1. Introduction
Sweet potato vine (SPV) is the by-product of sweet potato, consisting of leaf, stem, and stalk. In a previous study, we suggested that adding fresh SPV to gilt diet improved follicular development and thus benefited reproductive performance in Chinese Meishan (MS) gilt [
1]. A large number of microorganisms are present in the intestine of pigs, which form a complex and relatively stable ecosystem with the body [
2,
3,
4]. These microorganisms directly or indirectly participate in the nutrient metabolism and immune regulation of the host organism, but also suffer from various factors involving the immune system [
2], intestinal environment [
3], age [
4], and diet [
5]. Changes in the composition and metabolic activity of intestinal microbes caused by food intake have received extensive attention [
5,
6,
7]. The SPV [dry matter, 10.13%; digestible energy (DE), kcal/kg, 2837.00; crude protein (CP), 14.90%, at a dry matter basis] contains crude fiber (CF, 15.51%), neutral detergent fiber (NDF, 21.63%), acid detergent fiber (ADF, 19.37%), and acid detergent lignin (ADL, 3.3%) [
8,
9]. However, the SPV as a source of fiber for remodeling microbiota in MS gilt has not yet been reported.
Dietary fiber have a significant effect on the microflora structure of pig intestines [
10,
11]. Concurrently, different fiber types and sources may directly affect the composition of intestinal colonies in monogastric animals [
10,
11]. Although different sources of fiber (maize fiber, soybean fiber, wheat bran fiber, and pea fiber) to feed fattening pigs were used, no difference in total bacteria populations was observed between diets. However, the composition of the microbial community was affected, in which lower ileal
Lactobacillus populations and higher cecal
Escherichia coli populations were found in a soybean fiber treatment in comparison with the control diet. Meanwhile, cecal
Lactobacillus populations in pea fiber and
Bifidobacterium populations in wheat bran were higher than control [
10]. Feeding mice with soluble fiber (inulin) and insoluble fiber (cellulose) found that the addition of soluble fiber significantly increased the content of SCFA in the mouse cecum, indicating that soluble fiber is more easily fermented by intestinal microbes [
11]. The addition of soluble fiber (inulin) increases the abundance of butyrate-producing bacteria (such as
E. rectale,
Roseburia intestinalis, and
Anaerostipes) [
12] while reducing the abundance of
Akkermansia muciniphila, which affects the metabolism and absorption of host nutrients [
13]. Zhou et al. (2017) [
7] found that the addition of 1.5% soluble fiber inulin to the diet of a sow improved the maternal body weight, metabolism, inflammatory status, and distinct changes in the microbial community. The addition of SPV causes changes in the microbial fermentation substrate of the hindgut to affect the microbes in the intestines of the gilt. Whether SPV affects the gut health of the gilt by affecting the intestinal microbes and metabolites needs further study.
Therefore, this study investigated the effects of the sweet potato vine on the intestinal and plasma metabolites as well as the colon microbial composition in MS gilt.
2. Materials and Methods
Animal procedures in this study were approved by the Animal Experimental Committee of Sichuan Agricultural University, under the Ethic Approval Code: SCAUAC201408-10.
2.1. Animals and Experimental Design
This study used 20 prepubertal MS gilts with weights at 30 ± 0.18 kg (109.00 ± 2.10 day). Gilts were randomly allocated to control (CON, n = 10) and sweet potato vine (SPV, n = 10) treatments. Gilts were housed in individual stalls, and each gilt was one replicate. The animals as well as their basal diets, feeding regime, and treatments were the same as those found in our previous study [
1]. Briefly, basal diet included 3.08 Mcal DE /kg, 14.00% of CP, 0.68% Lysine, 0.53% calcium, and 0.48% phosphorus. Fresh SPV was provided as part (DE intake, 554 kcal/d; CP intake, 26 g/d,) of the basal diet to make sure gilt in SPV treatment had the same amount of energy and protein intake per day as in control group. CON treatment fed 1.64 kg basal diets per day. SPV treatment fed 1.46 kg basal diets and 2.00 kg fresh SPV per day. The two treatments offered a similar nutrient intake with the exception of CF (51.00 g/d vs. 73.94 g/d). Before feeding, fresh SPV was chopped into small pieces and then mixed with the powdered basal diet to feed the gilts. Gilts were fed twice daily (08:00 and 14:00) and water was offered ad libitum from a nipple drinker. The experiment ended at the morning of day 19 after the third estrus cycle of gilt.
