
Benthic Foraminiferal Response to Sedimentary Processes in a Prodeltaic Environment: The Gulf of Patti Case Study (Southeastern Tyrrhenian Sea)

 ,
, 

Abstract
:1. Introduction
Environmental Setting
2. Materials and Methods
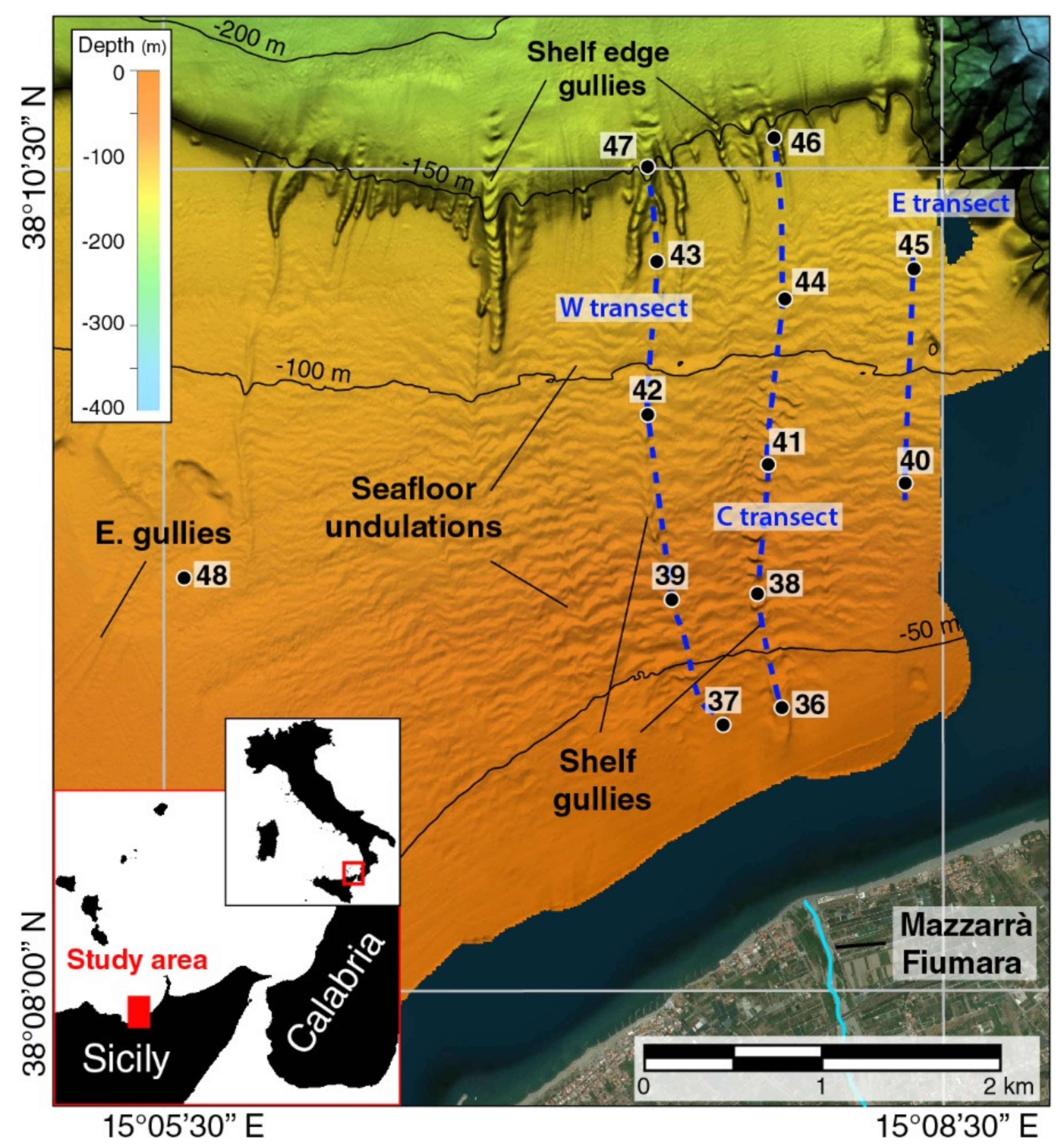
2.1. Sampling Area and Strategy
2.2. Grain-Size Analysis
2.3. Foraminiferal Assemblages
2.4. Statistical Analyses
3. Results
3.1. Grain Size Analysis
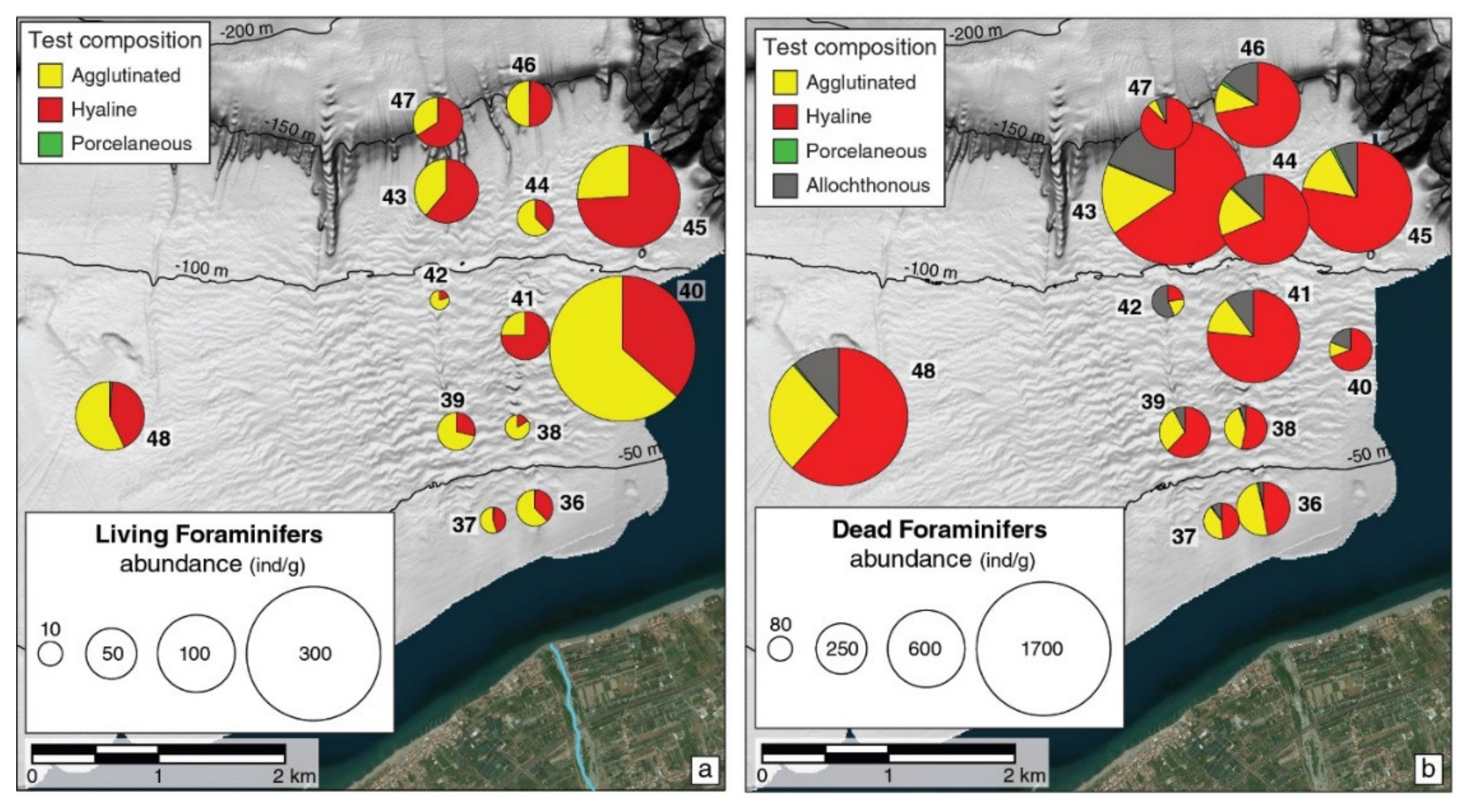
3.2. General Features of Living Foraminiferal Assemblages
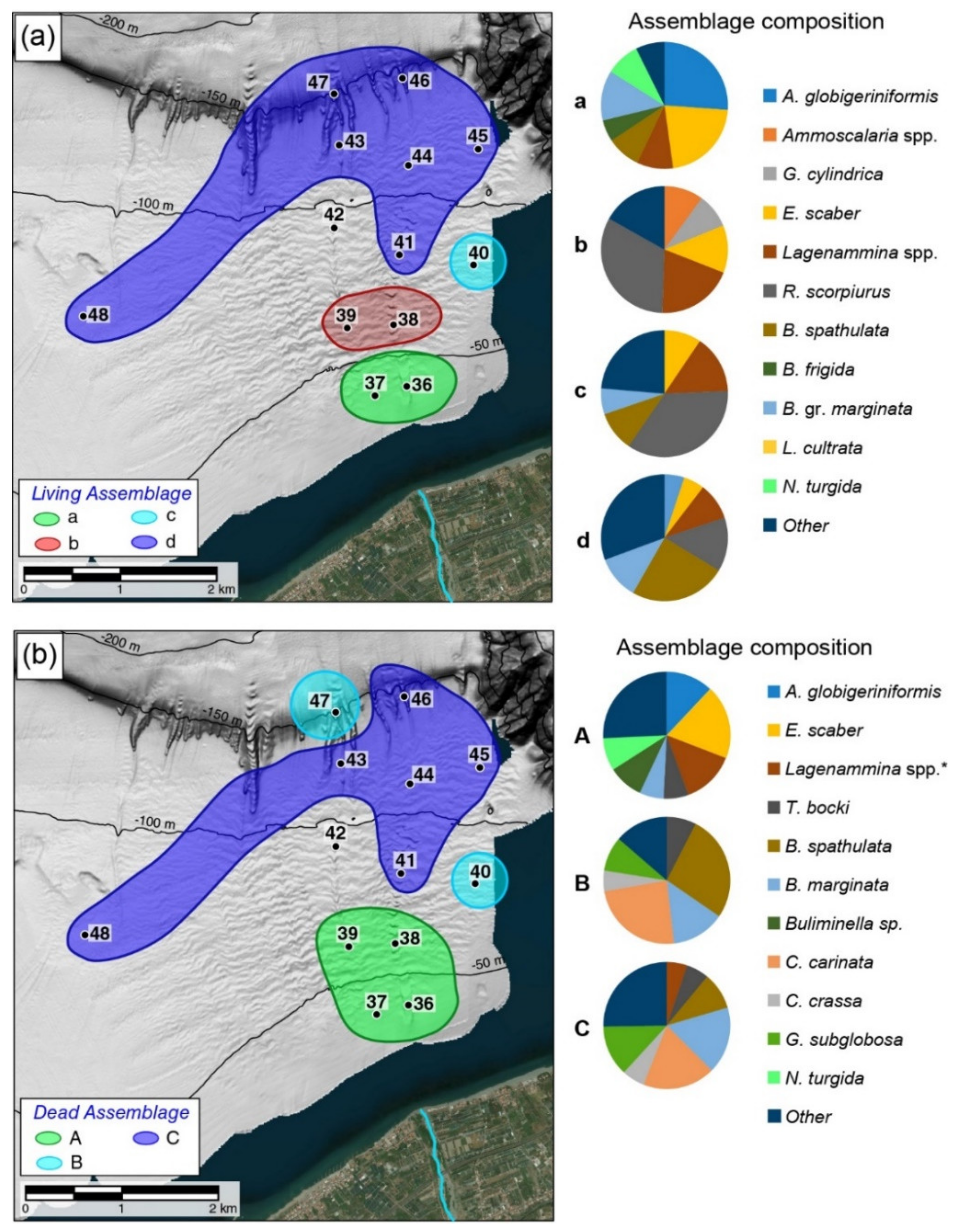
3.3. Living Foraminiferal Assemblages Distribution
3.4. General Features of Dead Foraminiferal Assemblages
3.5. Dead Foraminiferal Assemblages Distribution
4. Discussion
4.1. Response of Living Foraminiferal Assemblages to Sedimentary Disturbance
4.2. Dead Foraminiferal Assemblages and Allochthonous Taxa
4.3. Comparison with Other Mediterranean Prodeltas
5. Conclusions
- In highly energetic hydrodynamic settings, physical disturbance related to sediment transport processes and food supply represents the most significant environmental factors controlling the foraminiferal assemblages in faunal density, biodiversity, and taxonomics compositions.
- The dominance of opportunistic agglutinated taxa (mainly Reophax spp., Eggerelloides spp.) associated with hyaline eutrophic species (Bolivina spp., Bulimina spp., N. turgida) represents a distinctive character of the living benthic foraminiferal community in this sector of the southern Tyrrhenian margin. A clear decrease of these taxa is recorded in the marginal areas of the prodelta, confirming their correlation with river input. At depth >100 m, more oligotrophic conditions due to greater distance from the coastline favor developing typical deep-sea assemblages characterized by the occurrence of shallow infaunal species. The dead assemblage confirms that the distribution is controlled by sedimentary and organic matter input along a depth gradient.
