A Review of Endothelium-Dependent and -Independent Vasodilation Induced by Phytochemicals in Isolated Rat Aorta



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolated Rat Aortic Preparation
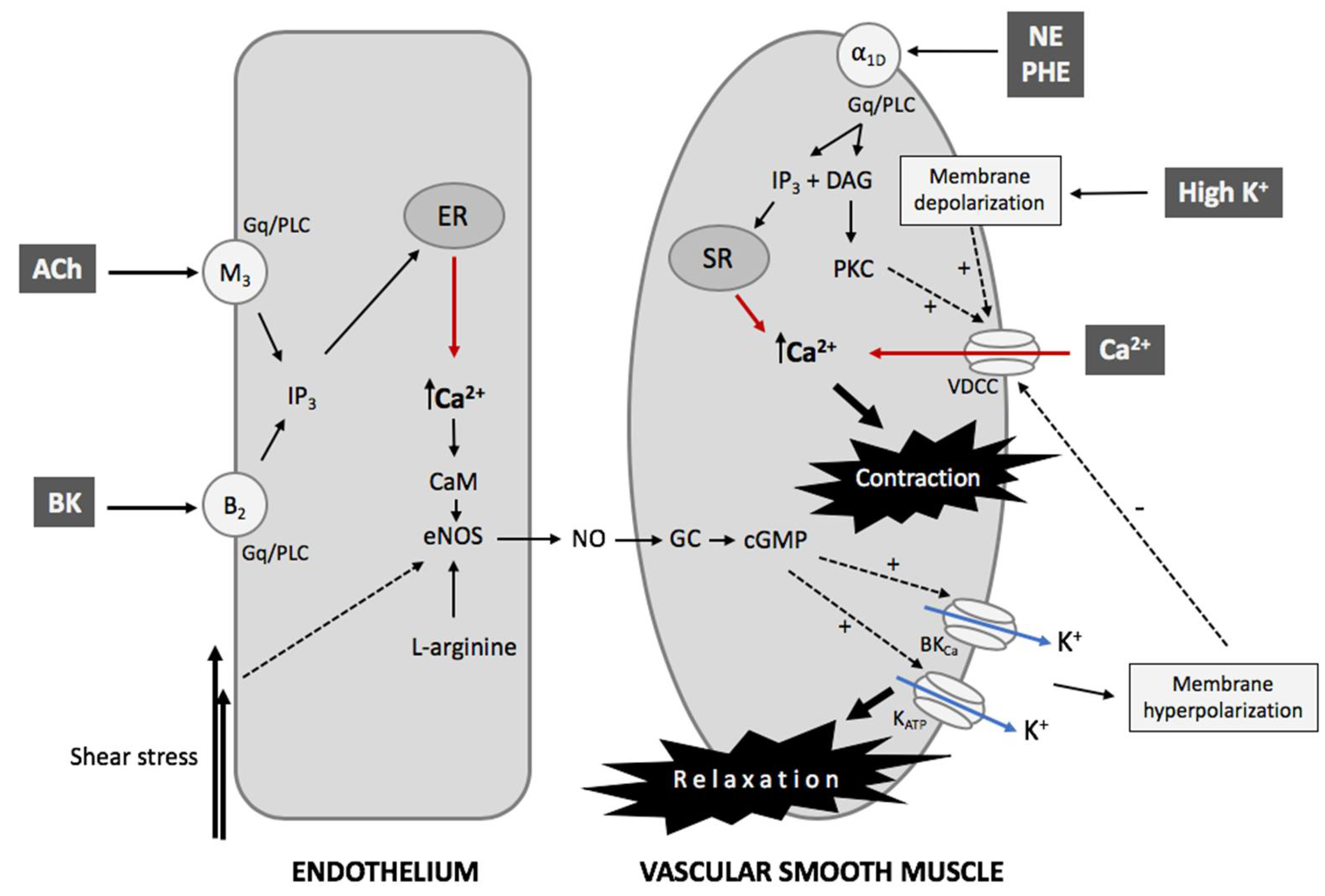
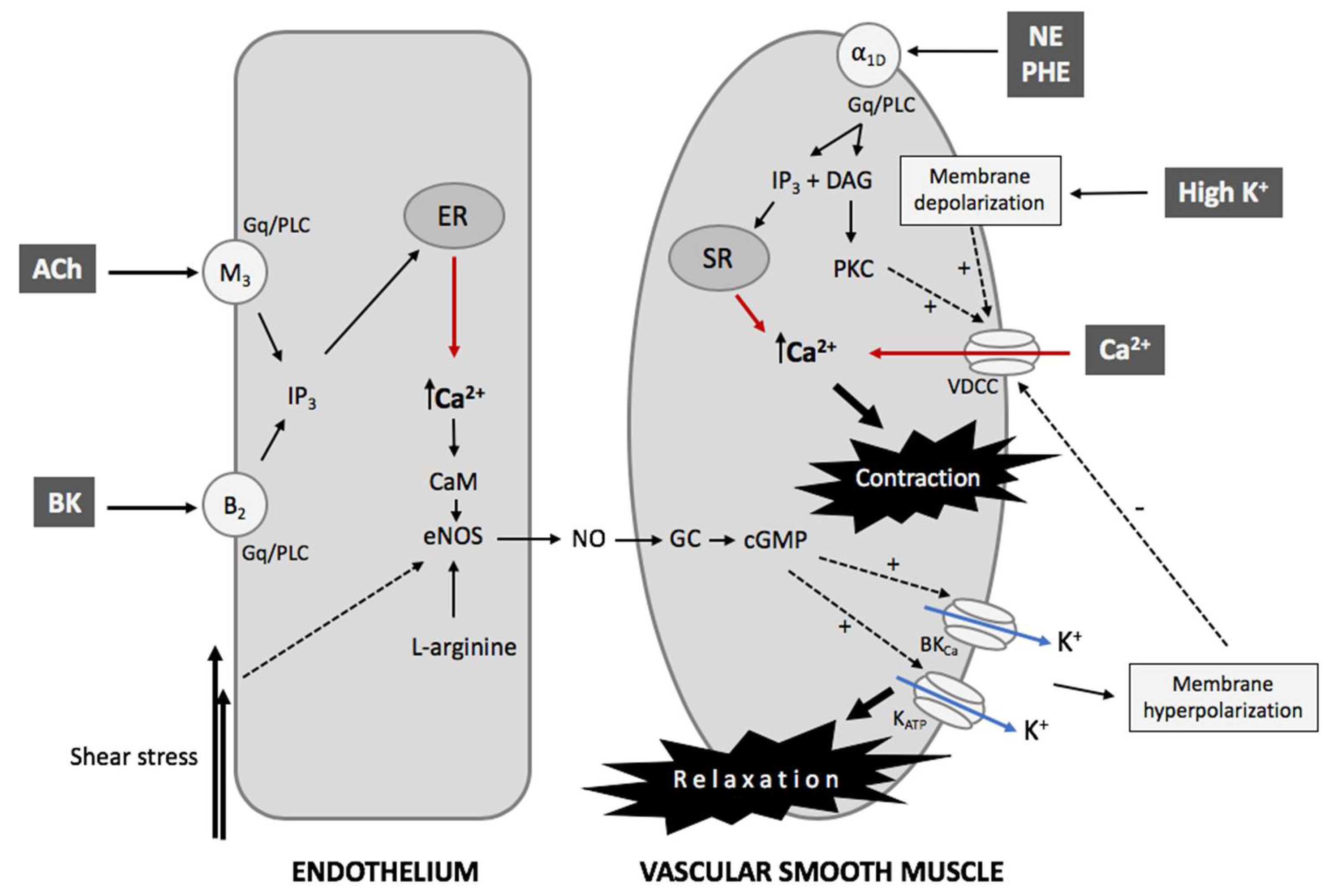
2.2. The Contractility Mechanism in Vessels
2.3. The Endothelial Control of VSMCs
3. Phytochemical Assays of Plant Extracts and Isolated Compounds
3.1. Plant Extracts
3.2. Alkaloids
3.3. Non-Alkaloid Compounds
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Furchgott, R.F.; Zawadzki, J.V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980, 288, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.M.; Ferrige, A.G.; Moncada, S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 1987, 327, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Ignarro, L.J.; Buga, G.M.; Wood, K.S.; Byrns, R.E.; Chaudhuri, G. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proc. Natl. Acad. Sci. 1987, 84, 9265–9269. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, S.; Kapela, A.; Tsoukias, N.M. Intercellular communication in the vascular wall: A modeling perspective. Microcirculation 2012, 19, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Reho, J.J.; Zheng, X.; Fisher, S.A. Smooth muscle contractile diversity in the control of regional circulations. Am. J. Physiol. Heart. Circ. Physiol. 2014, 306, H163–H172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, S.S.; Duling, B.R. Flow control among microvessels coordinated by intercellular conduction. Science 1986, 234, 868–870. [Google Scholar] [CrossRef] [PubMed]
- WHO. The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 1 April 2019).