2.2. Nutrient Digestibility Determination
When the body weight of gilt reached 40 kg, 12 healthy gilts (six gilts per treatment) were used in a nutrient digestibility trial to evaluate the effects of SPV on dry matter (DM), energy, nitrogen, CF, and fiber digestibility. Gilts were individually housed in metabolism cages and fed their respective experimental diets. The feeding was the same as the main study. The experimental diets were fed for a three-day adjustment period followed by a four-day formal feeding period. During the first and last day of formal feeding, 5 g ferric oxide was added to the basal diet as a color marker, respectively. Fecal samples were collected on a full day, on the first day at which red fecal samples appeared until the last day red fecal samples appeared. Samples were pooled, placed in plastic bags, and frozen at −20 °C. At the end of collection, feces were dried in a force-draft oven (65 °C) for 2 d and then weighed. Dried feces were ground through a 1-mm screen and frozen until subsequent fiber, nitrogen, crude fat, and gross energy determination could be performed. DM, ether extract (EE), and CF digestibility was determined using the AOAC procedure (2000) [
14]. Nitrogen content of diets and feces was determined by the combustion method using a Leco TruSpec analyzer. Gross energy of feed and feces was determined by a Parr 6400 bomb calorimetry (Parr Instrument Co., Moline, IL, USA).
2.3. Data Record and Sample Collection
On the morning of day 19 after the third estrus cycle of gilt (SPV treatment reached estrous 9.4 days later than control), live weight of all gilts (n = 10 / each treatment) was recorded, and then 10 mL of blood sample was collected by acute jugular puncture prior to slaughter to obtain the plasma. The plasma was stored at −20 °C until analysis.
The gilts were slaughtered by an intracardial injection of sodium pentobarbital (50 mg/kg body weight) and bled by exsanguination. After the gilt was slaughtered, the head, tail, legs, and all internal organs (visceral organs and endocrine glands), as well as perirenal and omental adipose depots, were removed. The hot carcass weight was then recorded to determine dressing percentage. Loin muscle area was measured by planimetry on a cut at the level of the 13th and 14th ribs. Backfat thickness in carcasses represents the average of measures at the levels of the third and eleventh thoracic vertebrae and of the smallest measure over the gluteus medius muscle in the lumbar region according to Rehfeldt et al. (2012) [
15]. For dissection-derived body composition, the left carcass was immediately further separated into muscle tissue, subcutaneous adipose tissue (SCAT), skin, and bones.
After the stomach and entire intestine was rapidly removed, the weight of stomach, small intestine (include the duodenum, jejunum, and ileum), large intestine (include the cecum, colon, and rectum) was recorded. Three segments of 2 cm were cut at the mid-jejunum and mid-ileum separately, as described by Wang et al. (2014) [
16]. The 2-cm intestinal segments were flushed gently with ice-cold PBS (pH 7.4) and then placed in 10% fresh, chilled formalin solution for histological measurement. A 10-cm section of caecum and colon was ligated and removed to avoid digesta admixture in different intestinal sections. The intestine section was placed on a chilled stainless steel tray. A small incision was opened in the distal end of each segment, and the digesta was collected in a 5-mL tube, then rapidly frozen in liquid nitrogen and stored at −80 °C. Samples of liver were collected, then quickly frozen in liquid nitrogen and stored at −80 °C until analysis.
2.4. Plasma Analyses
The concentrations of glucose (Glu), non-esterified fatty acid (NEFA), triglycerides (TG), total cholesterol (TC), and urea in plasma were detected by commercial kits according to the manufacturer’s instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, Jiangsu, China). Optical density (OD) values were determined at 520 nm, 440 nm, 546 nm, and 510 nm by a MuLtisKan MK3-Thermo Labsystems microplate reader (Thermo Labsystems, CA, USA). Plasma zonulin and endotoxin were measured using sow enzyme-linked immunosorbent assay (ELISA) Kits (R&D Systems Inc., Minneapolis, MN, USA). Optical density (OD) values were determined at 450 nm by a MuLtisKan MK3-Thermo Labsystems microplate reader (Thermo Labsystems, CA, USA). Minimal detection limits for Glu, NEFA, TG, TC, urea, endotoxin, and zonulin were 0.5 mmol/L, 10 μmol/L, 0.02 mmol/L, 0.1 mmol/L, 0.1 mmol/L, 3.1 ng/L, and 3 ng/L.
2.5. Analysis of Digesta
The endotoxin, lipocalin-2, interleukin-6 (IL-6), interleukin-10 (IL-10), tumor necrosis factor-α (TNF-α) of colonic digesta were measured using sow ELISA Kits (R&D Systems Inc., Minneapolis, MN, USA). The index analysis was performed following manufacturer’s instructions. Optical density (OD) values were determined at 450 nm through the use of a MuLtisKan MK3-Thermo Labsystems microplate reader (Thermo Labsystems, CA, USA). Minimal detection limits for endotoxin, lipocalin-2, IL-6, IL-10, TNF-α and zonulin were 3.1 ng/L, 10 ng/L, 12.5 ng/L, 5 ng/L, 7 ng/L, and 3 ng/L.