- Compared to other sites of the Tyrrhenian margin characterized by similar geological, hydrodynamic and sedimentary settings confirms the strong correlation between the assemblages and the organic matter enrichment and the concurrent physical disturbance by fluvial flows.
- The study provides insights for paleoenvironmental reconstructions in similar environments during the Holocene.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mopper, K.; Degens, E.T. Organic carbon in the ocean: Nature and cycling. Glob. Carb. Cycle 1979, 13, 293–316. [Google Scholar]
- Bauer, J.E.; Druffel, E.R. Ocean margins as a significant source of organic matter to the deep open ocean. Nature 1998, 392, 482. [Google Scholar]
- Lohrenz, S.E.; Dagg, M.J.; Whitledge, T.E. Enhanced primary production at the plume/oceanic interface of the Mississippi River. Cont. Shelf Res. 1990, 10, 639–664. [Google Scholar]
- McKee, B.A.; Aller, R.C.; Allison, M.A.; Bianchi, T.S.; Kineke, G.C. Transport and transformation of dissolved and particulate materials on continental margins influenced by major rivers: Benthic boundary layer and seabed processes. Cont. Shelf Res. 2004, 24, 899–926. [Google Scholar]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006; p. 426. [Google Scholar]
- Lohrenz, S.E.; Fahnenstiel, G.L.; Redalje, D.G.; Lang, G.A.; Chen, X.; Dagg, M.J. Variations in primary production of northern Gulf of Mexico continental shelf waters linked to nutrient inputs from the Mississippi River. Mar. Ecol. Prog. Ser. 1997, 155, 45–54. [Google Scholar]
- Dagg, M.J.; Breed, G.A. Biological effects of Mississippi River nitrogen on the northern Gulf of Mexico—A review and synthesis. J. Mar. Syst. 2003, 43, 133–152. [Google Scholar]
- Danovaro, R.; Gambi, C.; Manini, E.; Fabiano, M. Meiofauna response to a dynamic river plume front. Mar. Biol. 2000, 137, 359–370. [Google Scholar]
- Salen-Picard, C.; Arlhac, D.; Alliot, E. Responses of a Mediterranean soft bottom community to short-term (1993–1996) hydrological changes in the Rhone river. Mar. Environ. Res. 2003, 55, 409–427. [Google Scholar]
- Goineau, A.; Fontanier, C.; Jorissen, F.; Buscail, R.; Kerhervé, P.; Cathalot, C.; Pruski, A.M.; Lantoine, F.; Bourgeois, S.; Metzger, E.; et al. Temporal variability of live (stained) benthic foraminiferal faunas in a river-dominated shelf–Faunal response to rapid changes of the river influence (Rhône prodelta, NW Mediterranean). Biogeosciences 2012, 9, 1367–1388. [Google Scholar]
- Gooday, A.; Levin, L.A.; Linke, P.; Heeger, T. The role of benthic foraminifera in deep-sea food webs and carbon cycling. In Deep-Sea Food Chains and the Global Carbon Cycle; Rowe, G.T., Patiente, V., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992; pp. 63–69. [Google Scholar]
- Moodley, L.; van der Zwaan, G.J.; Herman, P.M.J.; Kempers, L.; van Breugel, P. Differential response of benthic meiofauna to anoxia with special reference to the Foraminifera (Protista: Sarcodina). Mar. Ecol. Prog. Ser. 1997, 158, 151–163. [Google Scholar]
- Debenay, J.P.; Fernandez, J.M. Benthic foraminifera records of complex anthropogenic environmental changes combined with geochemical data in a tropical bay of New Caledonia (SW Pacific). Mar. Pollut. Bull. 2009, 59, 311–322. [Google Scholar] [PubMed]
- Danovaro, R. Methods for the Study of Deep-Sea Sediments, Their Functioning and Biodiversity; CRC Press: Boca Raton, FL, USA, 2009; p. 428. [Google Scholar]
- Balsamo, M.; Semprucci, F.; Frontalini, F.; Coccioni, R. Meiofauna as a tool for marine ecosystem biomonitoring marine ecosystems. In Marine Ecosystems; Antonio, C., Ed.; In Tech: Rijeka, Hrvatska, 2012; pp. 77–710. [Google Scholar]
- Snider, L.J.; Burnett, B.R.; Hessler, R.R. The composition and distribution of meiofauna and nanobiota in a central North Pacific deep-sea are. Deep Sea Res. 1984, 31, 1225–1249. [Google Scholar]
- Van Der Zwaan, G.J.; Duijnstee, I.A.P.; Den Dulk, M.; Ernst, S.R.; Jannink, N.T.; Kouwenhoven, T.J. Benthic foraminifers: Proxies or problems? A review of paleocological concepts. Earth Sc. Rev. 1999, 46, 213–236. [Google Scholar]
- Cearreta, A.; Benito, X.; Ibáñez, C.; Trobajo, R.; Giosan, L. Holocene palaeoenvironmental evolution of the Ebro Delta (Western Mediterranean): Evidence for an early construction based on the benthic foraminiferal record. Holocene 2016, 26, 1438–1456. [Google Scholar] [CrossRef]
- Schmiedl, G.; de Bovée, F.; Buscail, R.; Charrière, B.; Hemleben, C.; Medernach, L.; Picon, P. Trophic control of benthic foraminiferal abundance and microhabitat in the bathyal Gulf of Lions, western Mediterranean Sea. Mar. Micropaleont. 2000, 40, 167–188. [Google Scholar]
- Hyams-Kaphzan, O.; Almogi-Labin, A.; Sivan, D.; Benjamini, C. Benthic foraminifera assemblage change along the southeastern Mediterranean inner shelf due to fall-off of Nile-derived siliciclastics. Neues Jahr. Geol. Paläontol. Abh. 2008, 248, 315–344. [Google Scholar]
- Jorissen, F.J. Benthic foraminifera from the Adriatic Sea; principles of phenotypic variations. Utrecht Micropaleontol. Bull. 1988, 37, 1–176. [Google Scholar]
- Asioli, A.; Trincardi, F.; Lowe, J.J.; Ariztegui, D.; Langone, L.; Oldfield, F. Sub-millennial scale climatic oscillations in the central Adriatic during the Lateglacial: Palaeoceanographic implications. Quat. Sci. Rev. 2001, 20, 1201–1221. [Google Scholar]
- Amorosi, A.; Colalongo, M.L.; Fiorini, F.; Fusco, F.; Pasini, G.; Vaiani, S.C.; Sarti, G. Palaeogeographic and palaeoclimatic evolution of the Po Plain from 150-ky core records. Glob. Planet. Chang. 2004, 40, 55–78. [Google Scholar]