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.; Coca, A.; De Simone, G.; Dominiczak, A.; et al. 2018 Practice Guidelines for the management of arterial hypertension of the European Society of Hypertension and the European Society of Cardiology: ESH/ESC task force for the management of arterial hypertension. J. Hypertens 2018, 36, 2284–2309. [Google Scholar] [CrossRef]
- Saklayen, M.G.; Deshpande, N.V. Timeline of history of hypertension treatment. Front. Cardiovasc. Med. 2016, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Cuspidi, C.; Tadic, M.; Grassi, G.; Mancia, G. Treatment of hypertension: The ESH/ESC guidelines recommendations. Pharmacol. Res. 2018, 128, 315–321. [Google Scholar] [CrossRef]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017. ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the prevention, detection, evaluation, and management of high blood pressure in adults: A report of the American College of Cardiology/American Heart Association task force on clinical practice guidelines. Circulation 2018, 138, e484–e594. [Google Scholar] [CrossRef]
- Serino, A.; Salazar, G. Protective Role of Polyphenols against Vascular Inflammation, Aging and Cardiovascular Disease. Nutrients 2019, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Poti, F.; Santi, D.; Spaggiari, G.; Zimetti, F.; Zanotti, I. Polyphenol health effects on cardiovascular and neurodegenerative disorders: A review and meta-analysis. Int. J. Mol. Sci. 2019, 20, 351. [Google Scholar] [CrossRef] [PubMed]
- Vinet, R.; Knox, M.; Mascher, D.; Paredes-Carbajal, C.; Martinez, J.L. Isolated aorta model and its contribution to phytopharmacology. Bol. Latinoam. Caribe Plant. Med. Aromat. 2012, 11, 35–45. [Google Scholar]
- Vinet, R.; Brieva, C.; Pinardi, G.; Penna, M. Modulation of Alpha-Adrenergic-Induced Contractions by Endothelium-Derived Relaxing Factor in Rat Aorta. Gen. Pharmacol. 1991, 22, 137–142. [Google Scholar] [CrossRef]
- Hill, M.A.; Zou, H.; Potocnik, S.J.; Meininger, G.A.; Davis, M.J. Invited review: Arteriolar smooth muscle mechanotransduction: Ca(2+) signaling pathways underlying myogenic reactivity. J. Appl. Physiol. 2001, 91, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Cole, W.C.; Welsh, D.G. Role of myosin light chain kinase and myosin light chain phosphatase in the resistance arterial myogenic response to intravascular pressure. Arch. Biochem. Biophys. 2011, 510, 160–173. [Google Scholar] [CrossRef]
- Brozovich, F.V.; Nicholson, C.J.; Degen, C.V.; Gao, Y.Z.; Aggarwal, M.; Morgan, K.G. Mechanisms of Vascular Smooth Muscle Contraction and the Basis for Pharmacologic Treatment of Smooth Muscle Disorders. Pharm. Rev. 2016, 68, 476–532. [Google Scholar] [CrossRef]
- Hille, B. Ion. Channels of Excitable Membranes, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001. [Google Scholar]
- Nelson, M.T.; Quayle, J.M. Physiological roles and properties of potassium channels in arterial smooth muscle. Am. J. Physiol. Cell. Physiol. 1995, 268, C799–C822. [Google Scholar] [CrossRef]
- Martin, P.; Rebolledo, A.; Palomo, A.R.; Moncada, M.; Piccinini, L.; Milesi, V. Diversity of potassium channels in human umbilical artery smooth muscle cells: A review of their roles in human umbilical artery contraction. Reprod. Sci. 2014, 21, 432–441. [Google Scholar] [CrossRef]
- Murga, C.; Arcones, A.C.; Cruces-Sande, M.; Briones, A.M.; Salaices, M.; Mayor, F. Protein-coupled receptor kinase 2 (GRK2) as a potential therapeutic target in cardiovascular and metabolic diseases. Front. Pharmacol. 2019, 10, 112. [Google Scholar] [CrossRef]
- Obejero-Paz, C.A.; Auslender, M.; Scarpa, A. PKC activity modulates availability and long openings of L-type Ca2+ channels in A7r5 cells. Am. J. Physiol. 1998, 275, C535–C543. [Google Scholar] [CrossRef] [PubMed]
