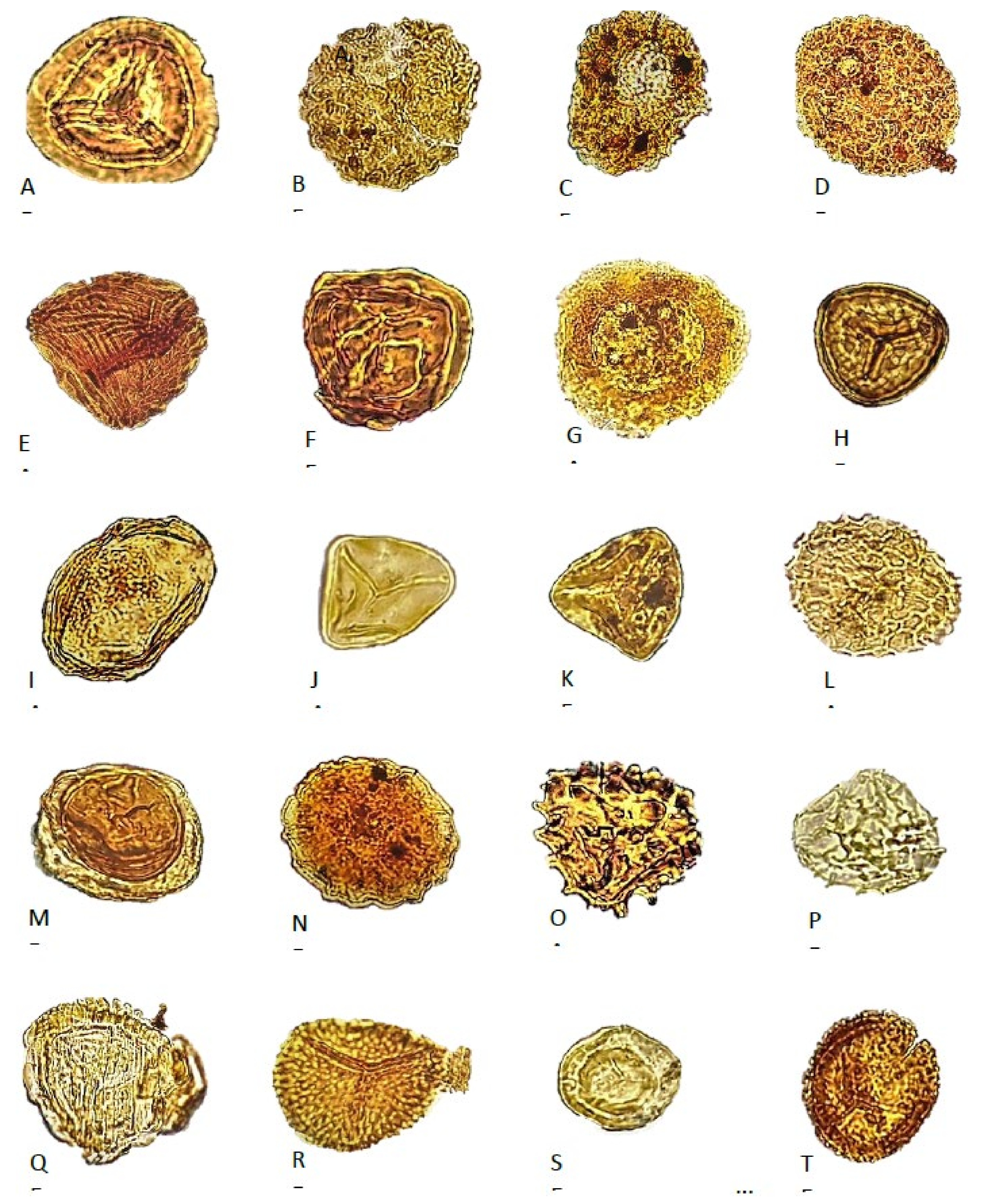
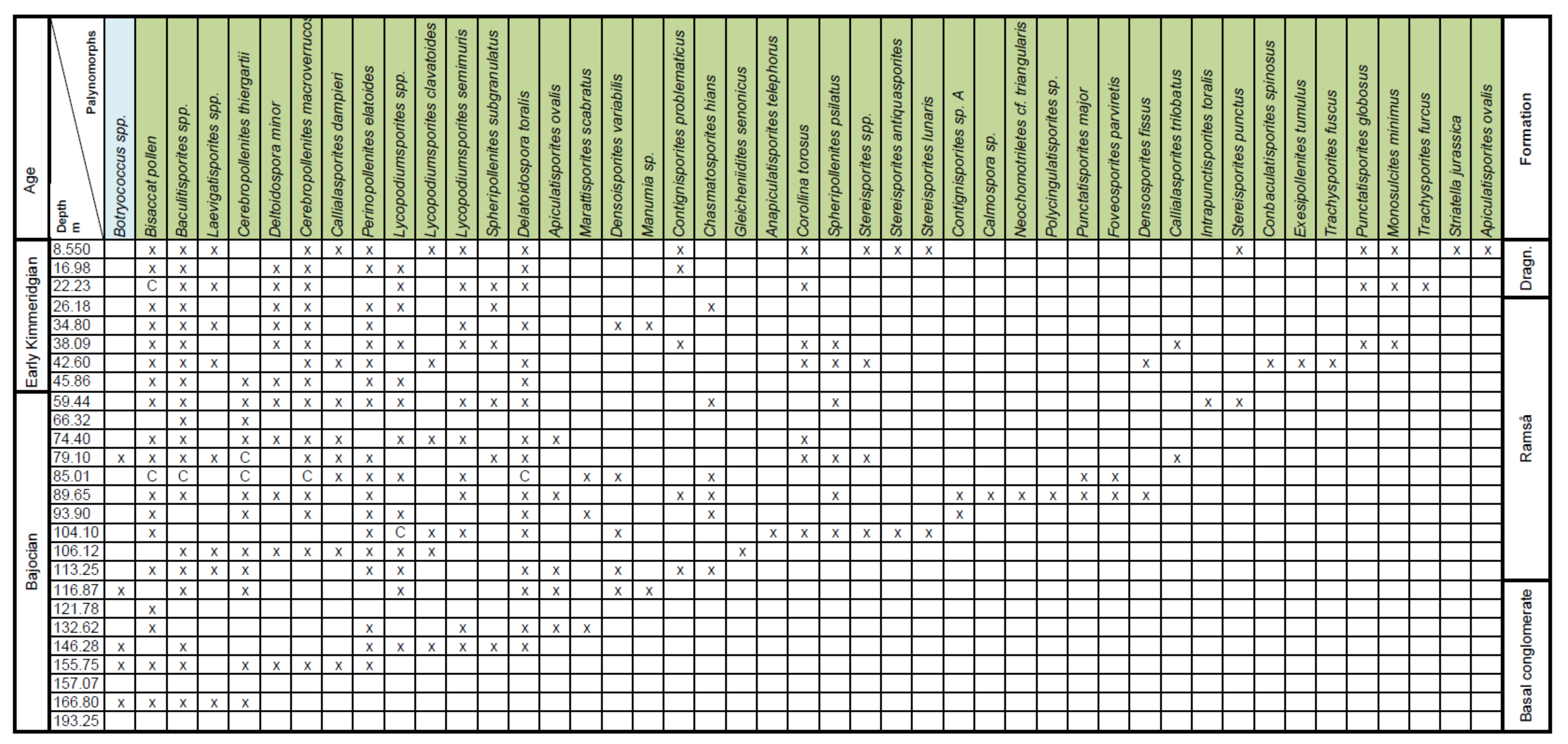
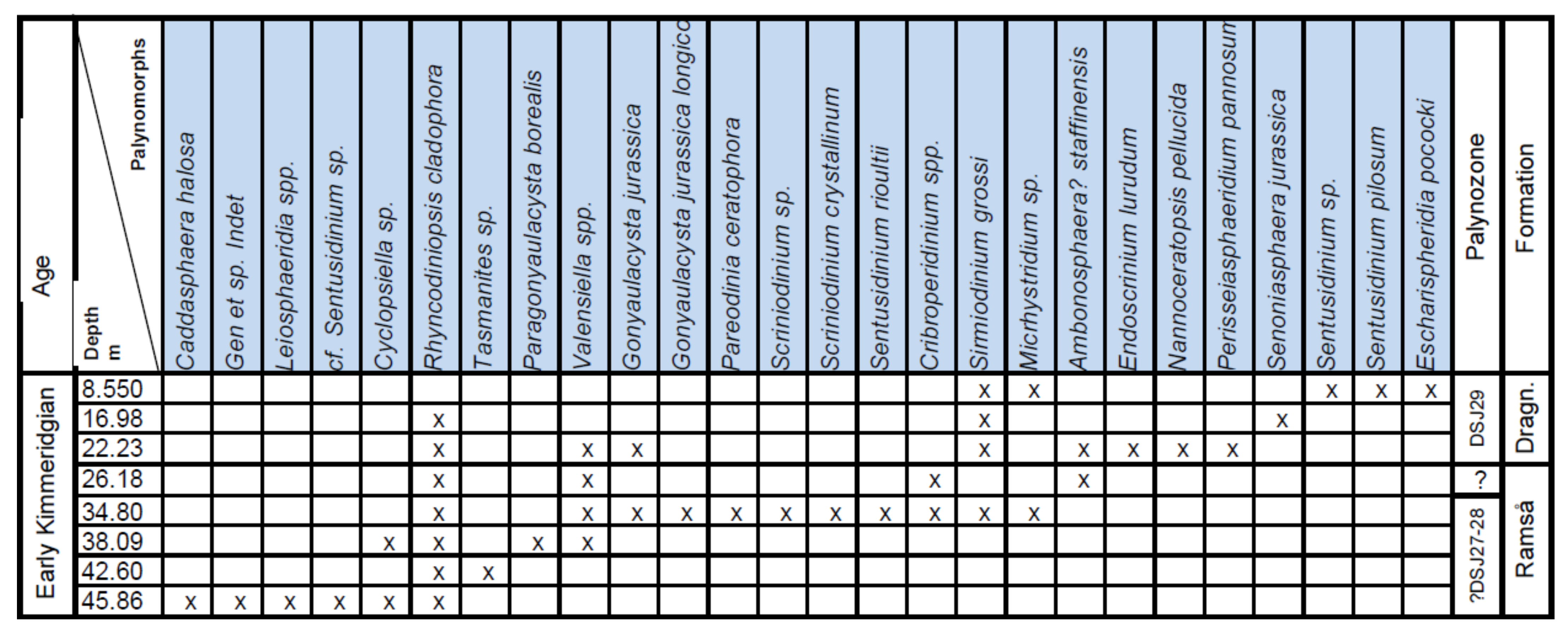
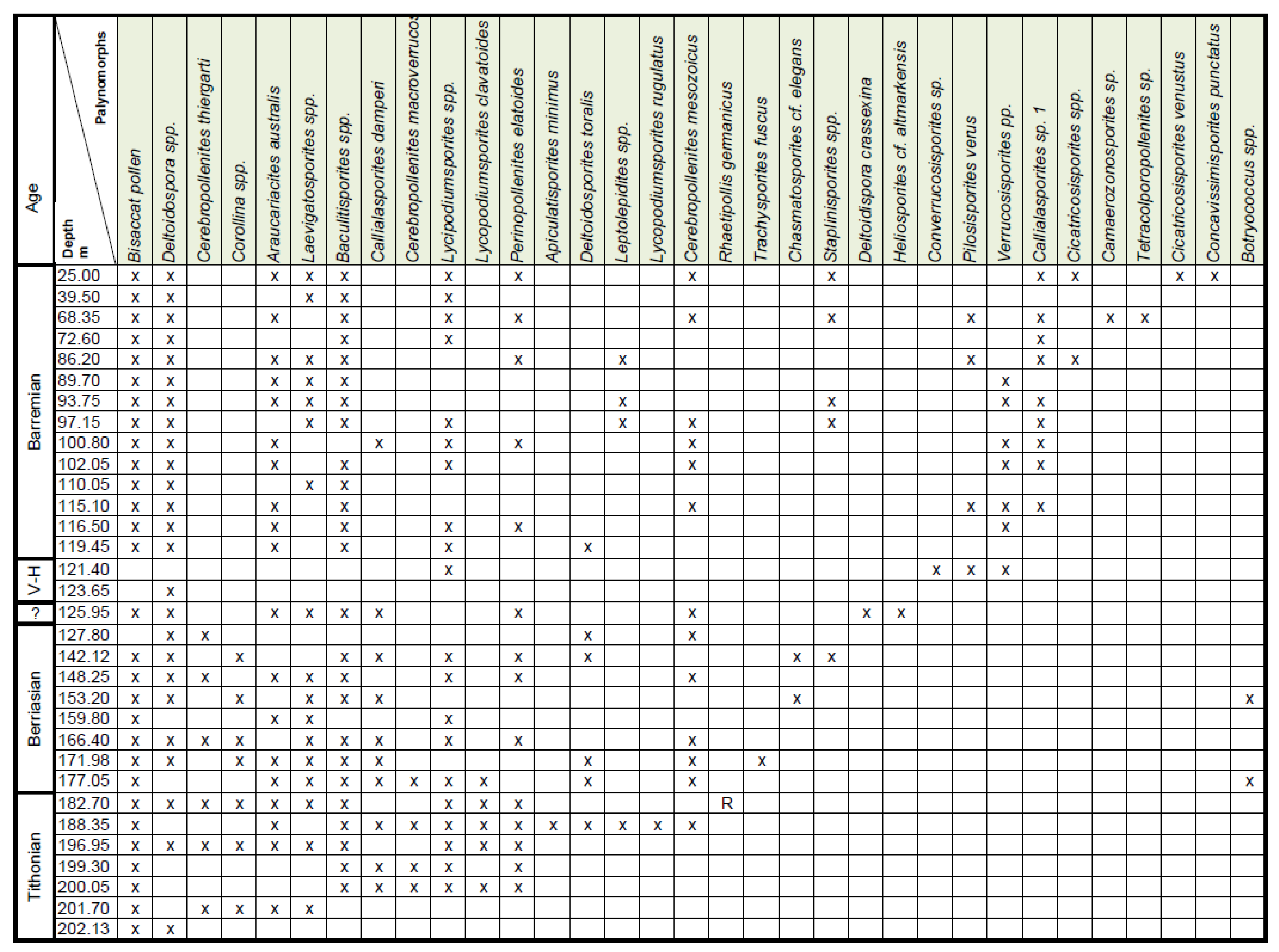
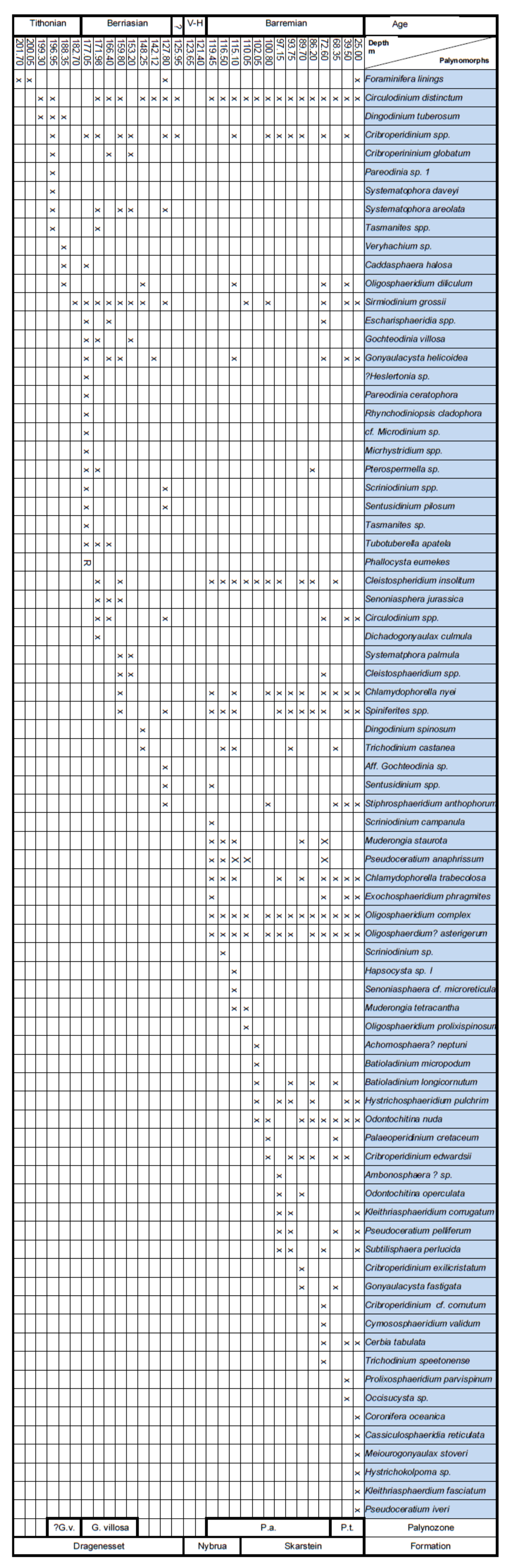
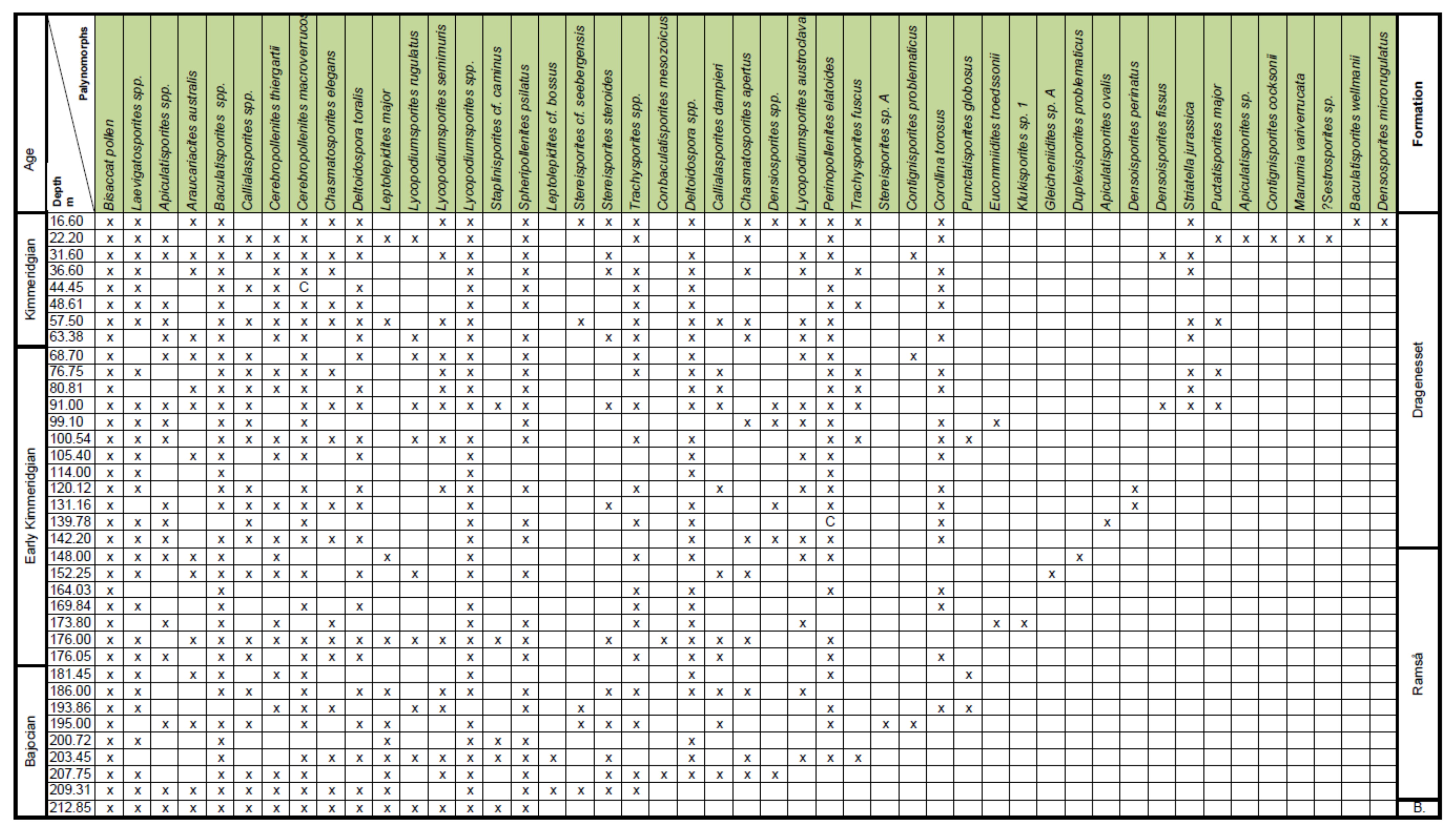
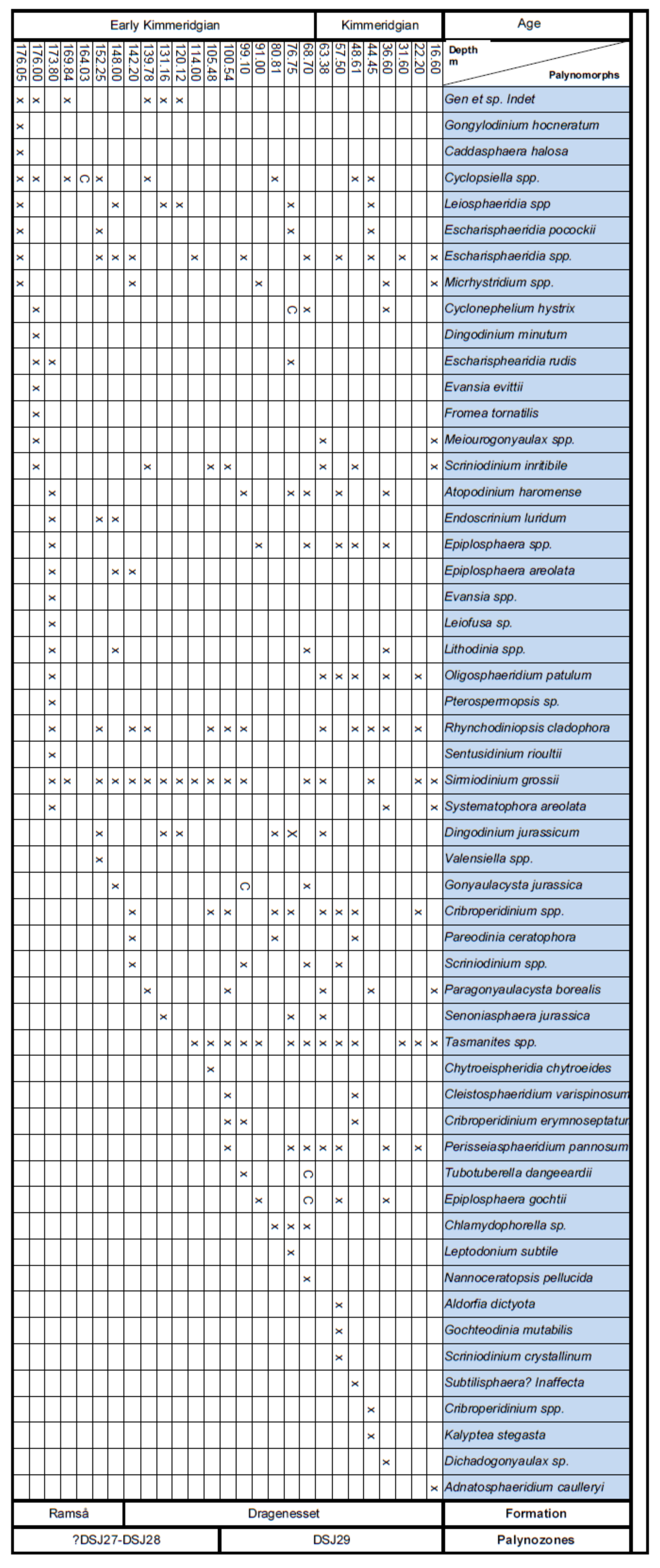
4. Palynostratigraphy
4.1. Borehole Bh 1
4.2. Borehole Bh 2
4.3. Borehole Bh 3
4.4. Borehole Bh 4
5. Palynofacies
6. Ages of Formations/Members, Stratigraphic Breaks and Transgressive Surfaces
7. T-R Cycles and Paleoenvironments
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A






References
- Kjerulf, T. Stenriget og Fjeldlæren; Werner & Co: Gelsenkirchen, Germany, 1870; pp. 272–275. [Google Scholar]
- Heer, O. Über die Pflanzen-Versteinerungen von Andö in Norwegen. Flora Foss. Arct. 1877, 4, 1–4. [Google Scholar]
- Lundgren, B. Faunaen i Andöns Jurabildningar. Christiania Vidensk.-Selsk. Forh. 1894, 5, 1–11. [Google Scholar]
- Friis, J.P. Andøens kulfelt. Norges Geol. Unders. 1903, 36, 38. [Google Scholar]
- Vogt, J.H.L. Om Andøens jurafelt, navnlig om landets langsomme nedsynken under juratiden og den senere hævning samt gravforkastning. Norges Geol. Unders. 1905, 43, 67. [Google Scholar]
- Sokolov, N. Fauna der mesozoischen Ablagerungen von Andö. Vitensk. Kristianias Skr. Mat. Nat. Kl. 1912, 6, 1–16. [Google Scholar]
- Ørvig, T. On the Mesozoic field of Andøya 1. Notes on the Ichthyosaurian remains collected in 1952, with the remarks on the age of the vertebrate bearing beds. Acta Boreal. A Sci. 1953, 4, 32. [Google Scholar]
- Ørvig, T. The Jurassic and Cretaceous of Andøy in Northern Norway. Norges Geol. Unders. 1960, 208, 344–350. [Google Scholar]
- Bose, L.I. Leaf cuticle and other plant microfossils from the Mesozoic rocks of Andøya, Norway. Paleobotanist 1959, 8, 1–7. [Google Scholar]
- Manum, S. A new species of Pseudotorellia Florin from the Jurassic of Andøya, Northern-Norway. Linn. Soc. Lond. Bot. J. 1968, 61, 197–200. [Google Scholar] [CrossRef][Green Version]