Short-chain fatty acids including acetate, propionate, and butyrate in digesta of caecum and colon were analyzed in a gas chromatographic system (GC, VARIAN CP-3800, Varian, Palo Alto, CA, USA). Briefly, a 2-g sample was suspended in distilled water (5 mL) and allowed to stand for 30 min, after which it was centrifuged at 4°C, 12,000 g for 10 min. The sample was then transferred to 2 mL of supernatant and mixed with 0.4 mL of metaphosphoric acid. Placed at 4 °C for 30 min, the sample was centrifuged again at 4 °C, 12000 g for 10 min before being transferred to 1.2 mL of supernatant and mixed with 15.2 μL of crotonic acid (210 mmol/L, internal standard), after which 0.3-mL liquid was mixed with 0.3-mL methanol. Finally, the mixed solution was filtered by the 0.22-μm filter membrane for the following gas chromatography analysis, which equipped with a micro-injector (10 μL), a flame ionization detector, and a capillary chromatographic column (CP-FFAP, 25 m × 0.32 mm × 0.3 μm).
2.6. Gene Expression
As described in our previous study [
1], total RNA was extracted from frozen liver samples by TRIzol reagent (TaKaRa Biotechnology Co. Ltd., Dalian, China) according to the manufacturer’s instructions. Reverse transcription (RT) kit (Invitrogen, Carlsbad, CA, USA) was used to synthesize cDNA. Real-time PCR was used to quantify fatty acid synthetase (
FAS), sterol regulatory element binding proteins (
SREBP-1), carnitine palmityl transferase (
CPT-1), and peroxisome proliferator activated receptor-α (
PPAR-α) mRNA expression levels using the Sybr Green Kit (Qiagen, Valencia, CA, USA). Amplification was carried out with 12.5 µL final reaction volume containing 5 µL of Sybr green master mix, 0.5 µL of each primer (final concentration 0.25 µM), 6 µL of diethyl pyrocarbonate (DEPC)-water, and 0.5 µL of sample cDNA. All real-time PCRs were carried out in triplicate by a DNA Engine thermal cycler (PTC-0200, Chromo4 Real-Time detector, Bio-Rad, Hercules, CA, USA). The thermal cycling conditions were denaturation for 10 min at 95 °C, amplification for 45 cycles with denaturation at 95 °C for 20 s, annealing at 60 °C for 10 sec, extension at 72 °C for 9 sec, followed by fluorescence acquisition at 60 °C for 5 sec. Using the 2
−∆∆Ct method to calculate the relative level of target gene expression [
17], β-actin was amplified as housekeeping gene. Sequences of the primers are listed below.
FAS, forward CTACATCGAGTGCATCAGACAGG and reverse GAACAGGAAGAGGCTGTGGTT;
SREBP-1, TTGAGGACAGCCAGATGAAGC and reverse GCAGGAGAGACAGAGGAAGAC;
CPT-1, CTACACAGAGACGGGGCACT and reverse AGGGCAAGAACTGGAAGCAG;
PPAR-α, CTATCATTTGGTGCGGAGACC and reverse GGAGTTTGGGGAAGAGAAAGAC;
β-Actin, forward GGCCGCACCACTGGCATTGTCAT and reverse AGGTCCAGACGCAGGATGGCG.
2.7. Bacterial Community Analysis
The Mo Bio PowerFecalTM DNA Isolation Kit (MO BIO Laboratories, Carlsbad, CA, USA) was utilized to extract the microbial DNA of thawed fecal sample (0.5 g). A nucleic acid/protein analyzer (Beckman DU-800, Beckman Coulter, Inc., CA, USA) was used to detect the concentration and purity of the genomic DNA. Following this, DNA samples were sent to perform amplicon pyrosequencing on Illumina HiSeq PE250 platforms at Novogene Bioinformatics Technology in Beijing, China. The V4 hypervariable region of the 16S rRNA gene was amplified using a forward primer 515f (5′-GTGCCAGCMGCCGCGGTAA-3′) and a reverse primer 806r (5′-GGACTACHVGGGTWTCTAAT-3′).
