The Blood and Muscle Expression Pattern of the Equine TCAP Gene during the Race Track Training of Arabian Horses

 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
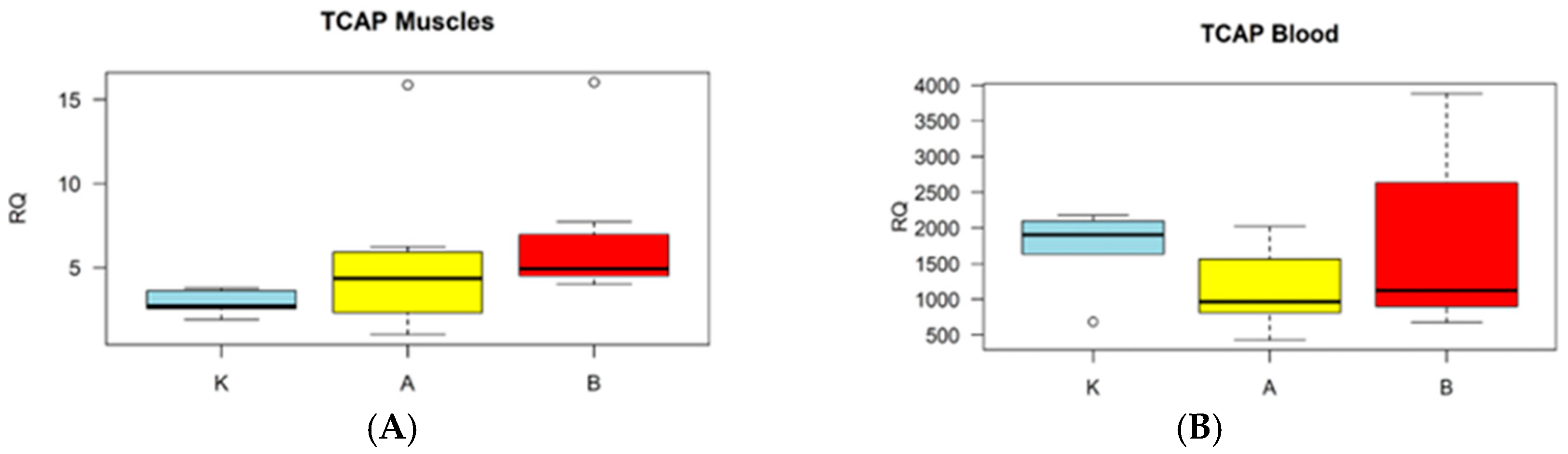
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stefaniuk, M.; Ropka-Molik, K. RNA sequencing as a powerful tool in searching for genes influencing health and performance traits of horses. J. Appl. Genet. 2016, 57, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.W.; Gu, J.; Eivers, S.S.; Fonseca, R.G.; McGivney, B.A.; Govindarajan, P.; Orr, N.; Katz, L.M.; MacHugh, D.E. A sequence polymorphism in MSTN predicts sprinting ability and racing stamina in thoroughbred horses. PLoS ONE 2010, 5, e8645. [Google Scholar] [CrossRef]
- Andersson, L.S.; Larhammar, M.; Memic, F.; Wootz, H.; Schwochow, D.; Rubin, C.J.; Patra, K.; Arnason, T.; Wellbring, L.; Hjälm, G.; et al. Mutations in DMRT3 affect locomotion in horses and spinal circuit function in mice. Nature 2012, 488, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Ropka-Molik, K.; Stefaniuk-Szmukier, M.; Żukowski, K.; Piórkowska, K.; Gurgul, A.; Bugno-Poniewierska, M. Transcriptome profiling of Arabian horse blood during training regimens. BMC Genet. 2017, 18, 31. [Google Scholar] [CrossRef] [PubMed]
- Ropka-Molik, K.; Stefaniuk-Szmukier, M.; Żukowski, K.; Piórkowska, K.; Bugno-Poniewierska, M. Exercise-induced modification of the skeletal muscle transcriptome in Arabian horses. Physiol. Genom. 2017, 149, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Ropka-Molik, K.; Stefaniuk-Szmukier, M.; Piórkowska, K.; Żukowski, K.; Bugno-Poniewierska, M. Molecular characterization of the apoptosis-related SH3RF1 and SH3RF2 genes and their association with exercise performance in Arabian horses. BMC Vet. Res. 2018, 14, 237. [Google Scholar] [CrossRef] [PubMed]
- Ropka-Molik, K.; Stefaniuk-Szmukier, M.; Musiał, A.D.; Piórkowska, K.; Szmatoła, T. Sequence analysis and expression profiling of the equine ACTN3 gene during exercise in Arabian horses. Gene 2019, 685, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Ropka-Molik, K.; Stefaniuk-Szmukier, M.; Szmatoła, T.; Piórkowska, K.; Bugno-Poniewierska, M. The use of the SLC16A1 gene as a potential marker to predict race performance in Arabian horses. BMC Genet. 2019, in press. [Google Scholar]
- Gurgul, A.; Jasielczuk, I.; Semik-Gurgul, E.; Pawlina-Tyszko, K.; Stefaniuk-Szmukier, M.; Szmatoła, T.; Polak, G.; Tomczyk-Wrona, I.; Bugno-Poniewierska, M. A genome-wide scan for diversifying selection signatures in selected horse breeds. PLoS ONE 2019, 14, e0210751. [Google Scholar] [CrossRef]
- Gunn, H.M. Muscle, bone and fat proportions and the muscle distribution of Thoroughbreds and other horses. In Equine Exercise Physiology 2; ICEEP Publications: Davis, CA, USA, 1987; pp. 253–264. [Google Scholar]