- Manum, S.B. Mesozoic Sciadopitys-like leaves with observation on four species from the Jurassic of Andøya, Northern Norway, and emendation of Sciadopityoides Sveshnikova. Rev. Palaeobot. Palynol. 1987, 51, 145–168. [Google Scholar] [CrossRef]
- Manum, S.B.; Bose, M.N.; Vigran, J.O. The Jurassic flora of Andøya, Northern Norway. Rev. Palaeobot. Palynol. 1991, 68, 233–256. [Google Scholar] [CrossRef]
- Birkelund, T.; Thusu, B.; Vigran, J.O. Jurassic-Cretaceous biostratigraphy of Norway, with comments on the British Rasenia cymodoce zone. Palaeontology 1978, 21, 31–63. [Google Scholar]
- Zakharov, V.A.; Surlyk, F.; Dalland, A. Upper Jurassic—Lower Cretaceous Buchia from Andøy, Northern Norway. Norsk Geol. Tidsskr. 1981, 61, 261–269. [Google Scholar]
- Løfaldli, M.; Thusu, B. Micropalaeontological studies of the Upper Jurassic and Lower Cretaceous of Andøya, Northern Norway. Palaeontology 1979, 22, 413–425. [Google Scholar]
- Aarhus, N.; Verdenius, J.; Birkelund, T. Biostratigraphy of a Lower Cretaceous section from Sklinnabanken, Norway, with some comments on the Andøya exposure. Norsk Geol. Tidsskr. 1986, 66, 17–43. [Google Scholar]
- Brönner, M.; Johansen, V.; Baranwal, A.; Črne, B.; Davidsen, H.; Elvebakk, A.; Engvik, T.; Forthun, J.; Gellein, T.; Henningsen, O.; et al. The Ramså Basin, Northern Norway: An Integrated Study; NGU Report No. 2017.027; Geological Survey of Norway: Trondheim, Norway, 2017; p. 290. [Google Scholar]
- Dalland, A. The Mesozoic rocks of Andøy, northern Norway. Nor. Geol. Unders. 1975, 316, 271–287. [Google Scholar]
- Sturt, B.A.; Dalland, A.; Mitchell, J.L. The Age of the Sub Mid-Jurassic Tropical Weathering Profile of Andøya, Northern Norway, and the Implications for the Late Palaeozoic Palaeography in the North Atlantic Region. Geol. Rundsch. 1979, 68, 523–542. [Google Scholar] [CrossRef]