- Rapacon-Baker, M.; Zhang, F.; Pucci, M.L.; Guan, H.; Nasjletti, A. Expression of myogenic constrictor tone in the aorta of hypertensive rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R968–R975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touyz, R.M.; Alves-Lopes, R.; Rios, F.J.; Camargo, L.L.; Anagnostopoulou, A.; Arner, A.; Montezano, A.C. Vascular smooth muscle contraction in hypertension. Cardiovasc. Res. 2018, 114, 529–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, R.A.; Weisbrod, R.M.; Gericke, M.; Yaghoubi, M.; Bierl, C.; Bolotina, V.M. Mechanism of nitric oxide-induced vasodilatation: Refilling of intracellular stores by sarcoplasmic reticulum Ca2+ ATPase and inhibition of store-operated Ca2+ influx. Circ. Res. 1999, 84, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Adachi, T.; Matsui, R.; Weisbrod, R.M.; Najibi, S.; Cohen, R.A. Reduced sarco/endoplasmic reticulum Ca2+ uptake activity can account for the reduced response to NO, but not sodium nitroprusside, in hypercholesterolemic rabbit aorta. Circulation 2001, 104, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, T.J.; Bowman, P.S.; Jefferson, A.; Tosun, M.; Lynch, R.M.; Paul, R.J. Na+-K+-ATPase and Ca2+ clearance proteins in smooth muscle: A functional unit. Am. J. Physiol Heart Circ. Physiol. 2010, 299, H548–H556. [Google Scholar] [CrossRef] [PubMed]
- Feletou, M.; Kohler, R.; Vanhoutte, P.M. Nitric oxide: Orchestrator of endothelium-dependent responses. Ann. Med. 2012, 44, 694–716. [Google Scholar] [CrossRef]
- Stuehr, D.J. Structure-function aspects in the nitric oxide synthases. Annu. Rev. Pharmacol. Toxicol. 1997, 37, 339–359. [Google Scholar] [CrossRef]
- Bredt, D.S.; Hwang, P.M.; Snyder, S.H. Localization of nitric oxide synthase indicating a neural role for nitric oxide. Nature 1990, 347, 768–770. [Google Scholar] [CrossRef]
- Ferreiro, C.R.; Chagas, A.C.; Carvalho, M.H.; Dantas, A.P.; Scavone, C.; Souza, L.C.; Buffolo, E.; da Luz, P.L. Expression of inducible nitric oxide synthase is increased in patients with heart failure due to ischemic disease. Braz. J. Med. Biol. Res. 2004, 37, 1313–1320. [Google Scholar] [CrossRef] [Green Version]
- Rao, K.M. Molecular mechanisms regulating iNOS expression in various cell types. J. Toxicol. Environ. Healthb. Crit. Rev. 2000, 3, 27–58. [Google Scholar] [CrossRef] [PubMed]
- De Mey, J.G.; Vanhoutte, P.M. End o’ the line revisited: Moving on from nitric oxide to CGRP. Life Sci. 2014, 118, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Widmer, R.J.; Lerman, A. Endothelial dysfunction and cardiovascular disease. Glob. Cardiol. Sci. Pract. 2014, 43, 291–308. [Google Scholar] [CrossRef] [PubMed]
- Whitt, M.D.; Jackson, M.J. Practicality and importance of selected endothelial dysfunction measurement techniques: Review. Biomed. Eng. Lett. 2019, 9, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 20, 1523–1526. [Google Scholar] [CrossRef]
- Fitzpatrick, D.F.; Hirschfield, S.L.; Coffey, R.G. Endothelium-dependent vasorelaxing activity of wine and other grape products. Am. J. Physiol. Heart Circ. Physiol. 1993, 265, H774–H778. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.D.; Kang, S.Y.; Park, J.H.; Schini-Kerth, V.B. Ginsenoside Rg3 mediates endothelium-dependent relaxation in response to ginsenosides in rat aorta: Role of K+ channels. Eur. J. Pharmacol. 1999, 367, 41–49. [Google Scholar] [CrossRef]
- Rhyu, M.R.; Kim, J.H.; Kim, E.Y. Radix angelica elicits both nitric oxide-dependent and calcium influx-mediated relaxation in rat aorta. J. Cardiovasc. Pharmacol. 2005, 46, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, Y.; Xing, L.; Dai, X.; Liu, S.; Yu, B.; Wang, Y. Endothelium-dependent vasodilation effects of Panax notoginseng and its main components are mediated by nitric oxide and cyclooxygenase pathways. Exp. Ther. Med. 2016, 12, 3998–4006. [Google Scholar] [CrossRef] [Green Version]
