Responsiveness Expressions of Bitter Taste Receptors Against Denatonium Benzoate and Genistein in the Heart, Spleen, Lung, Kidney, and Bursa Fabricius of Chinese Fast Yellow Chicken

 , ,
, , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Birds and Procedures
2.3. Feed Formulation and Mixing Procedure
2.4. Organs Weight Measurements
2.5. Sample Collection and RNA Extraction
2.6. Primer Design and RT-PCR
2.7. Housekeeping Gene for Internal Control (β-actin)
2.8. Determination of mRNA Expression by Real-Time PCR Using the Comparative ΔΔCt Method
2.9. Statistical Analysis
2.9.1. Organ Weight Measurements
2.9.2. Gene Expression
3. Results
3.1. Organ Weight Measurements
3.2. Detection of ggTas2Rs Responsiveness Expressions Against Denatonium Benzoate and Genistein
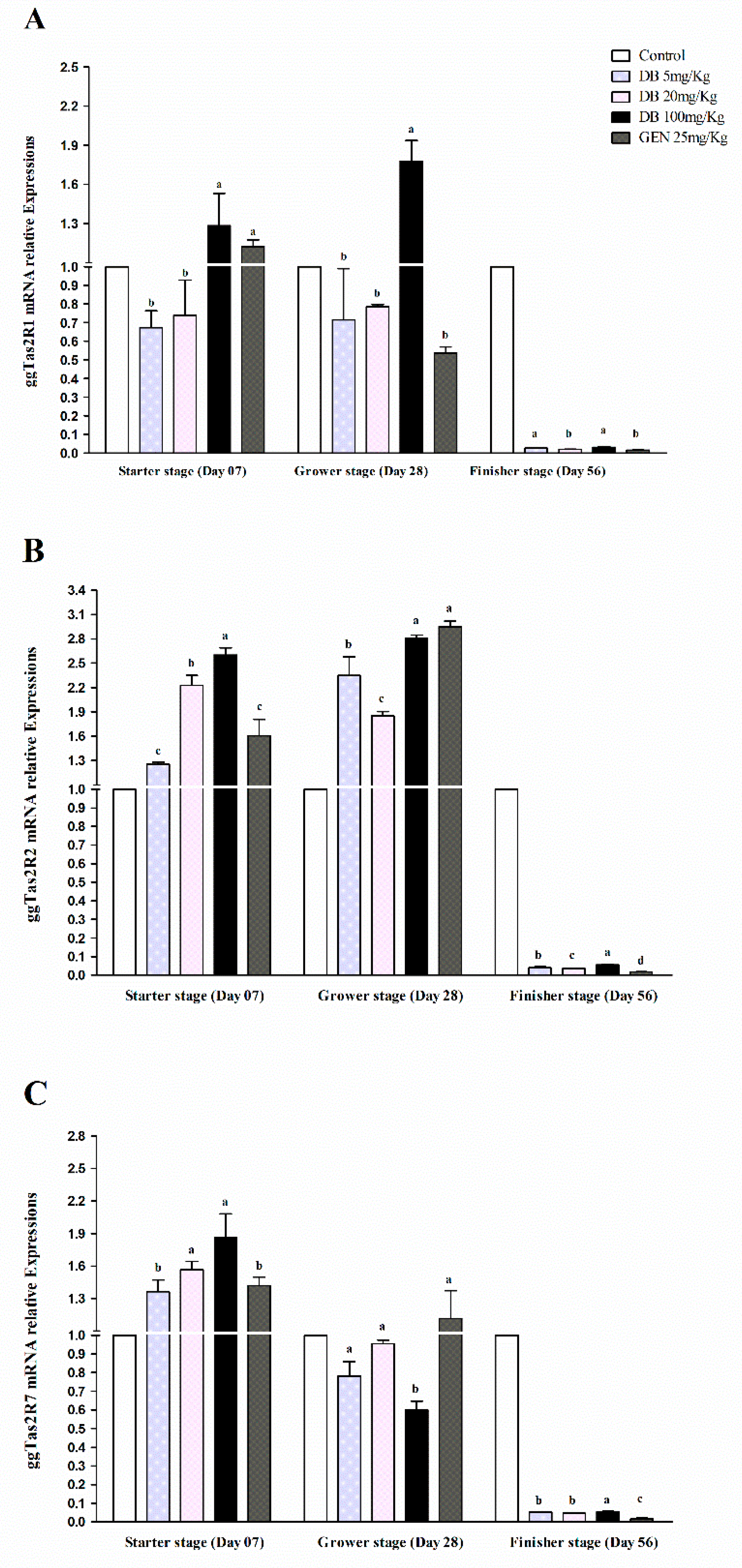
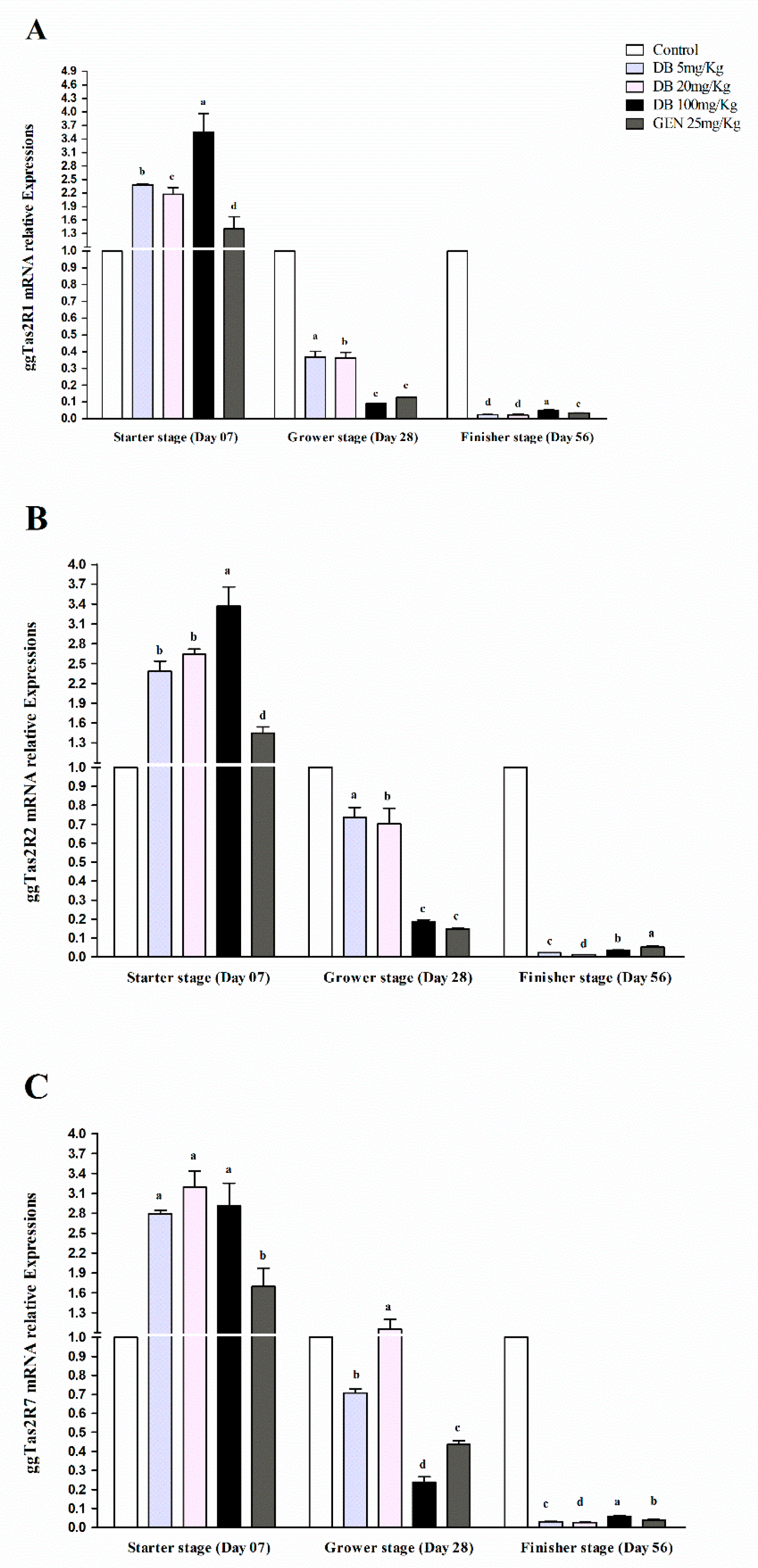
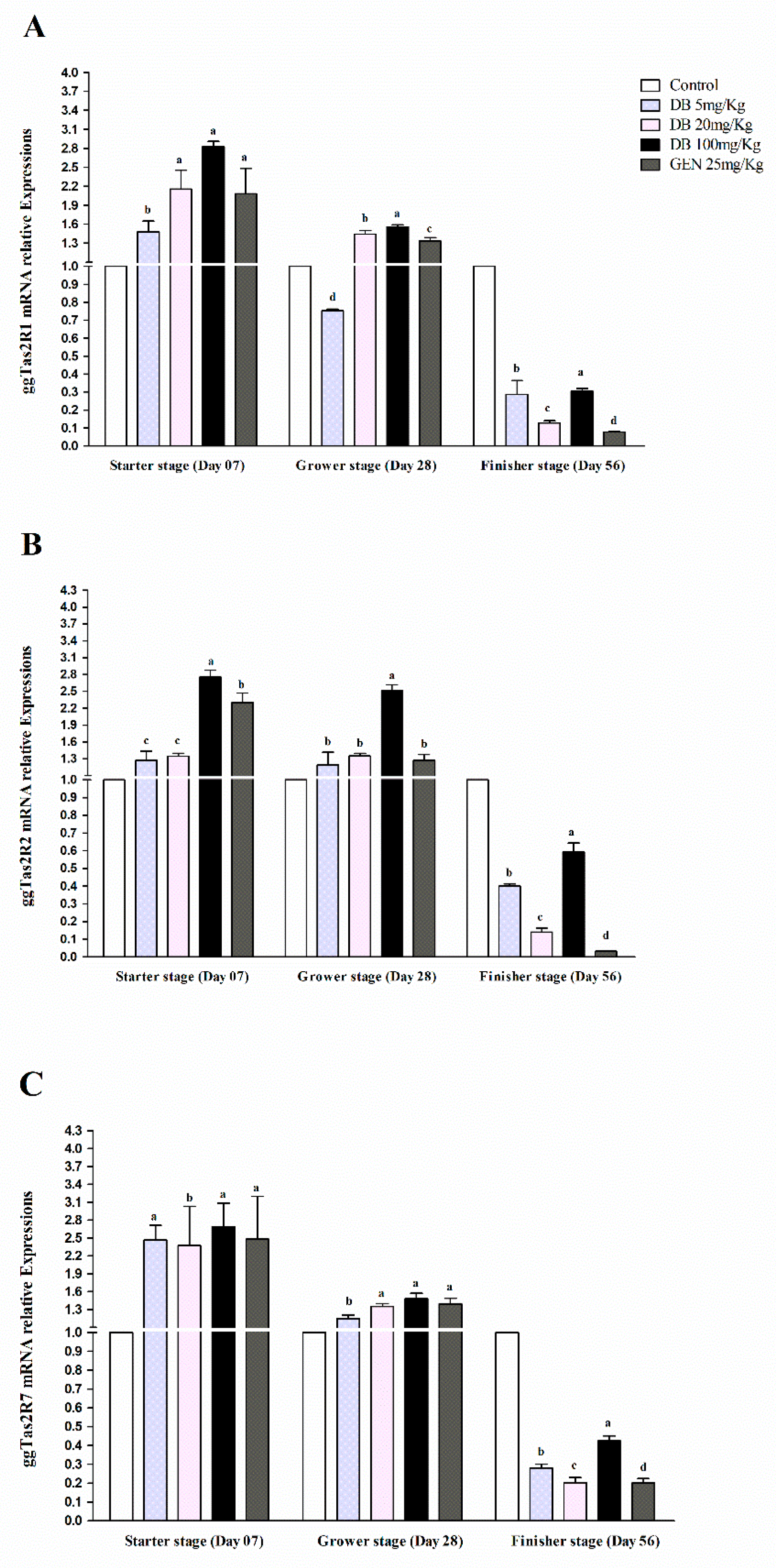
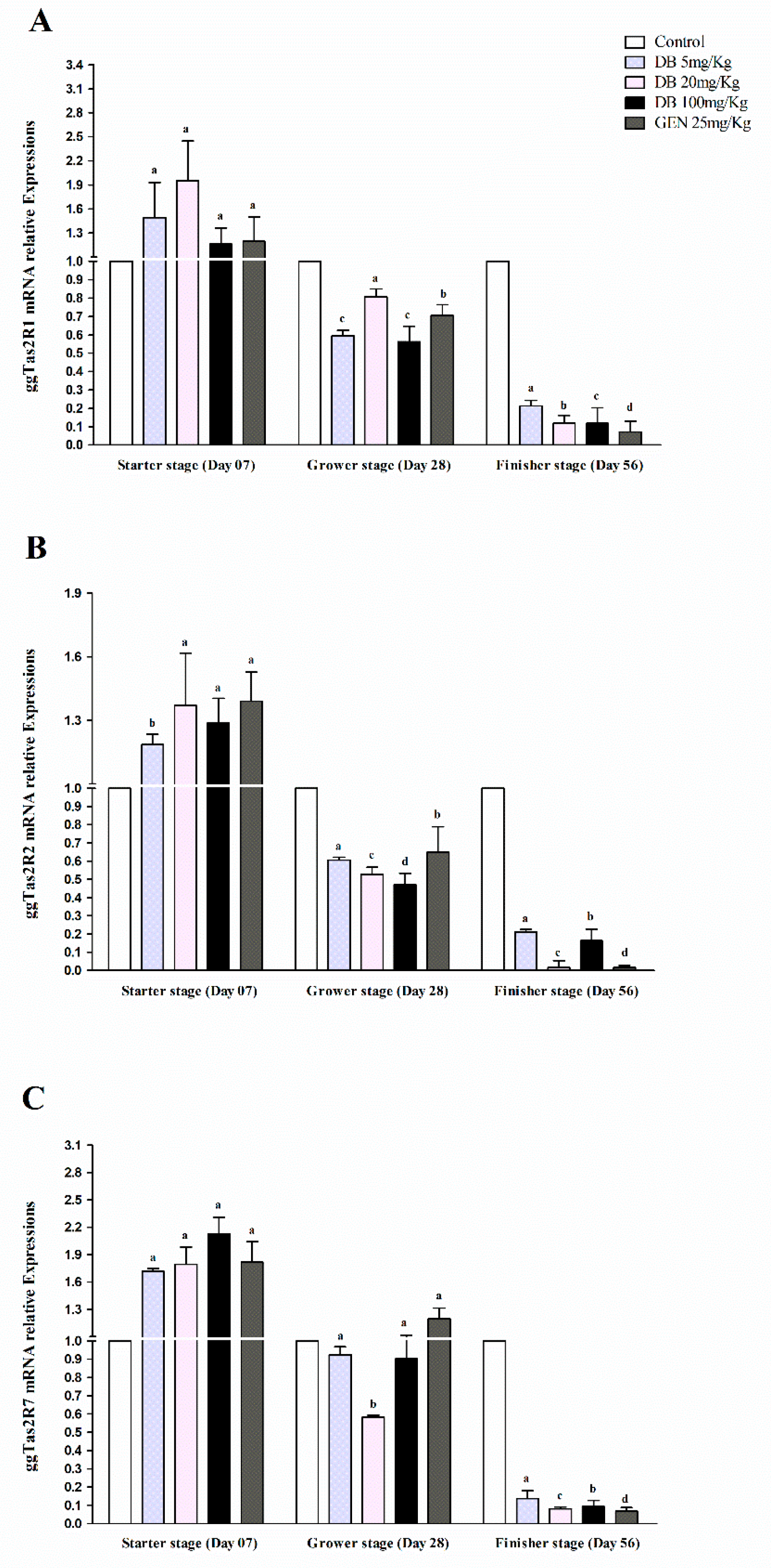
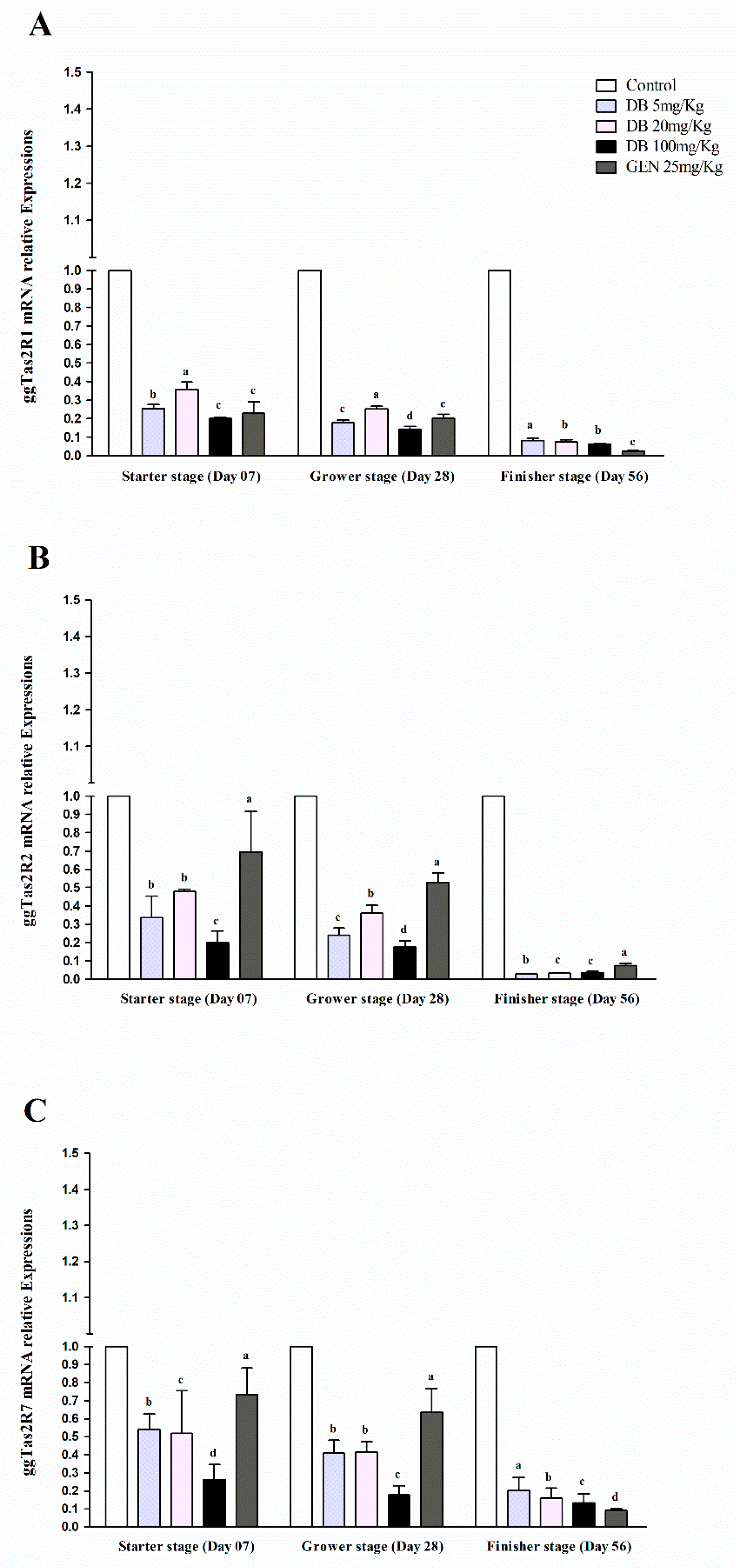
3.2.1. mRNA Responsiveness Expressions of ggTas2Rs in Chicken Heart
3.2.2. mRNA Responsiveness Expression of ggTas2Rs in the Spleen of Chicken
3.2.3. mRNA Responsiveness Expressions of ggTas2Rs in Chicken Lungs
3.2.4. mRNA Responsiveness Expressions of ggTas2Rs in Chicken Kidneys