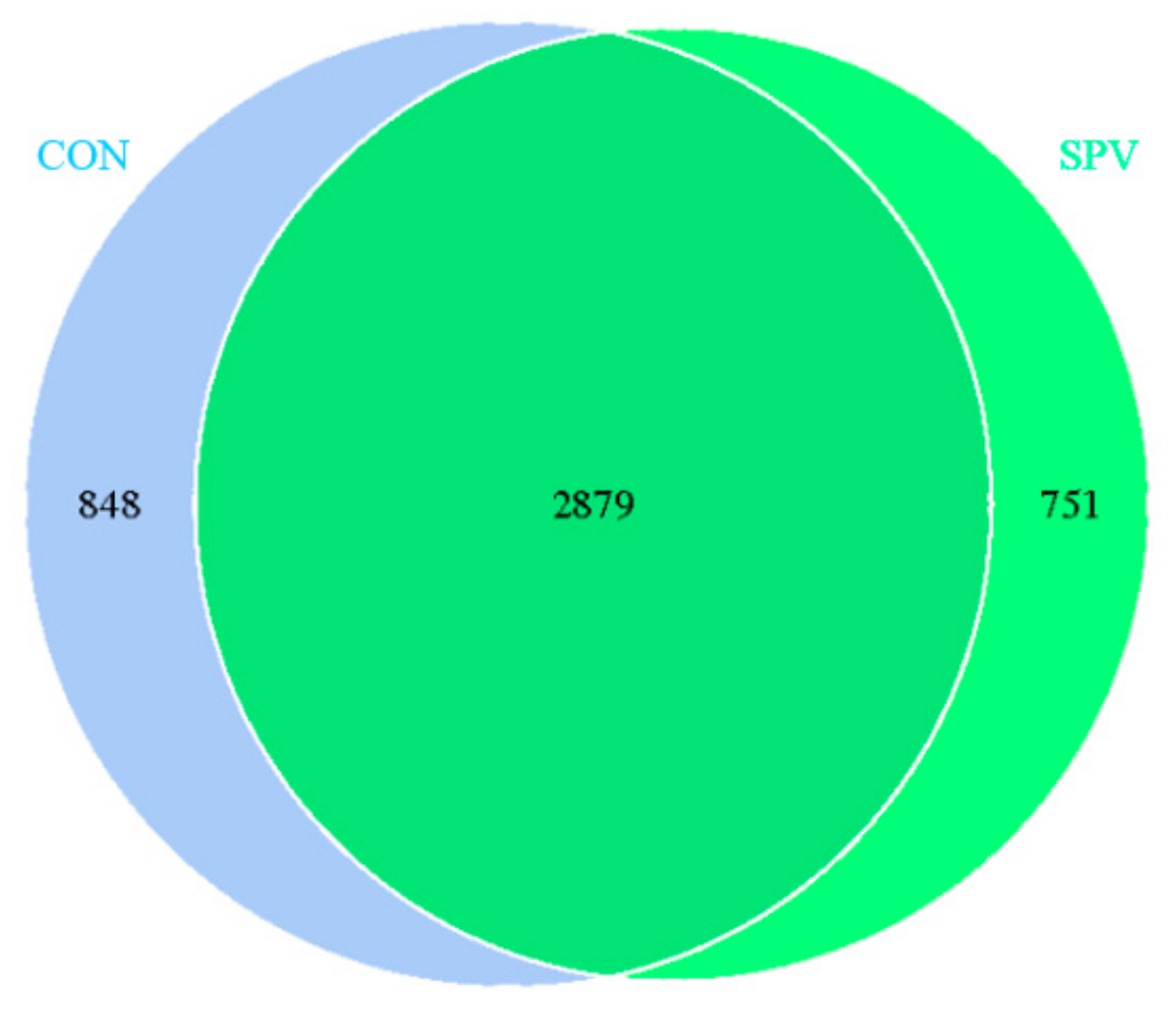
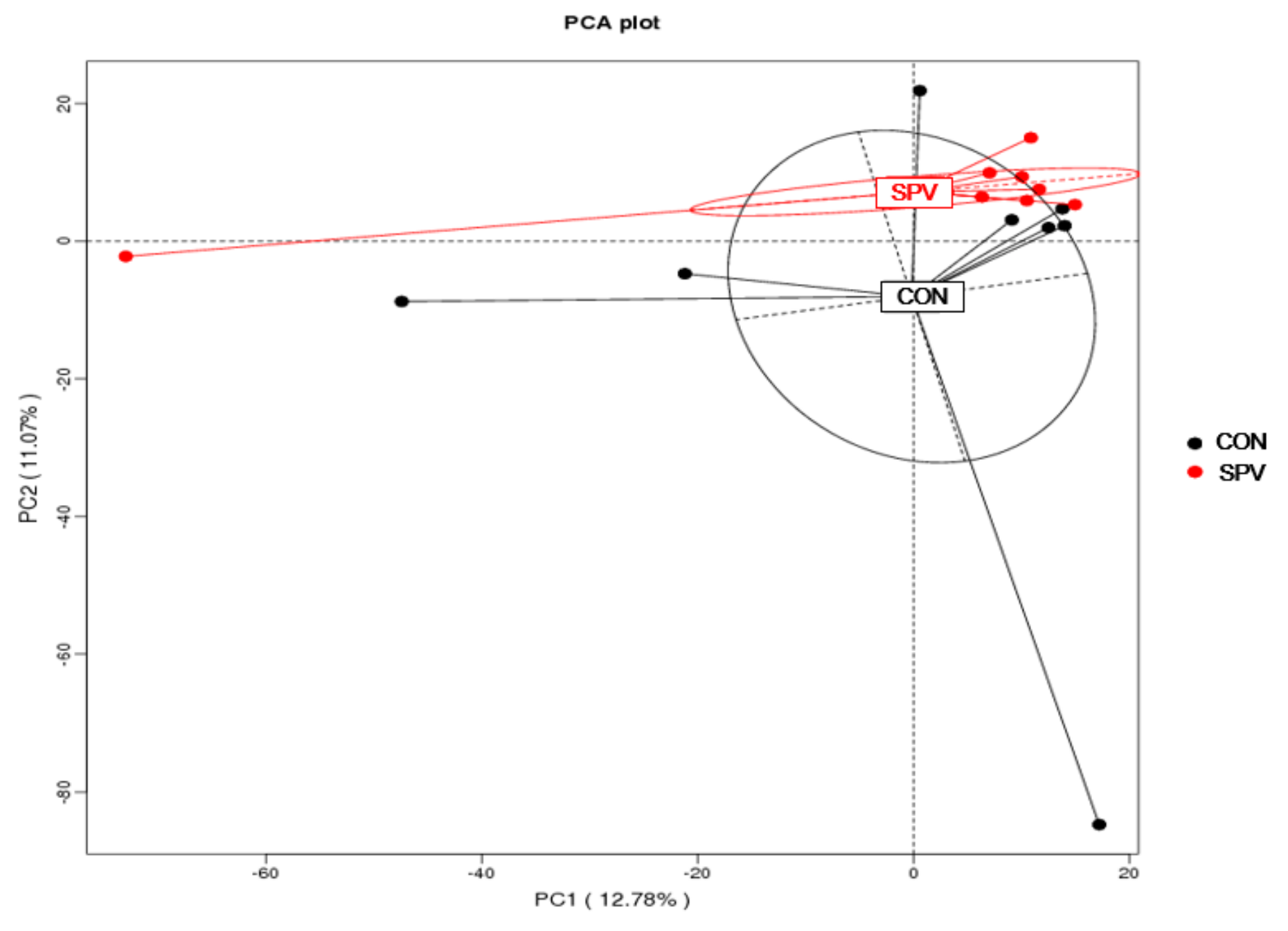
Operational taxonomic units (OTUs) come from high-quality tags utilizing Uparse v7.0.1001 (
http://drive5.com/uparse/) at an identity threshold of 97% used Ribosomal Database Project (RDP) classifier Version 2.2 (
http://github.com/rdpstaff/) to assign taxonomy for 16S rRNA gene sequences. The relative abundance of each OTU was explored at phylum and genus levels. A Venn diagram was generated for a comparison of the OTUs of the two treatments. Alpha diversity values for each sample were assessed by Qiime 1.7.0. (
http://qiime.org/home_static/dataFiles.html) Principal coordinates analysis (PCoA) plots were produced using unweighted UniFrac metrics.
2.8. Statistical Analysis
Data of relative abundance at phylum and genus level were log-transformed before statistical analysis. Following the normal distribution tested, data (Tables) were analyzed by an independent-samples t-test using SPSS 21.0 (IBM SPSS Company, Chicago, IL, USA). Other data, as seen in
Figure 1 and
Figure 2, were analyzed by t-test using GraphPad Prism analysis software. Differences were considered significant at
p ≤ 0.05, whereas
p < 0.10 was considered as a tendency.
4. Discussion
This was the first study to focus on the effect of SPV on intestinal digestion, the intestinal and plasma metabolites, and colon microbial composition in Chinese Meishan gilts. In this study, the addition of fresh SPV did not significantly affect the apparent digestibility of nutrients. On the basis of our original knowledge, the fiber in the diet cannot be digested in the small intestine; it will adhere and dilute other nutrients, thereby interfering with the absorption of other nutrients. Accordingly, the addition of fiber may not be conducive to production. Previous studies have confirmed that increased fiber levels in the diet are not beneficial to the digestion of nutrients, such as crude protein, energy, and fat [
18,
19]. After the addition of four different levels of fiber into the diets of four different hybrid pig breeds, the results showed that dietary fiber levels significantly affected the apparent digestibility of nutrients. With the reduction of local pig bloodline, the digestibility of acidic and neutral detergent fibers gradually decreased [
20]. However, the use of fresh alfalfa instead of a part of basal diet to feed the finishing pigs showed that the apparent digestibility of dry matter, crude protein, and crude ash increased compared with the control group, but the digestibility of crude fat tended to decline [
21]. These results suggest that the apparent digestibility of nutrients in pig diets varies according to the fiber amount and type, and the local Chinese pig breed exhibits much more fiber tolerance. The MS gilt may be tolerant of higher fiber levels. In this study, we used fresh SPV, which had lower dry matter, and may not have reached an extent to which apparent digestibility was substantially affected. In addition, this experiment utilized the principle of equal energy and nitrogen substitution; the two treatment gilts had almost the same dry matter intake.