- Kubota, Y.; Tanaka, N.; Umegaki, K.; Takenaka, H.; Mizuno, H.; Nakamura, K.; Shinozuka, K.; Kunitomo, M. Ginkgo biloba extract-induced relaxation of rat aorta is associated with increase in endothelial intracellular calcium level. Life Sci. 2001, 69, 2327–2336. [Google Scholar] [CrossRef]
- Abeywardena, M.; Runnie, I.; Nizar, M.; Suhaila, M.; Head, R. Polyphenol-enriched extract of oil palm fronds (Elaeis guineensis) promotes vascular relaxation via endothelium-dependent mechanisms. Asia Pac. J. Clin. Nutr. 2002, 11, S467–S472. [Google Scholar] [CrossRef] [PubMed]
- Ziyyat, A.; Mekhfi, H.; Bnouham, M.; Tahri, A.; Legssyer, A.; Hoerter, J.; Fischmeister, R. Arbutus unedo induces endothelium-dependent relaxation of the isolated rat aorta. Phytother. Res. 2002, 16, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Kwan, C.Y.; Chen, C.X.; Deyama, T.; Nishibe, S. Endothelium-dependent vasorelaxant effects of the aqueous extracts of the Eucommia ulmoides Oliv. leaf and bark: Implications on their antihypertensive action. Vasc. Pharmacol. 2003, 40, 229–235. [Google Scholar] [CrossRef]
- Lee, Y.J.; Cho, J.Y.; Kim, J.H.; Park, W.K.; Kim, D.K.; Rhyu, M.R. Extracts from Schizandra chinensis fruit activate estrogen receptors: A possible clue to its effects on nitric oxide-mediated vasorelaxation. Biol. Pharmaceut. Bull. 2004, 27, 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- Rhyu, M.R.; Kim, E.Y.; Yoon, B.K.; Lee, Y.J.; Chen, S.N. Aqueous extract of Schizandra chinensis fruit causes endothelium-dependent and -independent relaxation of isolated rat thoracic aorta. Phytomedicine 2006, 13, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Jho, H.K.; Kim, D.I.; Rhyu, M.R. Cirsium japonicum elicits endothelium-dependent relaxation via histamine H(1)-receptor in rat thoracic aorta. J. Ethnopharmacol. 2008, 116, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Korematsu, S.; Byun, E.B.; Nishizuka, T.; Ohshima, S.; Kanda, T. Apple procyanidins induced vascular relaxation in isolated rat aorta through NO/cGMP pathway in combination with hyperpolarization by multiple K+ channel activations. Biosci. Biotechnol. Biochem. 2009, 73, 2246–2251. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Qian, L.; Zhou, X.; Gao, Q.; Bruce, I.C.; Xia, Q. Endothelium-independent relaxation and contraction of rat aorta induced by ethyl acetate extract from leaves of Morus alba (L.). J. Ethnopharmacol. 2008, 120, 442–446. [Google Scholar] [CrossRef]
- Panth, N.; Paudel, K.R.; Gong, D.S.; Oak, M.H. Vascular protection by ethanol extract of Morus alba root bark: Endothelium-dependent relaxation of rat aorta and decrease of smooth muscle cell migration and proliferation. Evid. Based Complement. Alternat. Med. 2018, 2018, 7905763. [Google Scholar] [CrossRef]
- Intapad, S.; Suksamrarn, A.; Piyachaturawat, P. Enhancement of vascular relaxation in rat aorta by phytoestrogens from Curcuma comosa Roxb. Vasc. Pharmacol. 2009, 51, 284–290. [Google Scholar] [CrossRef]
- Jin, S.N.; Wen, J.F.; Kim, H.Y.; Kang, D.G.; Lee, H.S.; Cho, K.W. Vascular relaxation by ethanol extract of Xanthoceras sorbifolia via Akt- and SOCE-eNOS-cGMP pathways. J. Ethnopharmacol. 2010, 132, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Alves Filho, F.; Cavalcanti, P.M.; Passaglia, R.C.; Ballejo, G. Long-lasting endothelium-dependent relaxation of isolated arteries caused by an extract from the bark of Combretum leprosum. Einstein (Sao Paulo) 2015, 13, 395–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Getiye, Y.; Tolessa, T.; Engidawork, E. Antihypertensive activity of 80% methanol seed extract of Calpurnia aurea (Ait.) Benth. subsp. aurea (Fabaceae) is mediated through calcium antagonism induced vasodilation. J. Ethnopharmacol. 2016, 189, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Castro-Ruiz, J.E.; Rojas-Molina, A.; Luna-Vazquez, F.J.; Rivero-Cruz, F.; Garcia-Gasca, T.; Ibarra-Alvarado, C. Affinin (spilanthol), isolated from Heliopsis longipes, induces vasodilation via activation of gasotransmitters and prostacyclin signaling pathways. Int. J. Mol. Sci. 2017, 18, 218. [Google Scholar] [CrossRef] [PubMed]