- Dalland, A. Mesozoic Sedimentary Succession at Andøy, Northern Norway, and Relation to Structural Development of the North Atlantic Area. In Geology of the North Atlantic Borderlands; Kerr, J.W., Ferguson, A.J., Eds.; Canadian Society of Petroleum Geologists: Calgary, AB, Canada, 1981; Volume 7, pp. 563–584. [Google Scholar]
- Tyson, R.V. Sedimentary Organic Matter. Organic Facies and Palynofacies; Chapman & Hall: London, UK, 1994; p. 640. [Google Scholar]
- Koppelhus, E.B. Palynology of the Lower Jurassic Rønne Formation on Bornholm, eastern Denmark. Bull. Geol. Soc. Den. 1991, 39, 91–109. [Google Scholar]
- Holstad, T. Palynology of the uppermost Lower to Middle Jurassic strata on Bornholm, Denmark. Bull. Geol. Soc. Den. 1985, 34, 111–132. [Google Scholar]
- Dypkjær, K. Palynological zonation and stratigraphy of the Jurassic section in the Gassum No. 1-borehole, Denmark. Dan. Geol. Unders. Ser. A 1988, 21, 72. [Google Scholar]
- Lund, J.J.; Pedersen, K.R. Palynology of marine Jurassic formations in the Vardekløft ravine, Jameson land, East Greenland. Bull. Geol. Soc. Den. 1985, 33, 371–390. [Google Scholar]
- Riding, J.B.; Thomas, J.E. Dinoflagellate Cysts of the Jurassic System. In A Stratigraphic Index of Dinoflagellate Cysts; British Micropalaeontological Society Publication Series; Powell, A.J., Ed.; Chapman & Hall: London, UK, 1992; pp. 7–97. [Google Scholar]
- Poulsen, N.E.; Riding, J.B. The Jurassic dinoflagellate cyst zonation of Subboreal Northwest Europe. Geol. Surv. Den. Greenl. Bull. 2003, 1, 115–144. [Google Scholar] [CrossRef]
- Ilyina, V.I.; Nikitenko, B.L.; Glinskikh, L.A. Foraminifera and Dinoflagellate Cyst Zonation and Stratigaphy of the Callovian to Volgian Reference Section in the Tyumenskaya Superdeep Well (West Siberia, Russia). In Recent Developments in Applied Biostratigraphy; The Micropalaeontological Society, Special Publications; Powell, A.J., Riding, J.B., Eds.; The Geological Society: London, UK, 2005; Volume 109, pp. 109–144. [Google Scholar]