- Di Bella, L.; Bellotti, P.; Milli, S. The role of foraminifera as indicators of the Late Pleistocene-Holocene palaeoclimatic fluctuations on the deltaic environment: The example of Tiber delta succession (Tyrrhenian margin, Italy). Quat. Int. 2013, 303, 191–209. [Google Scholar]
- Rossi, V.; Vaiani, S.C. Benthic foraminiferal evidence of sediment supply changes and fluvial drainage reorganization in Holocene deposits of the Po Delta, Italy. Mar. Micropaleontol. 2008, 69, 106–118. [Google Scholar]
- Milli, S.; D’Ambrogi, C.; Bellotti, P.; Calderoni, G.; Carboni, M.G.; Celant, A.; Di Bella, L.; Di Rita, F.; Frezza, V.; Magri, D.; et al. The transition from wave-dominated estuary to wave-dominated delta: The Late Quaternary stratigraphic architecture of Tiber River deltaic succession (Italy). Sediment. Geol. 2013, 284–285, 159–180. [Google Scholar]
- Mendes, F.J.; Lobo, L.M.; Fernández-Salas, N.; López-González, P.; Bárcenas, J.; Schönfeld, J.; Ferreira, O. Multi-proxy evidence of rainfall variability recorded in subaqueous deltaic deposits off the Adra River, southeast Iberian Peninsula Estuarine Coast. Shelf Sci. 2015, 167, 300–312. [Google Scholar]
- Manini, E.; Danovaro, R.; Fabiano, M. 2002. Benthic-pelagic coupling in frontal system areas of the northern Adriatic Sea: Analysis of the carbon budgets. Chem. Ecol. 2015, 18, 155–160. [Google Scholar]
- Jorissen, F.J.; Barmawidjaja, D.M.; Puskaric, S.; Van der Zwaan, G.J. Vertical distribution of benthic foraminifera in the northern Adriatic Sea: The relation with the organic flux. Mar. Micropaleontol. 1992, 19, 131–146. [Google Scholar]
- McAllen, R.; Davenport, J.; Bredendieck, K.; Dunne, D. Seasonal structuring of a benthic community exposed to regular hypoxic events. J. Experim. Mar. Biol. Ecol. 2009, 368, 67–74. [Google Scholar]
- Eichler, P.B.; McGann, M.; Rodrigues, A.R.; Mendonça, A.; Amorim, A.; Bonetti, C.; Cordeiro de Farias, C.; Mello e Sousa, S.H.; Vital, H.; Gomes, M.P. The occurrence of the invasive foraminifera Trochammina hadai Uchio in Flamengo Inlet, Ubatuba, São Paulo State, Brazil. Micropaleontology 2018, 64, 391–402. [Google Scholar]
- Goineau, A.; Fontanier, C.; Jorissen, F.J.; Lansard, B.; Buscail, R.; Mouret, A.; Kerhervé, P.; Zaragosi, S.; Ernoult, E.; Artero, C.; et al. Live (stained) benthic foraminifera from the Rhône prodelta (Gulf of Lion, NW Mediterranean): Environmental controls on a river-dominated shelf. J. Sea Res. 2011, 65, 58–75. [Google Scholar]
- Yamashita, C.; Omachi, C.; Aoki Santarosa, A.C.; Sayuri Iwai, F.; Dias Araujo, B.; Trevisan Disaró, S.; Martins, M.V.A.; Vicente, T.M.; Taniguchi, N.; Burone, L.; et al. Living benthic foraminifera of Santos continental shelf, southeastern Brazilian continental margin (SW Atlantic): Chlorophyll-a and particulate organic matter approach. J. Sediment. Environ. 2019, 5, 17–34. [Google Scholar]
- Di Bella, L.; Pierdomenico, M.; Porretta, R.; Chiocci, F.L.; Martorelli, E. Living and dead foraminiferal assemblages from an active submarine canyon and surrounding sectors: The Gioia Canyon system (Tyrrhenian Sea, Southern Italy). Deep Sea Res. Part I 2017, 123, 129–146. [Google Scholar]
- Pierdomenico, M.; Martorelli, E.; Dominguez-Carrió, C.; Gili, J.M.; Chiocci, F.L. Seafloor characterization and benthic megafaunal distribution of an active submarine canyon and surrounding sectors: The case of Gioia Canyon (Southern Tyrrhenian Sea). J. Mar. Syst. 2016, 157, 101–117. [Google Scholar]
- Pierdomenico, M.; Cardone, F.; Carluccio, A.; Casalbore, D.; Chiocci, F.; Maiorano, P.; D’Onghia, G. Megafauna distribution along active submarine canyons of the central Mediterranean: Relationships with environmental variables. Prog. Ocean. 2019, 171, 49–69. [Google Scholar]
- Martins, M.V.; Jouanneau, J.-M.; Weber, O.; Rocha, F. Tracing the late Holocene evolution of the NW Iberian upwelling system. Mar. Micropaleont. 2006, 59, 35–55. [Google Scholar]
- Bartels-Jonsdottir, H.B.; Voelker, A.H.L.; Knudsen, L.; Abrantes, F. Twentieth century warming and hydrographical changes in the Tagus prodelta, eastern North Atlantic. Holocene 2009, 19, 369–380. [Google Scholar]
- Mendes, I.; Dias, J.A.; Schonfeld, J.; Ferreira, O. Distribution of living benthic foraminifera on the northern Gulf of Cadiz continental shelf. J. Foraminifer. Res. 2012, 42, 18–38. [Google Scholar]
- Casalbore, D.; Ridente, D.; Bosman, A.; Chiocci, F.L. Depositional and erosional bedforms in Late Pleistocene-Holocene pro-delta deposits of the Gulf of Patti (southern Tyrrhenian margin, Italy). Mar. Geol. 2017, 385, 216–227. [Google Scholar]
- Istituto Idrografico della Marina. Atlante delle Correnti Superficiali dei Mari d’Italia; Istituto Idrografico della Marina Publ.: Genova, Italy, 1982. [Google Scholar]
- Ferranti, L.; Antonioli, F.; Mauz, B.; Amorosi, A.; Dai Pra, G.; Mastronuzzi, G.; Monaco, C.; Orrù, P.; Pappalardo, M.; Radtke, U.; et al. Markers of the last interglacial sea-level high stand along the coast of Italy: Tectonic implications. Quat. Int. 2006, 145, 30–54. [Google Scholar]
- Sulli, A.; Lo Presti, V.; Morticelli, M.G.; Antonioli, F. Vertical movements in NE Sicily and its offshore: Outcome of tectonic uplift during the last 125 ky. Quat. Int. 2013, 288, 168–182. [Google Scholar]
- CPTI Working Group. Catalogo Parametrico dei Terremoti Italiani, Version 2004. (CPTI04); INGV: Bologna, Italy, 2004; Available online: http://emidius.mi.ingv.it/CPTI (accessed on 1 March 2021).
