1. Introduction
High temperatures negatively affect protein utilization efficiency [
1]. In addition, diets that contain a high content of dietary crude protein (CP) will increase internal heat production by the elevated heat increment in a high-temperature climate [
2]. Therefore, low levels of CP with supplementing limiting amino acids can overcome the bad effects of heat stress [
3,
4] and enhance protein utilization [
5,
6]. Recently, it has been reported that laying performance was equal among 14%, 15%, and 16% dietary CP [
7]. Since synthetic dietary L-Thr became commercially available, it is possible to decrease CP. Thr affects protein synthesis and is the third limiting amino acid [
8,
9]. Recently, it has been reported that Thr is a limiting amino acid in diets containing 14% CP [
10].
It has been showed that heat stress provoked lipid accumulation by elevated de novo lipogenesis, decreased lipolysis, and enhanced amino acid catabolism [
11]. In addition, during stress times, the bird’s body begins freeing heat shock proteins to secure itself from the harmful cellular effects of reactive oxygen species [
12]. Heat stress is usually accompanied by increasing levels of 70 kilodalton heat shock proteins (HSP70) [
13,
14]. In addition, high temperatures disturb oxidative status [
15] and increase serum total cholesterol, triglyceride, and zinc [
16,
17,
18,
19,
20].
The present research aimed to estimate the dietary Thr requirement in low CP diets for laying hens in a high-temperature environmental climate. In addition, the effects of increasing Thr on lipid peroxidation, antioxidants enzymes activities, mineral levels, and HSP70 were investigated.
2. Material and Methods
2.1. Management
All procedures in this study were conducted according to the guide for the care and use of agricultural animals in research and teaching (American society of animal science and poultry science association, 2010), through research group (No. RG-1440-146).
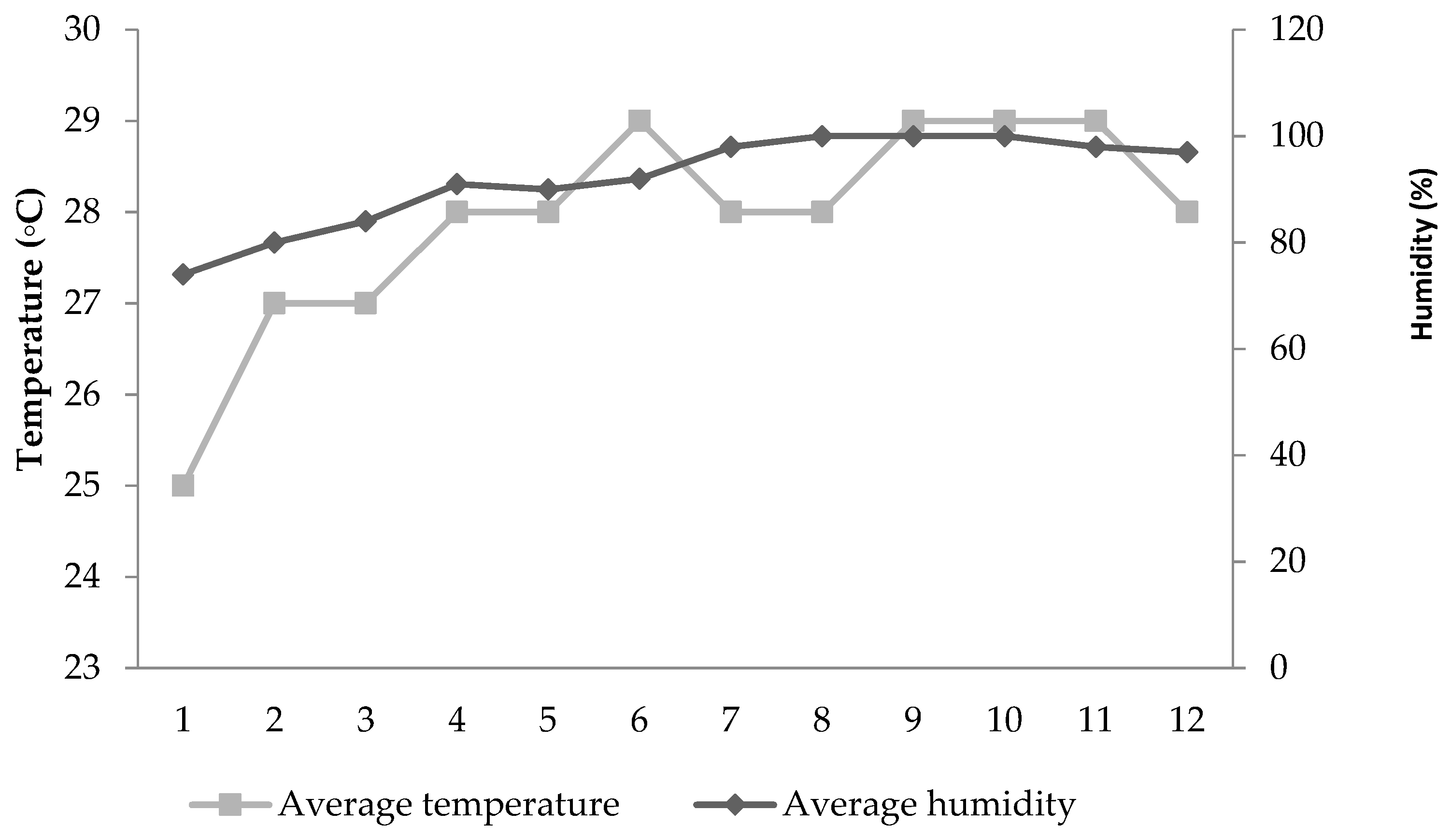
Lohmann Brown hens (n = 420), at 28 weeks of age, and with almost similar live body weights (1800 g), were divided into five dietary treatments. Each treatment included six replicates of 14 laying hens (4 birds/cage; 471.5 cm
2/hen). They were exposed to 16-h light. The study began during the middle of May and ended in August; it lasted 13 weeks, including one week for acclimation. The mean daily temperature and humidity are both presented in
Figure 1.
2.2. Experimental Diets
Hens were fed
adlibitum (mash form), and water was available through nipples. Dietary CP (14%) was presented as the control diet (
Table 1). Dietary Thr levels are 0.43%, 0.49%, 0.57%, 0.66%, and 0.74%, based on digestible base. Ingredient and analyzed CP and total amino acids are presented in
Table 1 and
Table 2, respectively. L-Thr (98.5% purity) was supplied at the expense of kaolin (inert filler). Samples from each diet were analyzed for CP and amino acids according to [