- Riding, J.B.; Fedorova, V.A.; Ilyina, V.I. Jurassic and lowermost Cretaceous dinoflagellate cyst biostratigraphy of the Russian Platform and northern Siberia, Russia. Am. Assoc. Strat. Palynol. Contr. Ser. 1999, 36, 183. [Google Scholar]
- Poulsen, N.E. Dinoflagellate Cysts from Marine Jurassic Deposits of Denmark and Poland. Am. Assos. Strat. Palynol. Contr. Ser. 1996, 31, 227. [Google Scholar]
- Dybkjær, K. Palynological dating of the Mandal Formation (uppermost Jurassic-lowermost Cretaceous, Norwegian Central Graben) and correlation to organic-rich shales in the Danish sector. Mar. Petrol. Geol. 1998, 15, 495–503. [Google Scholar] [CrossRef]
- Smelror, M.; Below, R. Dinoflagellate biostratigraphy of the Toarcian to Lower Oxfordian (Jurassic) of the Barents Sea Region. In Arctic Geology and Petroleum Potential; NPF Special Publication; Vorren, T., Bergsager, E., Dahl-Stamnes, Ø.A., Holter, E., Johansen, B., Lie, E., Lund, T.B., Eds.; Elsevier: Amsterdam, The Netherlands, 1992; Volume 2, pp. 495–513. [Google Scholar]
- Nøhr-Hansen, H. Dinoflagellate cyst stratigraphy of the Barremian to Albian, Lower Cretaceous, North-East Greenland. Grønlands Geol. Unders. Bull. 1993, 166, 171. [Google Scholar]
- Nøhr-Hansen, H.; Piasecki, S.; Alsen, P. A Cretaceous dinoflagellate cyst zonation for NE Greenland. Geol. Mag. 2019, 157, 1–35. [Google Scholar] [CrossRef]
- Heilmann-Clausen, C. Lower Cretaceous dinoflagellate biostratigraphy in the Danish Central Trough. Dan. Geol. Unders. Ser. A 1987, 17, 89. [Google Scholar]
- Costa, L.I.; Davey, R.J. Dinoflagellate Cysts of the Cretaceous System. In A Stratigraphic Index of Dinoflagellate Cysts; British Micropalaeontological Society Publication Series; Powell, A.J., Ed.; Chapman & Hall: London, UK, 1992; pp. 99–153. [Google Scholar]
- Poulsen, N.E. The dinoflagellate cyst genus Epiplosphaera Klement 1960—A reappraisal. J. Micropalaeont. 1992, 11, 65–72. [Google Scholar] [CrossRef]