- Casalbore, D.; Chiocci, F.L.; Mugnozza, G.S.; Tommasi, P.; Sposato, A. Flash-flood hyperpycnal flows generating shallow-water landslides at Fiumara mouths in Western Messina Strait (Italy). Mar. Geophys. Res. 2011, 32, 257. [Google Scholar]
- Kostic, S.; Casalbore, D.; Chiocci, F.; Lang, J.; Winsemann, J. Role of upper-flow-regime bedforms emplaced by sediment gravity flows in the evolution of deltas. J. Mar. Sci. Eng. 2019, 7, 5. [Google Scholar]
- Folk, R.L.; Ward, W.M. Brazos River bar: A study in the significance of grain size parameters. J. Sediment. Petrol. 1957, 27, 3–26. [Google Scholar]
- Folk, R.L. The distinction between grain size and mineral composition in sedimentary-rock nomenclature. J. Geol. 1954, 62, 344–359. [Google Scholar]
- Folk, R.L. Petrology of Sedimentary Rocks; Hemphill Publishing Co.: Austin, TX, USA, 1974; p. 182. [Google Scholar]
- Poppe, L.; Eliason, A. A Visual Basic program to plot sediment grain-size data on ternary diagrams. Comp. Geosci. 2008, 34, 561–565. [Google Scholar]
- Schönfeld, J.; Alve, E.; Geslin, E.; Jorissen, F.; Korsun, S.; Spezzaferri, S. Members of The Fobimo. The Fobimo (FOraminiferal BIo-MOnitoring) initiative—Towards a standardized protocol for soft-bottom benthic foraminiferal monitoring studies. Mar. Micropaleontol. 2012, 94–95, 1–13. [Google Scholar]
- Mojtahid, M.; Jorissen, F.; Lansard, B.; Fontanier, C.; Bombled, B.; Rabouille, C. Spatial distribution of live benthic foraminifera in the Rhône prodelta: Faunal response to a continental–marine organic matter gradient. Mar. Micropaleontol. 2009, 70, 177–200. [Google Scholar]
- Benito, X.; Trobajo, R.; Cearreta, A.; Ibanez, C. Benthic foraminifera as indicators of habitat in a Mediterranean delta: Implications for ecological and palaeoenvironmental studies Estuar. Coast. Shelf Sci. 2016, 180, 97–113. [Google Scholar]
- Di Bella, L.; Sabbatini, A.; Carugati, L.; Lo Martire, M.; Luna, G.M.; Pierdomenico, M.; Danovaro, R.A.; Negri, A. Living foraminiferal assemblages in two submarine canyons (Polcevera and Bisagno) of the Ligurian basin (Mediterranean Sea). Prog. Oceanogr. 2019, 173, 114–133. [Google Scholar]
- Walton, W.R. Tecniques for recognition of living foraminifera. Contrib. Cushman Found. Foraminifer. Res. 3, 56–60. Found. Foraminifer. Res. 1952, 3, 56–60. [Google Scholar]
- Lutze, G.F.; Altenbach, A. Technik und Signifikanz der Lebendfarbung benthischer Foraminiferen mit Bengalrot. Geol. Jahrb. A 1991, 128, 251–265. [Google Scholar]
- Bernhard, J.M. Distinguishing live from dead foraminifera: Methods review and proper applications. Micropaleontology 2000, 46, 38–46. [Google Scholar]
- Scott, D.B.; Medioli, F.S.; Schafer, C.T. Monitoring of Coastal Environments Using Foraminifera and Thecamoebian Indicators; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Bernhard, J.M. Postmortem vital staining in benthic foraminifera: Duration and importance in population and distributional studies. J. Foraminifer. Res. 1988, 18, 143–146. [Google Scholar]
- Hannah, F.; Rogerson, A. The temporal and spatial distribution of foraminiferans in marine benthic sediments of the Clyde Sea, Scotland. Estuar. Coast. Shelf Sci. 1997, 44, 377–383. [Google Scholar]
- Murray, J.W.; Bowser, S.S. Mortality, protoplasm decay rate, and reliability of staining techniques to recognize ‘living’ foraminifera: A review. J. Foraminifer. Res. 2000, 30, 66–77. [Google Scholar]
- Fontanier, C.; Jorissen, F.J.; Licari, L.; Alexandre, A.; Anschutz, P.; Carbonel, P. Live benthic foraminiferal faunas from the Bay of Biscay: Faunal density, composition, and microhabitats. Deep Sea Res. I 2002, 49, 751–785. [Google Scholar]
- Frontalini, F.; Semprucci, F.; Di Bella, L.; Caruso, A.; Cosentino, C.; Maccotta, A.; Scopelliti, G.; Sbrocca, C.; Bucci, C.; Balsamo, M.; et al. The Response of Cultured Meiofaunal and Benthic Foraminiferal Communities to Lead Exposure: Results from Mesocosm Experiments. Environ. Toxicol. Chem. 2018, 37, 2439–2447. [Google Scholar] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The relationship between the number of species and the number of individuals in random samples of an animal population. J. Anim. Ecol. 1943, 12, 42–58. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Jorissen, F.J.; de Stigter, H.C.; Widmark, J.G.V. A conceptual model explaining benthic foraminiferal microhabitats. Mar. Micropaleontol. 1995, 26, 3–15. [Google Scholar]
- Loeblich, R.; Tappan, H. Foraminiferal Genera and Their Classification; Van Nostrand Reinhold Company: New York, NY, USA, 1987; p. 970. [Google Scholar]
- Cimerman, F.; Langer, M.R. Mediterranean Foraminifera. Acad. Scient. Artium Sloven. 1991, 30, 1–118. [Google Scholar]
- Sgarrella, F.; Moncharmont Zei, M. Benthic foraminifera of the Gulf of Naples (Italy): Systematics and autoecology. Boll. Soc. Paleontol. Ital. 1993, 32, 145–264. [Google Scholar]