3.2.5. mRNA Responsiveness Expressions of ggTas2Rs in Chicken Bursa Fabricius
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roura, E.; Baldwin, M.W.; Klasing, K. The avian taste system: Potential implications in poultry nutrition. Anim. Feed Sci. Technol. 2013, 180, 1–9. [Google Scholar] [CrossRef]
- Chandrashekar, J.; Hoon, M.A.; Ryba, N.J.P.; Zuker, C.S. The receptors and cells for mammalian taste. Nature 2006, 444, 288. [Google Scholar] [CrossRef] [PubMed]
- Galindo, M.M.; Voigt, N.; Stein, J.; Lengerich, J.V.; Raguse, J.; Hofmann, T.; Meyerhof, W.; Behrens, M. G- protein-coupled receptors in human fat taste perception. Chem. Senses 2011, 37, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Gentle, M. Taste preference in the chicken (Gallus domesticus L.). Br. Poult. Sci. 1972, 13, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Simitzis, P.E.; Deligeorgis, S.G.; Bizelis, J.A.; Fegeros, K. Feeding preferences in lambs influenced by prenatal flavour exposure. Physiol. Behav. 2008, 93, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Mennella, J.A.; Forestell, C.A.; Morgan, K.L.; Beauchamp, G.K. Early milk feeding influences taste acceptance and liking during infancy. Am. J. Clin. Nutr. 2009, 90, 780S–788S. [Google Scholar] [CrossRef]