- Gorin, G.E.; Steffen, D. Organic facies as a tool for recording eustatic variations in marine fine-grained carbonates—Example of the Berriasian stratotype at Berrias (Ardèche, SE France). Palaeogeogr. Paleoclimatol. Palaeoecol. 1991, 85, 303–320. [Google Scholar] [CrossRef]
- Gregory, W.A.; Hart, G.F. Towards a Predictive Model for the Palynological Response to Sea-Level Changes. Palaios 1992, 7, 3–33. [Google Scholar] [CrossRef]
- Batten, D.J.; Stead, D.T. Palynofacies Analysis and its Stratigraphic Application. In Applied Stratigraphy. Topics in Geobiology; Koutsoukos, E.A.M., Ed.; Springer: Dordrecht, The Netherlands, 2005; Volume 23, pp. 203–226. [Google Scholar]
- Dalseg, T.S.; Nakrem, H.A.; Smelror, M. Dinoflagellate biostratigraphy, palynofacies, depositional environment and sequence stratigraphy of the Agardhfjellet Formation (Upper Jurassic–Lower Cretaceous) in central Spitsbergen (Arctic Norway). Nor. J. Geol. 2016, 96, 119–133. [Google Scholar] [CrossRef][Green Version]
- Sittler, C.; Ollivier-Pierre, M.-F. Palynology and palynofacies analyses: Some essential clues to assess and identify West-European Tertiary depositinal environments in terms of relative high or lowstands. Application to the case of three Eocene and Oligocene sections in France. Bull. Cent. Rech. Explor. Prod. Elf Aquitaine 1994, 18, 475–488. [Google Scholar]
- Embry, A. Transgressive-regressive (T-R) sequence analysis of the Jurassic succession of the Sverdrup Basin, Canadian Arctic Archipelago. Can. J. Earth Sci. 1993, 30, 301–320. [Google Scholar] [CrossRef]
- Embry, A. Sequence Boundaries and Sequence Hierarchies: Problems and Proposals: In Sequence Stratigraphy on the Northwest European Margin; Norwegian Petroleum Society Special Publication; Steel, R., Felt, V.L., Johannesson, E.P., Mathieu, C., Eds.; Elsevier: Amsterdam, The Netherlands, 1995; Volume 5, pp. 1–11. [Google Scholar]