- Crook, T.C.; Cruickshank, S.E.; McGowan, C.M.; Stubbs, N.; Wakeling, J.M.; Wilson, A.M.; Payne, R.C. Comparative anatomy and muscle architecture of selected hind limb muscles in the Quarter Horse and Arab. J. Anat. 2008, 212, 144–152. [Google Scholar] [CrossRef]
- Kawai, M.; Minami, Y.; Sayama, Y.; Kuwano, A.; Hiraga, A.; Miyata, H. Muscle fiber population and biochemical properties of whole body muscles in Thoroughbred horses. Anat. Rec. 2009, 292, 1663–1669. [Google Scholar] [CrossRef]
- Jürimäe, J.; Abernethy, P.J.; Blake, K.; McEniery, M.T. Changes in the myosin heavy chain isoform profile of the triceps brachii muscle following 12 weeks of resistance training. Eur. J. App. Physiol. Occup. Physiol. 1996, 74, 287–292. [Google Scholar] [CrossRef]
- Rivero, J.L.; Ruz, A.; Martí-Korff, S.; Estepa, J.C.; Aguilera-Tejero, E.; Werkman, J.; Sobotta, M.; Lindner, A. Effects of intensity and duration of exercise on muscular responses to training of thoroughbred racehorses. J. Appl. Physiol. 1985, 102, 1871–1882. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.L.; Klueber, K.M. Skeletal muscle following tonic overload: Functional and structural analysis. Med. Sci. Sports Exerc. 1991, 23, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Pinotsis, N.; Lange, S.; Song, Y.H.; Popov, A.; Mavridis, I.; Mayans, O.M.; Gautel, M.; Wilmanns, M. Palindromic assembly of the giant muscle protein titin in the sarcomeric Z-disk. Nature 2006, 439, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Osio, A.; Tan, L.; Chen, S.N.; Lombardi, R.; Nagueh, S.F.; Shete, S.; Roberts, R.; Willerson, J.T.; Marian, A.J. Myozenin 2 is a novel gene for human hypertrophic cardiomyopathy. Circ. Res. 2007, 100, 766–768. [Google Scholar] [CrossRef] [PubMed]
- Faulkne, R.G.; Pallavicini, A.; Comelli, A.; Salamon, M.; Bortoletto, G.; Ievolella, C.; Trevisan, S.; Kojic, S.; Vecchia, F.D.; Laveder, P.; et al. FATZ, a filamin-, actinin-, and telethonin-binding protein of the Z-disc of skeletal muscle. J. Biol. Chem. 2000, 275, 41234–41242. [Google Scholar] [CrossRef] [PubMed]
- Kojic, S.; Medeot, E.; Guccione, E.; Krmac, H.; Zara, I.; Martinelli, V.; Valle, G.; Faulkner, G. The Ankrd2 protein, a link between the sarcomere and the nucleus in skeletal muscle. J. Mol. Biol. 2004, 339, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Stefaniuk, M.; Ropka-Molik, K.; Piórkowska, K.; Bereta, A.; Szpar, P.; Czerwonka, Z.; Podstawski, Z. Evaluation of minimally invasive muscle biopsy method for genetic analysis in horse. Ann. Anim. Sci. 2015, 15, 621–627. [Google Scholar] [CrossRef]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques 1993, 15, 532–537. [Google Scholar] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008; Available online: http://www.R-project.org (accessed on 17 March 2019).