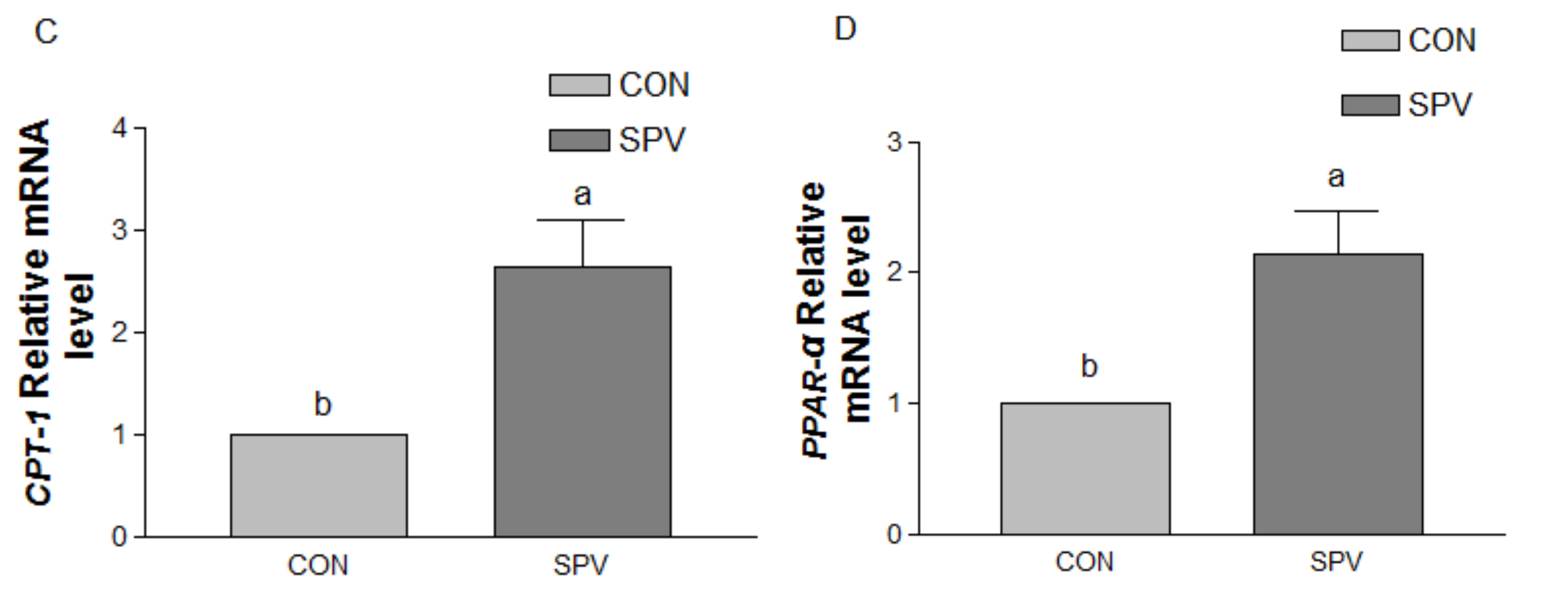
In the current study, the relative weight of the stomach, small intestine, and large intestine of MS gilt, as well as the thickness of the muscle layer of jejunum and ileum was increased significantly in SPV treatment. The addition of fiber into the diet affects the development of the gut of pigs [
22,
23]. Using high-, medium- and low-fiber diets to feed the piglets, the results demonstrated that a high fiber addition promoted the development of stomach and duodenum and increased the index of jejunum, ileum, cecum, and colon [
22]. An increase in the amount of NDF in the diet of piglets also resulted in an increase in weight and length of the large intestine [
23]. This might be attributable to the fiber in fresh SPV enters the digestive tract after which it swells and compresses gastrointestinal tract of the gilts, causing the weight and length of gastrointestinal tract to increase, resulting in a decrease in the slaughter rate (67.51% versus 63.68%) in this study. Moreover, the results of the body composition by manual dissection of the carcass in the present study showed that gilts in the SPV treatment had thinner backfat thickness and less percentage of subcutaneous adipose tissue; such evidence indicates that SPV could slow fat deposits. Consistent with these results, we also found that SPV treatment increased non-esterified fatty acid in the plasma as well as fatty acid catabolism gene expression level of CPT-1 and PPAR-α in the liver. Such results support our previous research that gilt in the SPV treatment reached puberty 9.4 days later than those in the CON treatment [
1]. Previous studies have proposed that puberty occurs only after the attainment of a minimum level of leanness or fatness, or ratio of fat to lean [
24]. Thus, gilt in the SPV treatment spent much more time to reach the minimum level of fatness to trigger the onset of puberty in comparison with those in the CON treatment.
In this study, the concentrations of butyric acid and acetate in caecal content and colonic content of gilt in SPV treatment were both increased; however, the concentrations of propionic acid did not. Supplemental fiber with different fiber components resulted in inconsistent SCFA production in the intestinal tract of pigs [
25]. Our results might be closely related to the fiber monosaccharide composition [
26]. Fibers with a high uronic acid content increased acetic acid production, fibers with high glucose content increased propionic acid production, and xylose promoted the production of butyric acid [
27]. Therefore, the content of uronic acid and xylose in the fresh SPV was relatively high; accordingly, the acetic acid and butyric acid produced by the fermentation were significantly enhanced. Studies in both rats and humans have demonstrated that SCFA can decrease the concentration of cholesterol in the blood [
28,
29], thus affecting lipid metabolism. However, in this study, SPV did not significantly reduce triglycerides and total cholesterol in the MS gilt blood. In addition, several studies have demonstrated that SCFA activates the oxidation of fatty acids by activating activity of AMPK in liver and muscle cells, inhibiting the denovo biogenesis of fatty acids and lipid catabolism [
30]. Accordingly, this may explain the slower fat deposition of gilt in the SPV treatment.