- Smelror, M.; Mørk, M.B.E.; Mørk, A.; Løseth, H.; Weiss, H.M. Middle Jurassic-Lower Cretaceous Transgressive-Regressive Sequences and Facies Distribution off Troms, Northern Norway. In Sedimentary Environments Offshore Norway—Palaeozoic to Recent; Norwegian Petroleum Society Special Publication; Martinsen, O.J., Dreyer, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; Volume 10, pp. 211–232. [Google Scholar]
- Bugge, T.; Elvebakk, G.; Fanavoll, S.; Mangerud, G.; Smelror, M.; Weiss, H.; Gjelberg, J.; Kristensen, S.E.; Nilsen, K. Shallow stratigraphic drilling applied in hydrocarbon exploration of the Nordkapp Basin, Barents Sea. Mar. Pet. Geol. 2002, 19, 13–37. [Google Scholar] [CrossRef]
- Surlyk, F. Sequence stratigraphy of the Jurassic–lowermost Cretaceous of East Greenland. Am. Assoc. Pet. Geol. Bull. 1991, 75, 1468–1488. [Google Scholar]
- Smelror, M.; Mørk, A.; Monteil, E.; Rutledge, D.; Leereveld, H. The Klippfisk Formation—A lithostratigraphic unit of Lower Cretaceous platform carbonates on the Western Barents Shelf. Polar Res. 1998, 17, 181–202. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Borehole Bh-1 | |||||||
|---|---|---|---|---|---|---|---|
| Depth (m) | Lithostrat. | Mar | Ter | Cut | Wod | Blk | T-R Sequence |
| 8.55 | Breisanden Member | 2 | 15 | 3 | 11 | 70 | Berriasian-Kimmeridgian |
| 16.98 | 2 | 11 | 1 | 7 | 79 | ||
| 22.23 | 3 | 12 | 1 | 7 | 77 | ||
| 26.18 | Bonteigen Member | 1 | 8 | 4 | 87 | ||
| 34.80 | 1 | 20 | 5 | 19 | 55 | ||
| 38.09 | 1 | 10 | 1 | 5 | 83 | ||
| 42.60 | 1 | 11 | 7 | 51 | 30 | ||
| 45.86 | 1 | 12 | 3 | 12 | 74 | ||
| 59.44 | 5 | 1 | 10 | 84 | Bajocian | ||
| 66.32 | Kullgrøfta Member | 2 | 23 | 72 | 3 | ||
| 74.40 | 7 | 18 | 73 | 2 | |||
| 79.10 | 21 | 10 | 64 | 5 | |||
| 85.01 | Hestberget Member | 23 | 12 | 59 | 6 | ||
| 89.65 | 19 | 9 | 67 | 5 | |||
| 93.10 | 8 | 2 | 10 | 80 | |||
| 104.10 | 5 | 1 | 9 | 85 | |||
| 106.12 | 2 | 2 | 91 | 5 | |||
| 113.25 | Basal conglomerate | 1 | 1 | 98 | |||
| 116.87 | 2 | 1 | 97 | ||||
| 121.78 | 1 | 1 | 98 | ||||
| 132.62 | 1 | 3 | 1 | 96 | |||
| 146.28 | 2 | 1 | 97 | ||||
| 155.75 | 2 | 1 | 2 | 94 | |||
| 157.07 | |||||||