- Koho, K.A.; Kouwenhoven, T.J.; de Stigter, H.C.; van der Zwaan, G.J. Benthic foraminifera in the Nazaré Canyon, Portuguese continental margin: Sedimentary environments and disturbance. Mar. Micropaleontol. 2007, 66, 27–51. [Google Scholar]
- Sen Gupta, B.; Lobegeier, M.; Smith, L. Foraminiferal Communites of Bathyal Hydrocarbon Seeps, Northen Gulf of Mexico: A taxonomic, Ecologic and Geologic Study; Louisiana State University: Baton Rouge, LA, USA, 2009; p. 213. [Google Scholar]
- Milker, Y.; Schmiedl, G. A taxonomic guide to modern benthic shelf foraminifera of the western Mediterranean Sea. Palaeontol. Electron. 2012, 15, 134. [Google Scholar]
- Fatela, F.; Taborda, R. Confidence limits of species proportions in microfossil assemblages. Mar. Micropaleontol. 2002, 45, 169–174. [Google Scholar]
- De Stigter, H.C.; van der Zwaan, G.J.; Langone, L. Differential rates of benthic foraminiferal test production in surface and subsurface sediment habitats in the southern Adriatic Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1999, 149, 67–88. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T. Paleontological Data Analysis. Blackwell Publishing: Oxford, UK, 2006. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T. Paleontological Data Analysis. Blackwell Publishing: Oxford, UK, 2008. [Google Scholar]
- Langer, M.R. Epiphytic foraminifera. Mar. Micropaleontol. 1993, 20, 235–265. [Google Scholar]
- Donnici, S.; Barbero, R.S. The benthic foraminiferal communities of the northern Adriatic continental shelf. Mar. Micropaleontol. 2002, 44, 93–123. [Google Scholar]
- Frezza, V.; Carboni, M.G.; Matteucci, R. Recent foraminiferal assemblages near Ponza Island (Central Tyrrhenian Sea, Italy). Boll. Soc. Paleontol. Ital. 2005, 44, 155–173. [Google Scholar]
- Frezza, V.; Carboni, M.G. Distribution of recent foraminiferal assemblages nearthe Ombrone River mouth (Northern Tyrrhenian Sea, Italy). Rev. Micropaleontol. 2009, 52, 43–66. [Google Scholar]
- Hess, S.; Kuhnt, W. Deep-sea benthic foraminiferal recolonization of the 1991 Mt. Pinatubo ash layer in the South China Sea. Mar. Micropaleontol. 1996, 28, 171–197. [Google Scholar]
- Mc Gann, M. Historical and modern distributions of benthic foraminifers on the continental shelf of Monterey Bay, California. Mar. Geol. 2002, 181, 115–156. [Google Scholar]
- Dessandier, P.A.; Bonnin, J.; Kim, J.H.; Bichon, S.; Grémare, A.; Deflandre, B.; de Stigter, H.; Malaizé, B. Lateral and vertical distributions of living benthic foraminifera off the Douro River (western Iberian margin): Impact of the organic matter quality. Mar. Micropaleontol. 2015, 120, 31–45. [Google Scholar]
- Di Bella, L.; Ingrassia, M.; Frezza, V.; Chiocci, F.L.; Martorelli, E. The response of benthic meiofauna to hydrothermal emissions in the Pontine Archipelago, Tyrrhenian Sea (central Mediterranean Basin). J. Mar. Syst. 2016, 164, 53–66. [Google Scholar]
- Di Bella, L.; Ingrassia, M.; Frezza, V.; Chiocci, F.L.; Pecci, R.; Bedini, R.; Martorelli, E. Spiculosiphon oceana (Foraminifera) a new bio-indicator of acidic environments related to fluid emissions of the Zannone Hydrothermal Field (central Tyrrhenian Sea). Mar. Environ. Res. 2018, 136, 89–98. [Google Scholar] [PubMed]
- Romano, E.; Bergamin, L.; Pierfranceschi, G.; Provenzani, C.; Marassich, A. The distribution of benthic foraminifera in Bel Torrente submarine cave (Sardinia, Italy) and their environmental significance. Mar. Environ. Res. 2018, 133, 114–127. [Google Scholar] [PubMed]
- Romano, E.; Bergamin, L.; Di Bella, L.; Frezza, V.; Marassich, A.; Pierfranceschi, G.; Provenzani, C. Benthic foraminifera as proxies of marine influence in the Orosei marine caves (Sardinia, Italy). Aquat. Conserv: Mar. Freshw Ecosyst. 2020, 30, 701–716. [Google Scholar]
- Dias, B.B.; Hart, M.B.; Smart, C.W.; Hall-Spencer, J.M. Modern seawater acidification: The response of foraminifera to high-CO2 conditions in the Mediterranean Sea. J. Geol. Soc. 2010, 167, 843–846. [Google Scholar]
- Panieri, G.; Gamberi, F.; Marani, M.; Barbieri, R. Benthic foraminifera from a recent, shallow-water hydrothermal environment in the Aeolian Arc (Tyrrhenian Sea). Mar. Geol. 2005, 218, 207–229. [Google Scholar]
- Lei, Y.L.; Li, T.G.; Bi, H.; Cui, W.L.; Song, W.P.; Li, J.Y.; Li, C.C. Responses of benthic foraminifera to the 2011 oil spill in the Bohai Sea, PR China. Mar. Pollut. Bull. 2015, 96, 245–260. [Google Scholar]
- Diz, P.; Francés, G. Distribution of live benthic foraminifera in the Ria de Vigo (NW Spain). Mar. Micropaleontol. 2008, 66, 165–191. [Google Scholar]
- Alve, E.; Goldstein, S.T. Dispersal, survival and delayed growth of benthic foraminiferal propagules. J. Sea Res. 2010, 63, 36–51. [Google Scholar]