- Smeets, P.A.; Charbonnier, L.; Meer, F.V.; Laan, L.N.V.D.; Spetter, M.S. Food-induced brain responses and eating behaviour. Proc. Nutr. Soc. 2012, 71, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Mojet, J.; Heidema, J.; Christ-Hazelhof, E. Taste perception with age: Generic or specific losses in supra-threshold intensities of five taste qualities? Chem. Senses 2003, 28, 397–413. [Google Scholar] [CrossRef]
- Clark, L.; Hagelin, J.; Werner, S. The chemical senses in birds. In Sturkie’s Avian Physiology, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 89–111. [Google Scholar]
- BERKHOUDT, H. Special sense organs: Structure and function of avian taste receptors. Form Func. Birds 1985, 3, 463–496. [Google Scholar]
- Lindemann, B. Taste reception. Physiol. Rev. 1996, 76, 719–766. [Google Scholar] [CrossRef]
- Drewnowski, A.; Gomez-Carneros, C. Bitter taste, phytonutrients, and the consumer: A review. Am. J. Clin. Nutr. 2000, 72, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Senses 2010, 35, 157–170. [Google Scholar] [CrossRef]
- Clark, L. Physiological, ecological, and evolutionary bases for the avoidance of chemical irritants by birds. In Current Ornithology; Springer: Amsterdam, The Netherlands, 1997; pp. 1–37. [Google Scholar]