21]. Total amino acids in diets were analyzed using HPLC, as described by the authors of [
10].
2.3. Laying Performance
Mortalities were recorded daily. Egg numbers and egg weight were recorded daily. However, feed consumption was recorded weekly. Egg mass was calculated according to this formula (egg weight × egg production), while feed conversion ratio (FCR) was calculated according to grams of feed consumption/grams of egg mass produced.
2.4. Blood, Liver, and Ileum Sampling and Laboratory Analyses
At the end of the trial (40 weeks), 6 hens per treatment were slaughtered. The blood was collected and was centrifuged (3000× g) for 10 min. It was aspirated by pipette and stored in Eppendorf tubes at −70 °C. Serum concentrations of zinc (Zn), copper (Cu), triglyceride (TG), total cholesterol (CHO), glutamic oxaloacetic transaminase (GOT), and glutamic pyruvic transaminase (GPT) were determined by kits from (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The levels of serum uric acid, high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C) were analyzed by commercial kits from the previous company. The HDL-C levels in serum supernatant were determined after precipitation of lipoprotein-B using phosphotungstic acid/Mg2+ (PTA/Mg2+).
Serum 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase was determined following the manufacturer’s protocols from Sigma (St. Louis, MO, USA).
After slaughter, the liver of each hen was collected immediately, snap-frozen with liquid nitrogen, and stored at −80 °C until analysis. About 0.5 g of liver of every bird was homogenized and analyzed for CHO, TG, GOT, and GPT, as described above in the serum.
2.5. Oxidant and Antioxidant Status
Serum levels malondialdehyde (MDA), superoxide dismutase (SOD), total antioxidative capability (T-AOC), and copper zinc superoxide dismutase (CuZn-SOD) were analyzed as described by [
22,
23]. Liver tissues were homogenized in ice-cold isotonic physiological saline to form homogenates at the concentration of 0.1 g liver/mL. The samples of liver tissues were homogenized and centrifuged, and the supernatants were collected to analyze MDA, T-AOC, and SOD.
2.6. HSP70 mRNA Expression Assay
Total RNA was isolated from 50 mg of ileum, according to the instructions (TRIzol; Invitrogen, Carlsbad, CA, USA). The quality of RNA was examined by both native RNA electrophoresis on 1.0% agarose gel and the UV absorbance at 260 nm and 280 nm. The cDNA was synthesized from 2 lg of total RNA by a reverse transcriptase at 42 °C for 60 min with oligo dT-adaptor primer, using the protocol of the manufacturer (M-MLV; Takara, Dalian, China). The abundance of mRNA was determined based on a Step-One-Plus Real-Time PCR (ABI 7500; Applied Biosystems, Foster, CA, USA). The PCR used a kit (SYBR Premix PCR kit; Takara, Dalian, China) as described by [
10]. Average gene expression relative to the endogenous control for each sample was calculated using the 2
−ΔΔCt method [
24]. The calibrator for each studied gene was the average
ΔCt value of the control group (
Table 3).