| 166.80 | 2 | 2 | 3 | 93 | |||
| 193.25 | |||||||
| Borehole Bh-2 | |||||||
|---|---|---|---|---|---|---|---|
| Depth (m) | Litostrat. | Mar | Ter | Cut | Wod | Blk | T-R Seq. |
| 25.00 | Skarstein Formation | 3 | 5 | 2 | 7 | 83 | Barremian-Aptian |
| 39.50 | 6 | 5 | 5 | 11 | 73 | ||
| 68.35 | 3 | 7 | 5 | 40 | 45 | ||
| 72.60 | 4 | 5 | 2 | 8 | 81 | ||
| 86.20 | 1 | 9 | 4 | 35 | 51 | ||
| 89.70 | 4 | 8 | 5 | 31 | 52 | ||
| 93.75 | 5 | 19 | 5 | 21 | 50 | ||
| 97.15 | 5 | 16 | 3 | 15 | 61 | ||
| 100.80 | 2 | 14 | 4 | 21 | 59 | ||
| 102.05 | 4 | 16 | 2 | 25 | 53 | ||
| 110.05 | 7 | 15 | 3 | 38 | 37 | ||
| 115.10 | Nybrua Formation | 10 | 6 | 1 | 5 | 78 | Valanginian- Barremian |
| 116.50 | 5 | 3 | 4 | 88 | |||
| 119.45 | 5 | 2 | 3 | 90 | |||
| 121.40 | 1 | 5 | 94 | ||||
| 123.65 | 1 | 99 | |||||
| 125.95 | Dragneset Formation | 1 | 9 | 6 | 39 | 45 | Kimmeridgian-Berriasian |
| 127.80 | 3 | 8 | 3 | 33 | 53 | ||
| 142.12 | 1 | 9 | 5 | 39 | 46 | ||
| 148.25 | 1 | 10 | 5 | 33 | 51 | ||
| 153.20 | 1 | 10 | 7 | 47 | 35 | ||
| 159.80 | 2 | 4 | 3 | 91 | |||
| 166.40 | 2 | 5 | 5 | 88 | |||
| 171.98 | 1 | 11 | 5 | 47 | 36 | ||
| 177.05 | 1 | 12 | 6 | 49 | 32 | ||
| 182.70 | 1 | 9 | 7 | 50 | 33 | ||
| 188.35 | 2 | 10 | 5 | 43 | 40 | ||
| 196.95 | 2 | 11 | 3 | 46 | 38 | ||
| 199.30 | 1 | 10 | 5 | 46 | 38 | ||
| 200.05 | 8 | 9 | 61 | 32 | |||
| 201.70 | 1 | 99 | B | ||||
| 202.13 | 3 | 1 | 1 | 95 | |||
| Borehole Bh-3 | |||||||
|---|---|---|---|---|---|---|---|
| Depth (m) | Lithostrat. | Mar | Ter | Cut | Wod | Blk | T-R Seq. |
| 16.60 | Taumhølet Member | 1 | 25 | 2 | 22 | 50 | Kimmeridgian-Berriasian |
| 22.20 | 1 | 9 | 4 | 39 | 47 | ||
| 31.60 | 1 | 15 | 5 | 49 | 30 | ||
| 36.60 | 1 | 10 | 5 | 41 | 53 | ||
| 44.45 | 1 | 14 | 2 | 14 | 69 | ||
| 48.61 | 1 | 20 | 3 | 25 | 51 | ||
| 57.50 | 1 | 19 | 6 | 33 | 41 | ||
| 63.38 | 2 | 15 | 1 | 12 | 70 | ||
| 68.70 | 1 | 21 | 5 | 38 | 35 | ||
| 76.75 | 2 | 24 | 4 | 31 | 39 | ||
| 80.81 | 2 | 30 | 2 | 31 | 35 | ||