- Boushy, A.R.E.; Raterink, R. Replacement of soybean meal by cottonseed meal and peanut meal or both in low energy diets for broilers. Poult. Sci. 1989, 68, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Cheled-Shoval, S.; Behrens, B.; Korb, A.; Pizio, A.D.; Meyerhof, W.; Uni, Z.; Niv, M.Y. From cell to beak: In-vitro and in-vivo characterization of chicken bitter taste thresholds. Molecules 2017, 22, 821. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.; Korsching, S.I.; Meyerhof, W. Tuning properties of avian and frog bitter taste receptors dynamically fit gene repertoire sizes. Mol. Biol. Evol. 2014, 31, 3216–3227. [Google Scholar] [CrossRef] [PubMed]
- Rowland, H.M.; Parker, M.R.; Jiang, P.; Reed, D.R.; Beauchamp, G.K. Comparative taste biology with special focus on birds and reptiles. Handb. Olfaction Gustation 2015, 43, 957–982. [Google Scholar]
- Cheled-Shoval, S.L.; Behrens, M.; Meyerhof, W.; Niv, M.Y.; Uni, Z. Perinatal administration of a bitter tastant influences gene expression in chicken palate and duodenum. J. Agric. Food Chem. 2014, 62, 12512–12520. [Google Scholar] [CrossRef]
- Gura, T.; Margolskee, R.F.; Kinnamon, S.C. Taste receptor cell responses to the bitter stimulus denatonium involve Ca2+ influx via store-operated channels. J. Neurophysiol. 2002, 87, 3152–3155. [Google Scholar]