2.7. Statistical Analyses
Data (Replicate; n = 6) were statistically analyzed by one-way ANOVA (SPSS Inc., Chicago, IL, USA). Polynomial comparisons were applied to test for linear and quadratic responses of dependent variables to dietary Thr. Inflection points in response curves at increasing dietary Thr levels were calculated following [
28]. To estimate the optimal Thr requirement, a quadratic regression equation based on 95% of the maximum or minimum response was used [
29,
30].
4. Discussion
It is important to formulate accurate diets to meet the requirements of laying hens because feed ingredients are expensive. In addition, laying hens have been selected for massive egg production, resulting in greater metabolic activity and reduced thermo-tolerance [
31,
32]. Heat stress has adverse effects on laying hens [
33,
34,
35]. In addition, high temperatures increase the hens’ discomfort and lead to behavioral and endocrinological changes.
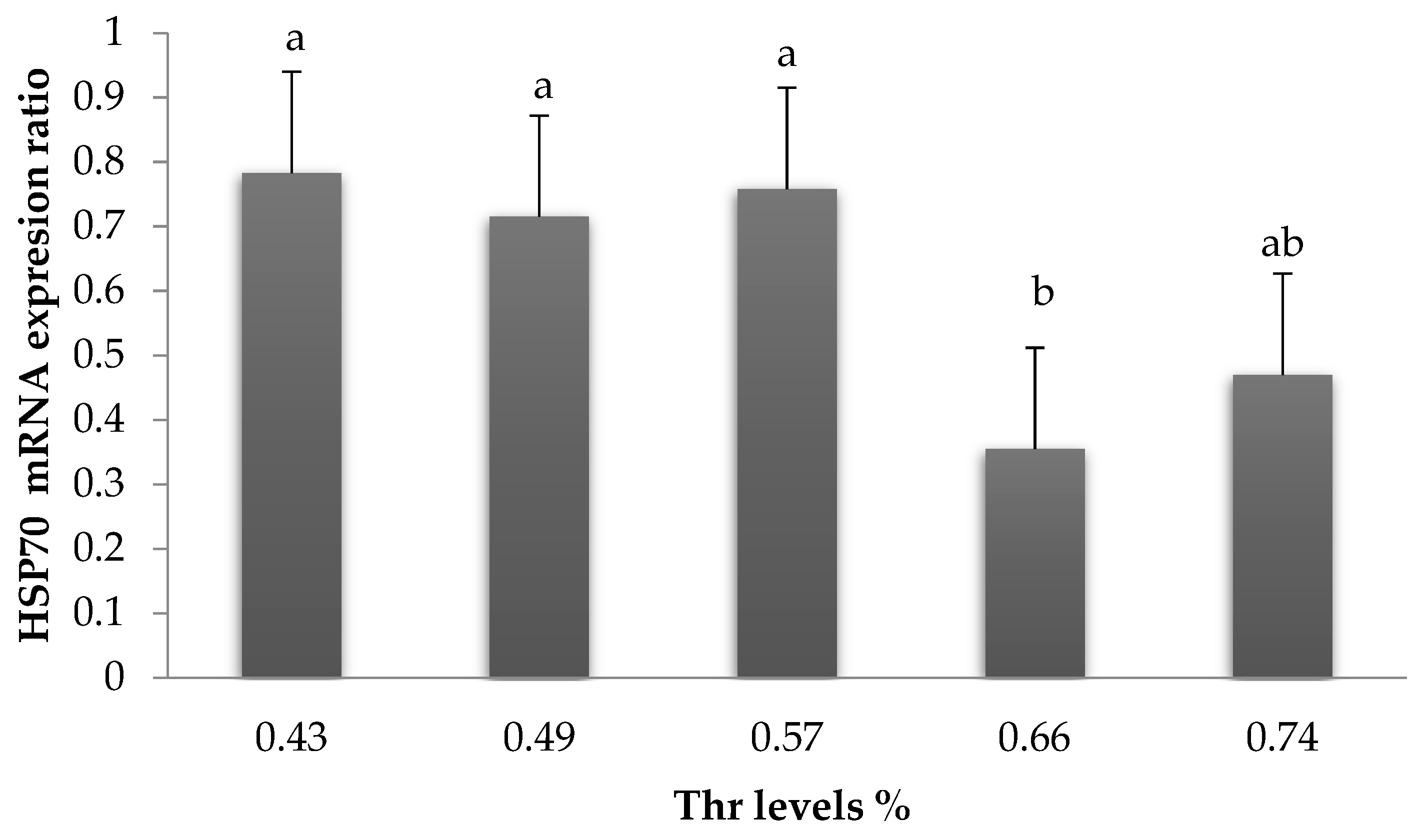
In the present study, rapid panting was noticed. In addition, the expression of ileal HSP70 protein decreased (
p < 0.05) at 0.66% Thr. It has been reported that heat shock protein protects birds from high temperatures by preventing unwanted protein aggregation and channelizing their degradation [
36]. The expression of mRNA HSP70 was measured in the gut [
14,
37], liver [
38], hypothalamus [
27,
39], and blood and feather [
40]. Here, we focused on detecting HSP70 in ileum because it plays a vital role in digestion and absorption, as well as immunity. In addition, the effects of Thr on intestinal function were known [
41,
42], and the effects of heat stress on gut function become obviously clear.