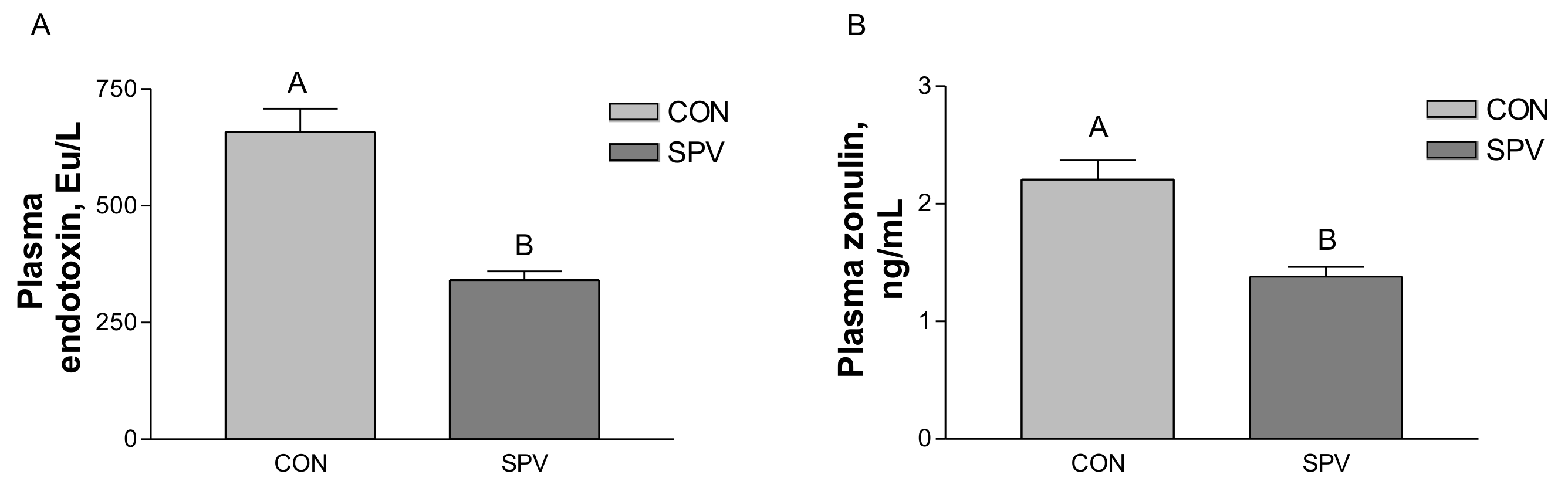
Zonulin concentration may enhance in blood when intestinal mucosal integrity is impaired [
31]. A previous study has found that, via a modulation of expression of tight junction proteins, butyrate maintains an intestinal barrier integrity [
32]. In this study, we observed that gilt fed SPV diets experienced decreased levels of zonulin in plasma and an increase in butyric acid concentration. Furthermore, SPV treatment reduced endotoxin in blood and colonic feces. In the colonic feces analysis, the anti-inflammatory cytokines IL-10 increased, and pro-inflammatory cytokines TNF-α and lipocalin-2 decreased in the SPV treatment. Cytokines play an important role in the inflammatory response [
33]. From these results, it may be concluded that SPV treatment decreased the gut permeability, ameliorated the inflammation state of intestinal mucosal epithelium as indicated by decreasing the zonulin concentration, increased the butyrate, reduced endotoxin, and changed inflammatory cytokines.