- Payne, H. Bitrex-a bitter solution to safety. Chem. Ind. 1988, 721–723. [Google Scholar]
- Chen, M.C.; Wu, S.V.; Joseph, R.R.J.; Rozengurt, E. Bitter stimuli induce Ca2+ signaling and CCK release in enteroendocrine STC-1 cells: Role of L-type voltage-sensitive Ca2+ channels. Am. J. Physiol. Cell Physiol. 2006, 291, C726–C739. [Google Scholar] [CrossRef]
- Kim, K.S.; Egan, J.M.; Jang, H.J. Denatonium induces secretion of glucagon-like peptide-1 through activation of bitter taste receptor pathways. Diabetology 2014, 57, 2117–2125. [Google Scholar] [CrossRef] [PubMed]
- Glendinning, J.I.; Yiin, Y.M.; Ackroff, K.; Sclafani, A. Intragastric infusion of denatonium conditions flavor aversions and delays gastric emptying in rodents. Physiol. Behav. 2008, 93, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Schier, L.A.; Davidson, T.L.; Powley, T.L. Ongoing ingestive behavior is rapidly suppressed by a preabsorptive, intestinal “bitter taste” cue. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1557–R1568. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Lee, S.H. Genistein, a soy isoflavone, is a potent α-glucosidase inhibitor. FEBS Lett. 2001, 501, 84–86. [Google Scholar] [CrossRef]