Both serum CHO and serum 3-HMG-CoA reductase decreased significantly (
p < 0.05) at 0.66% dietary Thr. It has been reported that 3-hydroxy-3-methylglutaric acid (HMG) is a potent agent for reducing serum triglyceride and cholesterol concentrations [
43,
44]. It has been reported that HMG causes a 40% to 50% reduction of [1–
14C] acetate incorporation into cholesterol in male rats [
45]. In addition, HMG inhibited fatty acid synthesis in vivo [
46]. In vitro, HMG inhibited 3-HMG-CoA reductase [mevalonate: NADP oxidoreductase (CoA-acylating), EC 1.1.1.34] and interfered with the enzymatic steps involved in the conversion of acetate to HMG-CoA [
46]. Taken together, the data suggest that dietary Thr level affect CHO, especially biochemical pathways in which HMG-CoA reductase is involved. Our findings are in agreement with the results of the previous study in broiler chickens [
47]. They found that plasma CHO levels decreased significantly (
p < 0.05) when dietary Thr was sufficient [
47]. Here, we did not find a decrease in total CHO levels in the liver. Recently, it has been reported that Thr supplementation did not have an effect (
p > 0.05) on hepatic cholesterol in Pekin ducks [
48]. They suggested that dietary Thr supplementation enhanced hepatic lipid metabolism by regulating lipid synthesis, transport, and oxidation. It has been reported that there is no relationship between the plasma CHO level and the level of yolk cholesterol [
49,
50,
51], and, consequently, liver CHO.
The levels of GOT and GPT did not change due to treatments. The enzymatic activity of GOT and GPT are indicators of liver health. These enzymes are elevated in acute hepatotoxicity, but they are decreased with prolonged intoxication [
52].
In the present study, dietary Thr at 0.49% increased serum levels of T-SOD (
p < 0.05). In addition, dietary Thr at 0.49%, 0.57%, and 0.66% increased the levels of CuZn-superoxide dismutase (Cu-ZnSOD). The present result suggests that Thr may promote the antioxidative ability of laying hens. Previous studies [
22,
53] also found that supplemental amino acids (L-Thr and L-Trp) increased T-SOD in serum and the liver.