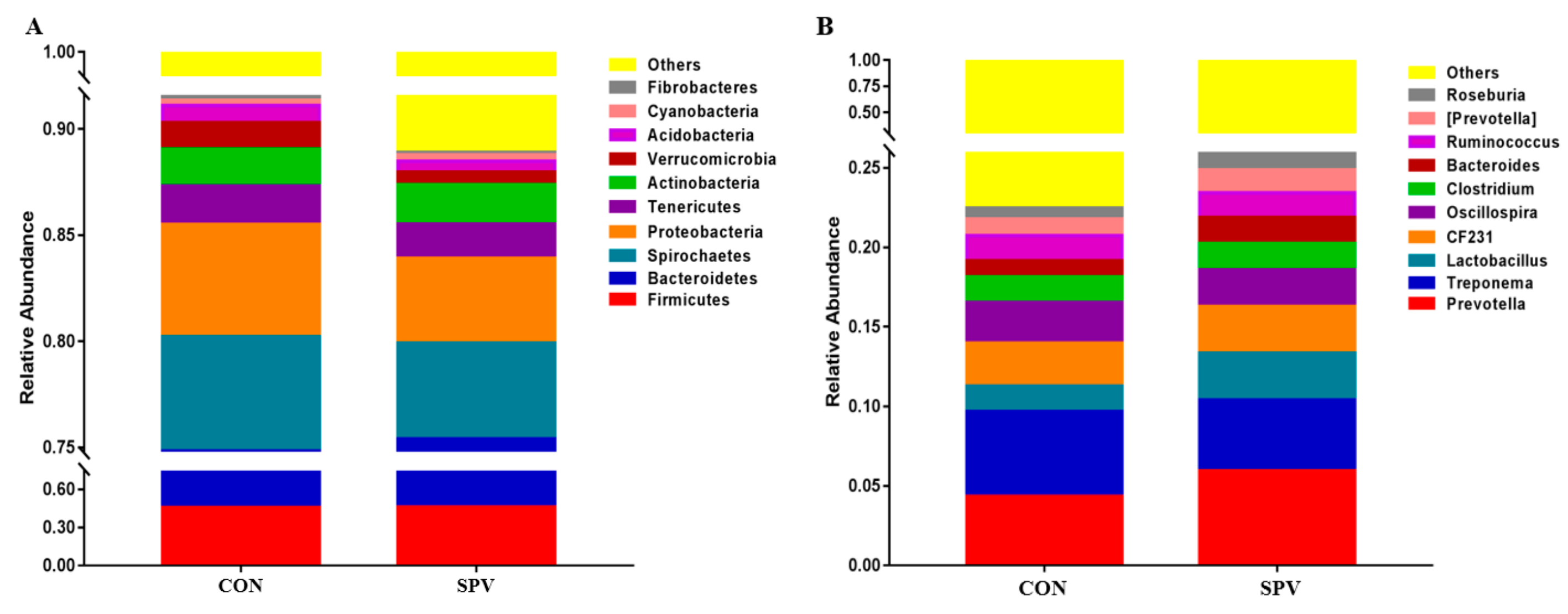
Abundant phyla
Firmicutes and
Bacteroidetes could be found in more than 75% of the colonic feces in this study, which play a role in degrading polysaccharides and promoting energy absorption in the gilt intestinal tract. Fresh SPV contained cellulose, hemicellulose, and pectin, which provided the substrate for the intestinal fiber-degrading bacteria. The microbes in the intestines of animals are “organs” that are composed of a series of microorganisms such as bacteria, archaea, and fungi. Microbes form a symbiotic relationship with the animal body and maintain the homeostasis of the intestinal environment. The hindgut digestion of a pig is mainly microbial digestion, in which the colon is the most developed in the hindgut. Most of the hard-to-digest fiber materials are decomposed by microbial fermentation in the hindgut to produce products such as acetic acid and butyric acid. The proportion of
Firmicutes and
Bacteroidetes is related to the body’s energy metabolism [
34]. In this study, the SPV feed increased the abundance of butyrate-producing bacteria like
Roseburia and
Lachnospira in the colonic feces in Meishan gilts.
Roseburia was part of commensal bacteria producing short-chain fatty acids, especially butyrate, affecting the colonic motility, immunity maintenance, and anti-inflammatory properties [
35]. It also reported that the reduction in
Roseburia contributed to the dysbiosis of ulcerative colitis patients [
36]. Weber [
37] and other studies have found that the
Lachnospira is the main pectin degrading bacterium in the human intestine. At the same time,
Bacteroides,
Lactobacillus, and
Bifidobacteria can grow in the presence of pectin and pectin oligosaccharides [
38]; pectin contained in the SPV may provide conditions for the growth of these bacteria. Accordingly, the bacterial abundance such as
Lactobacillus and
Bacteroides was enhanced in this study. Therefore, the SPV feed provided different fermentation substrates for the fermentation of intestinal microorganisms, thereby affecting the abundance of the bacteria and the SCFA production.
On the other hand, beneficial microorganisms colonized the intestinal mucosa to constitute a protective barrier to prevent colonization by other pathogenic bacteria [
39]. Pathogenic bacteria in the intestinal tract are harmful to the mucosal barrier and may induce intestinal diseases. In this study, SPV treatment increased the abundance of
Lactobacillus and
Bacteroides.
Lactobacillus is the only genus of the
Lactobacillus family, including the species of
Lactobacillus mucosae and
Lactobacillus reuteri.
Lactobacillus presents in the gut of animals and plays a major role in maintaining bacterial balance and preventing colonization of pathogenic microorganisms.
Lactobacillus enters the intestine after being adhered to the intestinal mucosa, during which a biological barrier is formed that can prevent the invasion of pathogenic microorganisms [
40,
41]. Simultaneously,
Lactobacillus reuteri can also secrete reuterin, which has a strong inhibitory effect on harmful bacteria [
42]. Therefore, SPV treatment may improve the intestinal health by promoting the abundance of certain beneficial bacteria, thereby inhibiting the growth of pathogenic bacteria.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}