- Assaidi, A.; Dib, I.; Tits, M.; Angenot, L.; Bellahcen, S.; Bouanani, N.; Legssyer, A.; Aziz, M.; Mekhfi, H.; Bnouham, M.; et al. Chenopodium ambrosioides induces an endothelium-dependent relaxation of rat isolated aorta. J. Integr. Med. 2019, 17, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Vinet, R.; Cortes, M.; Alvarez, R.; Guzman, L.; Flores, E. Centaurium cachanlahuen (Mol.) Robinson, a chilean native plant with a vasodilatory effect. Bol. Latinoam. Caribe Plant. Med. Aromat. 2012, 11, 61–65. [Google Scholar]
- Vinet, R.; Alvarez, R.; Knox, M.; Guzman, L.; Martinez, J.L.; Flores, E. Vasodilatory properties of Solanum crispum Ruiz & Pav. a South American native plant. Bol. Latinoam. Caribe Plant. Med. Aromat. 2016, 15, 94–98. [Google Scholar]
- Razavi, B.M.; Alyasin, A.; Hosseinzadeh, H.; Imenshahidi, M. Saffron induced relaxation in isolated rat aorta via endothelium dependent and independent mechanisms. Iran. J. Pharm. Res. 2018, 17, 1018–1025. [Google Scholar] [CrossRef]
- Kukula-Koch, W.A.; Widelski, J. Alkaloids. In Pharmacognosy; Delgoda, R., Ed.; Academic Press: Boston, MA, USA, 2017; pp. 163–198. [Google Scholar] [CrossRef]
- Wong, K.K. Differential effect of tetrandrine on aortic relaxation and chronotropic activity in rat isolated aorta and atria. Planta Med. 1998, 64, 663–665. [Google Scholar] [CrossRef]
- Wong, K.K. Mechanism of the aortic relaxation induced by low concentrations of berberine. Planta Med. 1998, 64, 756–757. [Google Scholar] [CrossRef]
- Tur, R.; Magraner, J.; Catret, M.; Elorriaga, M.; Ivorra, M.D.; D’Ocon, P.; Bermejo, A.; Cabedo, N.; Cortes, D.; Anselmi, E. Mechanism of vascular relaxation by thaligrisine: Functional and binding assays. Life Sci. 2000, 67, 1535–1548. [Google Scholar] [CrossRef]
- Mitamura, M.; Horie, S.; Sakaguchi, M.; Someya, A.; Tsuchiya, S.; Van de Voorde, J.; Murayama, T.; Watanabe, K. Mesaconitine-induced relaxation in rat aorta: Involvement of Ca2+ influx and nitric-oxide synthase in the endothelium. Eur. J. Pharmacol. 2002, 436, 217–225. [Google Scholar] [CrossRef]
- Martinez, J.L.; Benites, J.; Gomez, H.; Vargas-Rueda, S.; Vinet, R.; Jaimes, L.; Laurido, C. Effects of two bisbenzylisoquinoline alkaloids, antioquine and tetrandrine, compared to verapamil in rat thoracic aorta. An. Acad. Bras. Cienc. 2018, 90, 1459–1463. [Google Scholar] [CrossRef]
- Prieto, J.; Cortes, D.; Jaimes, L.; Laurido, C.; Vinet, R.; Martinez, J.L. Pseudoxandra sclerocarpa Maas, Colombian medicinal plant: A review. Bol. Latinoam. Caribe Plant. Med. Aromat. 2015, 14, 308–316. [Google Scholar]
- Cifuentes, F.; Palacios, J.; Paredes, A.; Nwokocha, C.R.; Paz, C. 8-oxo-9-dihydromakomakine isolated from Aristotelia chilensis induces vasodilation in rat aorta: Role of the extracellular calcium influx. Molecules 2018, 23, 3050. [Google Scholar] [CrossRef]
- Zhang, P.Y. Polyphenols in health and disease. Cell Biochem. Biophys. 2015, 73, 649–664. [Google Scholar] [CrossRef]
- Chen, C.K.; Pace-Asciak, C.R. Vasorelaxing activity of resveratrol and quercetin in isolated rat aorta. Gen. Pharmacol. 1996, 27, 363–366. [Google Scholar] [CrossRef]
- Chan, E.C.; Pannangpetch, P.; Woodman, O.L. Relaxation to flavones and flavonols in rat isolated thoracic aorta: Mechanism of action and structure-activity relationships. J. Cardiovasc. Pharmacol. 2000, 35, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.G.; Oropeza, M.V.; Villanueva, T.; Aguilar, M.I.; Delgado, G.; Ponce, H.A. Xanthorrhizol induces endothelium-independent relaxation of rat thoracic aorta. Life Sci. 2000, 67, 327–333. [Google Scholar] [CrossRef]