- Azadbakht, L.; Shakerhosseini, R.; Atabak, S.; Jamshidian, M.; Mehrabi, Y.; Esmaill-Zadeh, A. Beneficiary effect of dietary soy protein on lowering plasma levels of lipid and improving kidney function in type II diabetes with nephropathy. Eur. J. Clin. Nutr. 2003, 57, 1292. [Google Scholar] [CrossRef] [PubMed]
- Vedavanam, K.; Srijayanta, S.; O’Reilly, J.; Raman, A.; Wiseman, H. Antioxidant action and potential antidiabetic properties of an isoflavonoid-containing soyabean phytochemical extract (SPE). Pharmacol. Pharm. 1999, 13, 601–608. [Google Scholar] [CrossRef]
- Retana-Márquez, S.; Juárez-Rojas, L.; Hernández, A.; Romero, C.; López, G.; Miranda, L.; Guerrero-Aguilera, A.; Solano, F.; Hernández, E.; Chemineau, P.; et al. Comparison of the effects of mesquite pod and Leucaena extracts with phytoestrogens on the reproductive physiology and sexual behavior in the male rat. Physiol. Behav. 2016, 164, 1–10. [Google Scholar]
- Tizzano, M.; Finger, T.E. Chemosensors in the nose: Guardians of the airways. Physiology 2013, 28, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.; Al-Rammahi, M.; Moran, A.; Marcello, M.; Ninomiya, Y.; Shirazi-Beechey, S.P. Sensing of amino acids by the gut-expressed taste receptor T1R1-T1R3 stimulates CCK secretion. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 304, G271–G282. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.R.; Porrello, E.R.; Purdue, B.; Chan, H.W.; Voigt, A.; Frenzel, S.; Hannan, R.D.; Moritz, K.M.; Simmons, D.G.; Molenaar, P.; et al. Expression, regulation and putative nutrient-sensing function of taste GPCRs in the heart. PLoS ONE 2013, 8, e64579. [Google Scholar] [CrossRef]
- Xu, J.; Cao, J.; Iguchi, N.; Riethmacher, D.; Huang, L. Functional characterization of bitter-taste receptors expressed in mammalian testis. Mol. Hum. Reprod. 2012, 19, 17–28. [Google Scholar] [CrossRef]
- Lee, R.J.; Xiong, G.X.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.H.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef]
- Clark, A.A.; Liggett, S.B.; Munger, S.D. Extraoral bitter taste receptors as mediators of off-target drug effects. FASEB J. 2012, 26, 4827–4831. [Google Scholar] [CrossRef]
- Lu, P.; Zhang, C.H.; Lifshitz, L.M.; ZhuGe, R. Extraoral bitter taste receptors in health and disease. J. Gen. Physiol. 2017, 149, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Gentle, M.J. Extra-lingual chemoreceptors in the chicken (Gallus domesticus). Chem. Senses 1978, 3, 325–329. [Google Scholar] [CrossRef]
- Gentle, M.J. Oral behaviour following forebrain ablations in Gallus domesticus. Behav. Processes 1981, 6, 109–120. [Google Scholar] [CrossRef]