- Huxley, H.; Hanson, J. Changes in the cross-striations of muscle during contraction and stretch and their structural interpretation. Nature 1954, 173, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Herzog, W. The multiple roles of titin in muscle contraction and force production. Biophys. Rev. 2018, 10, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, C.C.; Trombitás, K.; Centner, T.; Kolmerer, B.; Stier, G.; Kunke, K.; Suzuki, K.; Obermayr, F.; Herrmann, B.; Granzier, H.; et al. The NH2 terminus of titin spans the Z-disc: Its interaction with a novel 19-kD ligand (T-cap) is required for sarcomeric integrity. J. Cell. Biol. 1998, 143, 1013–1027. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Gao, M.; Pinotsis, N.; Wilmanns, M.; Schulten, K. Mechanical strength of the titin Z1Z2-telethonin complex. Structure 2006, 3, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Knoll, S.; Fürst, K.; Thomas, S.; Villanueva Baselga, S.; Stoll, A.; Schaefer, S.; Pützer, B.M. Dissection of cell context-dependent interactions between HBx and p53 family members in regulation of apoptosis: A role for HBV-induced HCC. Cell Cycle 2011, 10, 3554–3565. [Google Scholar] [CrossRef] [PubMed]
- Dahlqvist, J.R.; Voss, L.G.; Lauridsen, T.; Krag, T.O.; Vissing, J. A pilot study of muscle plasma protein changes after exercise. Muscle Nerve 2014, 49, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yang, J.; Zhu, J.; Xu, X. Depletion of zebrafish Tcap leads to muscular dystrophy via disrupting sarcomere-membrane interaction, not sarcomere assembly. Hum. Mol. Genet. 2009, 18, 4130–4140. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, M.; Horita, Z.; Nemoto, M.; Langdon, T.G. Processing of metals by equal-channel angular pressing. J. Mate. Sci. 2001, 36, 2835–2843. [Google Scholar] [CrossRef]
- Nicholas, G.; Thomas, M.; Langley, B.; Somers, W.; Patel, K.; Kemp, C.F.; Sharma, M.; Kambadur, R. Titin-cap associates with, and regulates secretion of, Myostatin. J. Cell Physiol. 2002, 1, 120–131. [Google Scholar] [CrossRef]
- Vega, R.B.; Konhilas, J.P.; Kelly, D.P.; Leinwand, L.A. Molecular Mechanisms Underlying Cardiac Adaptation to Exercise. Cell Metab. 2017, 25, 1012–1026. [Google Scholar] [CrossRef] [PubMed]
- Frey, N.; Olson, E.N. Calsarcin-3, a novel skeletal muscle-specific member of the calsarcin family, interacts with multiple Z-disc proteins. J. Biol. Chem. 2002, 277, 13998–14004. [Google Scholar] [CrossRef]
- Franzini-Armstrong, C.; Protasi, F.; Ramesh, V. Shape, size, and distribution of Ca (2+) release units and couplons in skeletal and cardiac muscles. Biophys. J. 1999, 77, 1528–1539. [Google Scholar] [CrossRef]
- Hayashi, T.; Arimura, T.; Itoh-Satoh, M.; Ueda, K.; Hohda, S.; Inagaki, N.; Takahashi, M.; Hori, H.; Yasunami, M.; Nishi, H.; et al. Tcap gene mutations in hypertrophic cardiomyopathy and dilated cardiomyopathy. J. Am. Coll. Cardiol. 2004, 44, 2192–2201. [Google Scholar] [CrossRef]


{kind=link}
| Ensemble Accession Number | Gene Symbol | Primer Sequence | Amplicon Length |
|---|---|---|---|
| ENSECAG00000011860 | TCAP | TCAPF:GGCTGAATGGAAGGATCTGA TCAPR:TGGTAGGGCAGCTGGTACTC | 192 |
| ENSECAG00000022051 | GAPDH | GAPDHF:GCCGTAACTTCTGTGCTGTG GAPDHR:AATGAAGGGGTCATTGATGG | 156 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefaniuk-Szmukier, M.; Szmatoła, T.; Łątka, J.; Długosz, B.; Ropka-Molik, K. The Blood and Muscle Expression Pattern of the Equine TCAP Gene during the Race Track Training of Arabian Horses. Animals 2019, 9, 574. https://doi.org/10.3390/ani9080574
Stefaniuk-Szmukier M, Szmatoła T, Łątka J, Długosz B, Ropka-Molik K. The Blood and Muscle Expression Pattern of the Equine TCAP Gene during the Race Track Training of Arabian Horses. Animals. 2019; 9(8):574. https://doi.org/10.3390/ani9080574
Chicago/Turabian StyleStefaniuk-Szmukier, Monika, Tomasz Szmatoła, Joanna Łątka, Bogusława Długosz, and Katarzyna Ropka-Molik. 2019. "The Blood and Muscle Expression Pattern of the Equine TCAP Gene during the Race Track Training of Arabian Horses" Animals 9, no. 8: 574. https://doi.org/10.3390/ani9080574
APA StyleStefaniuk-Szmukier, M., Szmatoła, T., Łątka, J., Długosz, B., & Ropka-Molik, K. (2019). The Blood and Muscle Expression Pattern of the Equine TCAP Gene during the Race Track Training of Arabian Horses. Animals, 9(8), 574. https://doi.org/10.3390/ani9080574