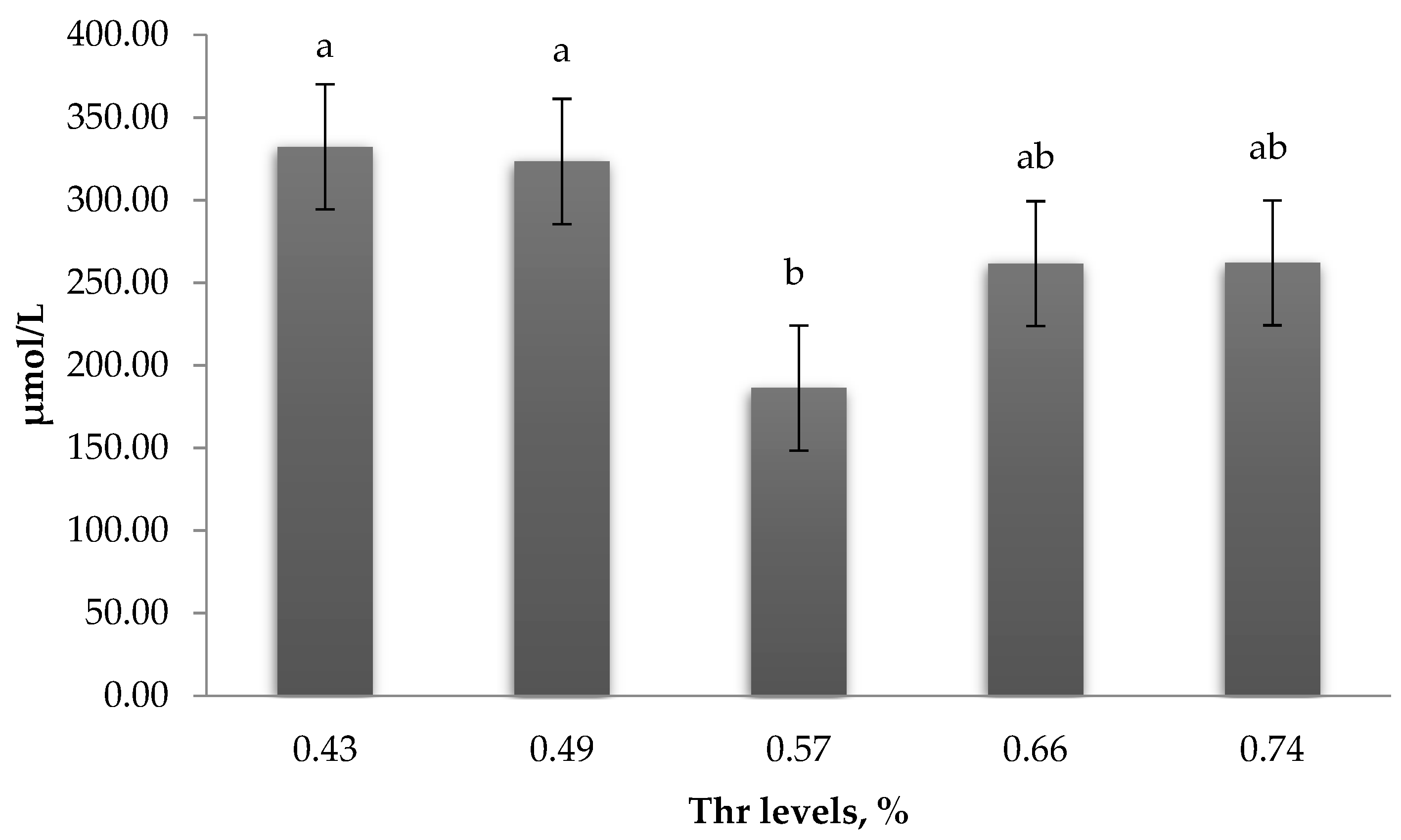
Uric acid is the metabolic product of protein metabolism and has been suggested as a dominant scavenger of free radicals [
54]. We found that level of serum uric acid was declined at 0.57% Thr, which confirms that sufficient Thr increases amino acid utilization. Thr is considered a limiting amino acid in low CP diets [
55,
56], affecting utilization of TSAA and Lys [
57]. It has been reported that the levels of plasma uric acid and excreta were higher from increasing CP than from lowering CP in the diet [
58]. In addition, a decrease in the level of uric acid excretion was reported with supplementing limiting amino acids, which indicated better N utilization [
59,
60]. The plasma urea nitrogen and uric acid have been used to estimate amino acids requirements in swine and broilers [
61,
62,
63].
It has been reported that laying performance decreased by feeding low CP diets [
64]. Here, reduction CP (14%) in the control group reduced egg production quadratically. The effect of the low crude protein diet was pronounced clearly during the late cycle of laying production (43–63 weeks of age) [
65]. This study was conducted during the first cycle of egg production (28–40 weeks).
Increasing dietary Thr to 0.57% improved egg production quadratically. It has been found that egg mass and hen-day egg production were reduced (
p < 0.05) by feeding hens a Thr deficient diet [
66]. This means that increasing recognition of Thr as a critical amino acid in the diet of laying hens fed a low CP diet under high-temperature environmental climate. The current results showed that 0.58% dietary Thr based on quadratic regressions guaranteed the best egg production. The present results are in agreement with [
67]. They estimated that Thr requirement was 0.57% of dietary Thr from 24 to 40 weeks in Hy-Line W36.
Egg weight, feed consumption, and FCR were similar among dietary Thr levels. Previous studies reported no effect of Thr levels on the egg weight and FCR in laying hens [
68,
69,
70,
71]. It has been indicated that total Thr deficiency beyond 0.42% decreased feed intake [
72].

 ,
, 




{kind=link}
{kind=link}
{kind=link}