- Nardelli, M.P.; Jorissen, F.J.; Pusceddu, A.; Morigi, C.; Dell’Anno, A.; Danovaro, R.; De Stigter, H.C.; Negri, A. Living benthic foraminiferal assemblages along a latitudinal transect at 1000 m depth off the Portuguese margin. Micropaleontology 2010, 56, 323–344. [Google Scholar]
- Mendes, I.; Gonzalez, R.; Dias, J.M.A.; Lobo, P.; Martins, V. Factors influencing recent benthic foraminifera distribution on the Guadiana shelf (southwestern Iberia). Mar. Micropaleontol. 2004, 51, 171–192. [Google Scholar]
- De Rijk, S.; Jorissen, F.J.; Rohling, E.J.; Troelstra, S.R. Organic flux on bathymetric zonation of Mediterranean benthic foraminifera. Mar. Micropaleontol. 2000, 40, 151–166. [Google Scholar]
- Frezza, V.; Pignatti, J.; Matteucci, R. Benthic foraminiferal biofacies in temperate carbonate sediment in the western pontine archipelago (Tyrrhenian sea, Italy). J. Foraminifer. Res. 2010, 40, 313–326. [Google Scholar]
- Jonasson, K.E.; Schrijder-Adams, C.J.; Patterson, R.T. Benthic foraminiferal distribution at Middle Valley, Juan de Fuca Ridge, a northeast Pacific hydrothermal venting site. Mar. Micropaleontol. 1995, 25, 151–167. [Google Scholar]
- Contreras-Rosales, L.A.; Koho, K.A.; Duijnstee, I.A.P.; De Stigter, H.C.; García, R.; Koning, E.; Epping, E. Living deep-sea benthic foraminifera from the Cap de Creus Canyon (western Mediterranean): Faunal-geochemical interactions. Deep Sea Res. Part I Oceanogr. Res. 2012, 64, 22–42. [Google Scholar]
- Fontanier, C.; Jorissen, F.J.; Lansard, B.; Mouret, A.; Buscail, R.; Schmidt, S.; Kerhervé, P.; Buron, F.; Zaragosi, S.; Hunault, G.; et al. Live foraminifera from the open slope between Grand Rhone and Petit Rhone Canyons (Gulf of Lions, NW Mediterranean). Deep Sea Res. Part I Oceanogr. Res. 2008, 55, 1532–1553. [Google Scholar]
- Alavi, S.N. Late Holocene deep-sea benthic foraminifera from the Sea of Marmara. Mar. Micropaleontol. 1988, 13, 213–237. [Google Scholar]
- Sen Gupta, B.K.; Machain-Castillo, M.L. Benthic foraminifera in oxygen-poor habitats. Mar. Micropaleontol. 1993, 20, 183–201. [Google Scholar]
- Bernhard, J.M.; Sen Gupta, B.K. Foraminifera of Oxygen-Depleted Environments. In Modern Foraminifera; Sen Gupta, B., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 201–216. [Google Scholar]
- Jorissen, F.J. The distribution of benthic foraminifera in the Adriatic Sea. Mar. Micropaleontol. 1987, 12, 21–48. [Google Scholar]
- Schmiedl, G.; Mackensen, A.; Müller, P.J. Recent benthic foraminifera from the eastern South Atlantic Ocean: Dependence on food supply and water masses. Mar. Micropaleontol. 1997, 32, 249–287. [Google Scholar]
- Martins, M.V.; Dubert, J.; Jouanneau, J.-M.; Weber, O.; Da Silva, E.F.; Patinha, C.; Alveirinho Dias, J.M.; Rocha, F. A multiproxy approach of the Holocene evolution of shelf-slope circulation on the NW Iberian Continental Shelf. Mar. Geol. 2007, 239, 1–18. [Google Scholar]
- Sorokin, P.Y. Oxidation of reduced compounds of sulfur in volcanically active areas of the Bay of Plenty (New Zealand) and Matupi Harbor (The New Britain Island, Papua New Guinea). Izv. AN USSR Ser. Biol. 1991, 3, 376–387. [Google Scholar]
- Tarasov, V.G.; Gebruk, A.V.; Mironov, A.N.; Moskalev, L.I. Deep-sea and shallowwater hydrothermal vent communities: Two different phenomena? Chem. Geol. 2005, 224, 5–39. [Google Scholar]
- Bárcenas, P.; Lobo, F.; Macías, J.; Fernández-Salas, L.; del Río, V.D. Spatial variability of surficial sediments on the northern shelf of the Alboran Sea: The effects of hydrodynamic forcing and supply of sediment by rivers. J. Iber. Geol. 2011, 37, 195–214. [Google Scholar]
- Tortora, P. La superficie deposizionale del delta sottomarino del Tevere: Zonazione del sedimento e processi associati. Boll. Soc. Geol. Ital. 1995, 114, 89–105. [Google Scholar]
- Díaz, J.; Palanques, A.; Nelson, C.H.; Guillén, J. Morpho-structure and sedimentology of the Holocene Ebro prodelta mud belt (northwestern Mediterranean Sea). Cont. Shelf Res. 1996, 16, 435–445. [Google Scholar]
- Mulder, T.; Syvitski, J.P.M. Turbidity currents generated at river mouths during exceptional discharges to the world oceans. J. Geol. 1995, 103, 285–299. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Latitude | Longitude | Location | Depth (m) |
|---|---|---|---|---|
| 36 | 38°08′52″ N | 15°07′53″ E | Inner shelf (g) | 41 |
| 37 | 38°08′48″ N | 15°07′39″ E | Inner shelf (u) | 35 |
| 38 | 38°09′12″ N | 15°07′47″ E | Middle shelf (g) | 60 |
| 39 | 38°09′11″ N | 15°07′27″ E | Middle shelf (g) | 64 |
| 40 | 38°09′32″ N | 15°08′21″ E | Middle shelf (u) | 80 |
| 41 | 38°09′36″ N | 15°07′50″ E | Middle shelf (g) | 82 |
| 42 | 38°09′45″ N | 15°07′22″ E | Middle shelf (g) | 96 |
| 43 | 38°10′13″ N | 15°07′24″ E | Outer shelf (g) | 120 |
| 44 | 38°10′06″ N | 15°07′53″ E | Outer shelf (u) | 112 |
| 45 | 38°10′12″ N | 15°08′23″ E | Outer shelf (u) | 113 |
| 46 | 38°10′36″ N | 15°07′51″ E | Outer shelf (g) | 144 |