| 91.00 | 1 | 15 | 7 | 35 | 42 | ||
| 99.10 | 1 | 12 | 6 | 37 | 44 | ||
| 100.54 | 1 | 35 | 6 | 23 | 35 | ||
| 105.40 | 1 | 15 | 1 | 8 | 75 | ||
| 114.00 | Breisanden Member | 1 | 21 | 5 | 18 | 55 | |
| 120.12 | 1 | 19 | 8 | 72 | |||
| 131.16 | 1 | 9 | 1 | 9 | 80 | ||
| 139.78 | 1 | 5 | 1 | 8 | 85 | ||
| 142.20 | 2 | 32 | 2 | 23 | 41 | ||
| 148.00 | 2 | 23 | 3 | 27 | 45 | ||
| 152.25 | Bonteigen Member | 1 | 4 | 1 | 5 | 89 | |
| 164.03 | 1 | 2 | 2 | 95 | |||
| 169.84 | 1 | 5 | 2 | 92 | |||
| 173.80 | 3 | 17 | 4 | 21 | 35 | ||
| 176.00 | 1 | 6 | 1 | 4 | 88 | ||
| 176.05 | 8 | 10 | 1 | 11 | 70 | ||
| 181.45 | 3 | 1 | 16 | 80 | Bajocian | ||
| 186.00 | 5 | 5 | 61 | 29 | |||
| 193.86 | Kullgrøfta Member | 5 | 11 | 81 | 3 | ||
| 195.00 | 6 | 10 | 49 | 35 | |||
| 200.72 | 2 | 1 | 7 | 90 | |||
| 203.45 | 8 | 9 | 52 | 31 | |||
| 207.75 | Hestberget Member | 4 | 5 | 41 | 50 | ||
| 209.31 | 3 | 1 | 14 | 82 | |||
| 212.85 | B | 2 | 6 | 49 | 43 | B | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smelror, M. Palynostratigraphy, Palynofacies, T-R Cycles and Paleoenvironments in the Middle Jurassic–Early Cretaceous Ramså Basin, Andøya, Northern Norway. Geosciences 2021, 11, 354. https://doi.org/10.3390/geosciences11090354
Smelror M. Palynostratigraphy, Palynofacies, T-R Cycles and Paleoenvironments in the Middle Jurassic–Early Cretaceous Ramså Basin, Andøya, Northern Norway. Geosciences. 2021; 11(9):354. https://doi.org/10.3390/geosciences11090354
Chicago/Turabian StyleSmelror, Morten. 2021. "Palynostratigraphy, Palynofacies, T-R Cycles and Paleoenvironments in the Middle Jurassic–Early Cretaceous Ramså Basin, Andøya, Northern Norway" Geosciences 11, no. 9: 354. https://doi.org/10.3390/geosciences11090354
APA StyleSmelror, M. (2021). Palynostratigraphy, Palynofacies, T-R Cycles and Paleoenvironments in the Middle Jurassic–Early Cretaceous Ramså Basin, Andøya, Northern Norway. Geosciences, 11(9), 354. https://doi.org/10.3390/geosciences11090354