- Campos, M.; Oropeza, M.; Ponce, H.; Fernandez, J.; Jimenez-Estrada, M.; Torres, H.; Reyes-Chilpa, R. Relaxation of uterine and aortic smooth muscle by glaucolides D and E from Vernonia liatroides. Biol. Pharm. Bull. 2003, 26, 112–115. [Google Scholar] [CrossRef]
- Chan, S.S.; Cheng, T.Y.; Lin, G. Relaxation effects of ligustilide and senkyunolide A, two main constituents of Ligusticum chuanxiong, in rat isolated aorta. J. Ethnopharmacol. 2007, 111, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.G.; Choi, D.H.; Lee, J.K.; Lee, Y.J.; Moon, M.K.; Yang, S.N.; Kwon, T.O.; Kwon, J.W.; Kim, J.S.; Lee, H.S. Endothelial NO/cGMP-dependent vascular relaxation of cornuside isolated from the fruit of Cornus officinalis. Planta Med. 2007, 73, 1436–1440. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Lee, S.J.; Yun, M.R.; Seo, K.W.; Bae, S.S.; Park, J.W.; Lee, Y.J.; Shin, W.J.; Choi, Y.W.; Kim, C.D. Gomisin A from Schisandra chinensis induces endothelium-dependent and direct relaxation in rat thoracic aorta. Planta Med. 2007, 73, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Feng, T.; Shan, L.; Cai, B.; Chu, W.; Niu, H.; Lu, Y.; Yang, B. Scutellarin-induced endothelium-independent relaxation in rat aorta. Phytother. Res. 2008, 22, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Assreuy, A.M.; Pinto, N.V.; Lima Mota, M.R.; Passos Meireles, A.V.; Cajazeiras, J.B.; Nobre, C.B.; Soares, P.M.; Cavada, B.S. Vascular smooth muscle relaxation by a lectin from Pisum arvense: Evidences of endothelial NOS pathway. Protein Pept. Lett. 2011, 18, 1107–1111. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Hou, X.; Zhang, M.; Liang, T.; Zhi, J.; Han, L.; Li, Q. Relaxation of rat aorta by farrerol correlates with potency to reduce intracellular calcium of VSMCs. Int. J. Mol. Sci. 2014, 15, 6641–6656. [Google Scholar] [CrossRef] [PubMed]
- Vinet, R.; Araos, P.; Gentina, J.C.; Knox, M.; Guzman, L. p-Coumaric acid reduces high glucose-mediated impairment of endothelium-dependent relaxation in rat aorta. Bol. Latinoam. Caribe Plant. Med. Aromat. 2014, 13, 232–237. [Google Scholar]
- Yan, Y.; Chen, Y.C.; Lin, Y.H.; Guo, J.; Niu, Z.R.; Li, L.; Wang, S.B.; Fang, L.H.; Du, G.H. Brazilin isolated rom the heartwood of Caesalpinia sappan L induces endothelium-dependent and -independent relaxation of rat aortic rings. Acta Pharmacol. Sin. 2015, 36, 1318–1326. [Google Scholar] [CrossRef] [PubMed]
- Latif, R.; Badar, A. Role of various potassium channels in caffeine-induced aortic relaxation in rats. Saudi J. Med. Sci. 2016, 4, 197–201. [Google Scholar] [CrossRef]

{kind=link}
| Plant Extracts | Endothelium Participation | Postulated Mechanism | Reference |
|---|---|---|---|
| Vitis vinifera (grape products) | EDR | NOS/cGMP pathway | [38] |
| Panax ginseng (ginsenosides) | EDR | NOS/TEA-sensitive K+ channels | [39] |
| Radix angelica (aqueous and butanol fraction) | EDR and EIR | NOS pathway | [40] |
| Panax notoginseng (saponins) | EDR | NOS and COX pathways | [41] |
| Ginkgo biloba | EDR | NOS pathway | [42] |
| E. guineensis, C. citrates, C. papaya, and C. frutescens (Extracts enriched in polyphenols) | EDR | [43] | |
| Arbutus unedo (aqueous extract) | EDR | NOS pathway | [44] |
| Eucommia ulmoides (aqueous extracts) | EDR | NOS pathway | [45] |
| Schizandra chinensis (aqueous extract) | EDR | NOS pathway/via estrogen receptors | [47] |
| Cirsium japonicum (aqueous extract) | EDR | NOS/cGMP pathway/via histamine H1-receptors | [48] |
| Malus pumila (apple procyanidins) | EDR | NOS/cGMP pathway | [49] |
| Morus alba (ethyl acetate extract from leaves) | EIR | Inhibition of voltage- and receptor-dependent Ca2+ channels in VSMCs | [50] |
| Morus alba (ethanol extract from root bark) | EDR | NOS pathway/K+ channel activation | [51] |