| 47 | 38°10′31″ N | 15°07′21″ E | Outer shelf (g) | 160 |
| 48 | 38°09′15″ N | 15°05′35″ E | Middle shelf | 83 |
| ID | % Gravel | % Sand | % Silt | % Clay | Folk Sediment Class | Sorting | Skewness | Kurtosis |
|---|---|---|---|---|---|---|---|---|
| 36 | 0 | 19.97 | 67.07 | 12.96 | Sandy silt | 1.82 | 0.17 | 0.88 |
| 37 | 0 | 30.82 | 58.75 | 10.43 | Sandy silt | 1.86 | 0.28 | 0.89 |
| 38 | 0 | 25.55 | 60.93 | 13.52 | Sandy silt | 1.9 | 0.16 | 0.79 |
| 39 | 0.29 | 15.51 | 67.98 | 16.22 | (s) Gravelly sandy mud | 1.86 | 0.06 | 0.86 |
| 40 | 0.18 | 6.85 | 74.54 | 18.43 | (s) Gravelly mud | 1.69 | 0.1 | 0.92 |
| 41 | 0 | 21.01 | 63.07 | 15.92 | Sandy silt | 1.9 | 0.07 | 0.79 |
| 42 | 0 | 10.2 | 70.31 | 19.49 | Sandy silt | 1.81 | 0.05 | 0.89 |
| 43 | 0 | 4.44 | 73.41 | 22.15 | Silt | 1.67 | 0.1 | 0.9 |
| 44 | 0 | 6.89 | 73.36 | 19.75 | Silt | 1.7 | 0.09 | 0.91 |
| 45 | 0 | 11.84 | 69.52 | 18.64 | Sandy silt | 1.84 | 0.04 | 0.87 |
| 46 | 1.97 | 22.32 | 57.42 | 18.29 | (s) Gravelly sandy mud | 3.23 | −0.37 | 1.15 |
| 47 | 11.5 | 11.5 | 58.8 | 18.2 | Gravelly mud | 3.84 | −0.48 | 1.62 |
| 48 | 6.38 | 4.99 | 68.61 | 20.02 | Gravelly mud | 2.98 | −0.25 | 2.09 |
| ID | Location | Transect | Depth (m) | ALD5 | Averaged Values for 0–5 cm Interval | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ind/g | Aggl. | Hyal. | Porc. | α-Fisher | H | |||||
| 36 | Inner shelf (g) | C | 41 | 1.5 | 17.05 | 10.67 | 6.28 | 0.09 | 7.74 | 2.12 |
| 37 | Inner shelf (u) | W | 35 | 1.2 | 11.25 | 6.21 | 4.87 | 0.17 | 4.69 | 1.87 |
| 38 | Middle shelf (g) | C | 60 | 1.2 | 10.11 | 8.48 | 1.63 | 0.00 | 8.09 | 2.13 |
| 39 | Middle shelf (g) | W | 64 | 0.9 | 23.82 | 16.95 | 6.88 | 0.00 | 7.14 | 1.55 |
| 40 | Middle shelf (u) | E | 80 | 1.9 | 353.70 | 224.69 | 129.01 | 0.00 | 8.72 | 2.46 |
| 41 | Middle shelf (g) | C | 82 | 1.9 | 38.99 | 9.68 | 29.31 | 0.00 | 6.79 | 2.01 |
| 42 | Middle shelf (g) | W | 96 | 1.3 | 6.06 | 4.25 | 1.21 | 0.00 | 4.53 | 1.41 |
| 43 | Outer shelf (g) | W | 120 | 1.0 | 68.18 | 26.51 | 41.67 | 0.00 | 9.18 | 1.89 |
| 44 | Outer shelf (u) | C | 112 | 1.1 | 12.45 | 7.78 | 4.67 | 0.00 | 10.97 | 2.30 |
| 45 | Outer shelf (u) | E | 113 | 1.7 | 175.86 | 45.69 | 130.17 | 0.00 | 9.30 | 2.25 |
| 46 | Outer shelf (g) | C | 144 | 0.8 | 35.29 | 17.65 | 17.47 | 0.18 | 8.36 | 1.93 |
| 47 | Outer shelf (g) | W | 160 | 1.5 | 41.66 | 14.23 | 26.13 | 0.13 | 8.33 | 1.94 |
| 48 | Middle shelf | - | 83 | 1.1 | 79.13 | 45.04 | 33.33 | 0.76 | 16.63 | 2.48 |
| ID | Location | Depth (m) | Averaged Values for 0–5 cm Interval | ||||
|---|---|---|---|---|---|---|---|
| Ind/g | Aggl. | Hyal. | Porc. | Allocht. | |||
| 36 | Inner shelf (g) | 41 | 260.92 | 131.03 | 126.44 | 3.45 | 7.74 |
| 37 | Inner shelf (u) | 35 | 111.39 | 48.92 | 61.29 | 1.07 | 4.69 |
| 38 | Middle shelf (g) | 60 | 161.25 | 69.90 | 90.66 | 0.69 | 8.09 |
| 39 | Middle shelf (g) | 64 | 223.26 | 76.74 | 146.52 | 0.00 | 7.14 |
| 40 | Middle shelf (u) | 80 | 162.70 | 23.02 | 139.68 | 0.00 | 8.72 |
| 41 | Middle shelf (g) | 82 | 760.56 | 105.63 | 654.93 | 0.00 | 6.79 |
| 42 | Middle shelf (g) | 96 | 41.75 | 19.21 | 22.55 | 0.00 | 4.53 |
| 43 | Outer shelf (g) | 120 | 1689.87 | 297.47 | 1348.10 | 44.30 | 9.18 |
| 44 | Outer shelf (u) | 112 | 668.42 | 136.84 | 526.32 | 5.26 | 10.97 |
| 45 | Outer shelf (u) | 113 | 1047.37 | 163.15 | 873.68 | 10.53 | 9.30 |
| 46 | Outer shelf (g) | 144 | 569.18 | 78.62 | 481.13 | 9.43 | 8.36 |
| 47 | Outer shelf (g) | 160 | 234.88 | 15.12 | 219.77 | 0.00 | 8.33 |
| 48 | Middle shelf | 83 | 1572.65 | 478.63 | 1085.47 | 8.55 | 16.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Bella, L.; Pierdomenico, M.; Bove, C.; Casalbore, D.; Ridente, D. Benthic Foraminiferal Response to Sedimentary Processes in a Prodeltaic Environment: The Gulf of Patti Case Study (Southeastern Tyrrhenian Sea). Geosciences 2021, 11, 220. https://doi.org/10.3390/geosciences11050220
Di Bella L, Pierdomenico M, Bove C, Casalbore D, Ridente D. Benthic Foraminiferal Response to Sedimentary Processes in a Prodeltaic Environment: The Gulf of Patti Case Study (Southeastern Tyrrhenian Sea). Geosciences. 2021; 11(5):220. https://doi.org/10.3390/geosciences11050220
Chicago/Turabian StyleDi Bella, Letizia, Martina Pierdomenico, Cristiano Bove, Daniele Casalbore, and Domenico Ridente. 2021. "Benthic Foraminiferal Response to Sedimentary Processes in a Prodeltaic Environment: The Gulf of Patti Case Study (Southeastern Tyrrhenian Sea)" Geosciences 11, no. 5: 220. https://doi.org/10.3390/geosciences11050220
APA StyleDi Bella, L., Pierdomenico, M., Bove, C., Casalbore, D., & Ridente, D. (2021). Benthic Foraminiferal Response to Sedimentary Processes in a Prodeltaic Environment: The Gulf of Patti Case Study (Southeastern Tyrrhenian Sea). Geosciences, 11(5), 220. https://doi.org/10.3390/geosciences11050220