| Curcuma comosa (hexane extract) | Increase ACh-induced EDR | Estrogen receptor/NO-cGMP-dependent mechanisms and ER-Akt-eNOS pathway | [52] |
| Xanthoceras sorbifolia (ethanol extract) | EDR | NOS-cGMP pathways and Akt- and SOCE-eNOS-sGC pathways | [53] |
| Combretum leprosum (ethanol extract) | EDR | Ca2+-dependent stimulation of ECs | [54] |
| Calpurnia aurea (methanol extract) | EIR | It is suggested VDCCs participation. | [55] |
| Heliopsis longipes (ethanolic extract) | EDR | It is suggested gasotransmitters and prostacyclin participation. | [56] |
| Chenopodium ambrosioides (methanolic extracts) | EDR | Stimulation of muscarinic receptors in ECs | [57] |
| Centaurium cachanlahuen (aqueous and hydroalcoholic extracts) | EDR | NOS pathway | [58] |
| Solanum crispum (hydroalcoholic extract) | EDR | NOS pathway | [59] |
| Crocus sativus (aqueous extract) | EDR and EIR | NOS pathway and VDCCs in VSMCs | [60] |
| Alkaloids | Endothelium Participation | Postulated Mechanism | Reference |
|---|---|---|---|
| Tetrandrine (bisbenzylisoquinoline alkaloid) | EIR | Blocked VDCCs in VSMCs | [62] |
| Berberine (benzylisoquinoline alkaloid) | EDR/EIR | EDR (low concentrations) EIR (high concentrations) | [63] |
| Thaligrisine (bisbenzyl tetrahydroisoquinoline alkaloid) | EIR | Blocked VDCCs in VSMCs | [64] |
| Mesaconitine (diterpene alkaloid) | EIR | Blocked VDCCs in VSMCs | [65] |
| Antioquine (bisbenzylisoquinoline alkaloids) | EIR | Blocked VDCCs in VSMCs | [66] |
| 8-Oxo-9-dihydromakomakine (isolated from Aristotelia chilensis) | EIR | Blocked VDCCs in VSMCs | [68] |
| Non-Alkaloid Compounds | Endothelium Participation | Postulated Mechanism | Reference |
|---|---|---|---|
| Resveratrol and quercetin (polyphenols) | EDR and EIR | NOS (low concentrations) and non-NOS (high concentrations) pathways. | [70] |
| Flavonols (fisetin, quercetin, and 3,3’,4’-trihydroxyflavone) and flavones (apigenin, chrysin, and luteolin) | EDR and EIR | NOS (flavonols) and non-NOS (flavones) pathways. | [71] |
| Xanthorrhizol (sesquiterpene) | EIR | Inhibiting voltage/receptor-gated Ca2+ channels. | [72] |
| Glaucolides D and E | EIR | Inhibiting voltage/receptor-gated Ca2+ channels. | [73] |
| Ligustilide and senkyunolide A (phthalide derivatives) | EIR | [74] | |
| Cornuside (bisiridoid glucoside) | EDR | NOS/cGMP pathways | [75] |
| Gomisin A (lignan) | EDR | NOS pathway. | [76] |
| Scutellarin (flavonoid) | EIR | Blocked Ca2+ entry independent of VDCCs. | [77] |
| Lectin (seed lectin of P. arvense) | EDR | NOS pathway. | [78] |
| Farrerol (flavanone) | EIR | Blocked VDCCs in VSMCs | [79] |
| p-Coumaric acid | EDR | NOS pathway. | [80] |
| Brazilin (heterotetracyclic) | EDR and EIR | Blocked VDCCs in VSMCs | [81] |
| Caffeine | EDR and EIR | Voltage-dependent K+ channels. | [82] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knox, M.; Vinet, R.; Fuentes, L.; Morales, B.; Martínez, J.L. A Review of Endothelium-Dependent and -Independent Vasodilation Induced by Phytochemicals in Isolated Rat Aorta. Animals 2019, 9, 623. https://doi.org/10.3390/ani9090623
Knox M, Vinet R, Fuentes L, Morales B, Martínez JL. A Review of Endothelium-Dependent and -Independent Vasodilation Induced by Phytochemicals in Isolated Rat Aorta. Animals. 2019; 9(9):623. https://doi.org/10.3390/ani9090623
Chicago/Turabian StyleKnox, Marcela, Raúl Vinet, Lida Fuentes, Bernardo Morales, and José L. Martínez. 2019. "A Review of Endothelium-Dependent and -Independent Vasodilation Induced by Phytochemicals in Isolated Rat Aorta" Animals 9, no. 9: 623. https://doi.org/10.3390/ani9090623
APA StyleKnox, M., Vinet, R., Fuentes, L., Morales, B., & Martínez, J. L. (2019). A Review of Endothelium-Dependent and -Independent Vasodilation Induced by Phytochemicals in Isolated Rat Aorta. Animals, 9(9), 623. https://doi.org/10.3390/ani9090623