- Gentle, M.J.; Harkin, C. The effect of sweet stimuli on oral behaviour in the chicken. Chem. Senses 1979, 4, 183–190. [Google Scholar] [CrossRef]
- Gentle, M.; Dewar, W. The effects of vitamin A deficiency on oral gustatory behaviour in chicks. Br. Poult. Sci. 1981, 22, 275–279. [Google Scholar] [CrossRef]
- Ganchrow, D.; Ganchrow, J.R. Number and distribution of taste buds in the oral cavity of hatchling chicks. Physiol. Behav. 1985, 34, 889–894. [Google Scholar] [CrossRef]
- Behrens, M.; Meyerhof, W. Gustatory and extragustatory functions of mammalian taste receptors. Physiol. Behav. 2011, 105, 4–13. [Google Scholar] [CrossRef]
- Behrens, M.; Meyerhof, W. Bitter taste receptor research comes of age: From characterization to modulation of TAS2Rs. Semin. Cell Dev. Biol. 2013, 24, 215–221. [Google Scholar] [CrossRef]
- Kinnamon, S.C. Taste receptor signalling-from tongues to lungs. Acta. Physiol. 2012, 204, 158–168. [Google Scholar] [CrossRef]
- Shaik, F.A.; Singh, N.; Arakawa, M.; Duan, K.; Bhullar, R.P.; Chelikani, P. Bitter taste receptors: Extraoral roles in pathophysiology. Int. J. Biochem. Cell Biol. 2016, 77, 197–204. [Google Scholar] [CrossRef]
- Iwata, S.; Yoshida, R.; Ninomiya, Y. Taste transductions in taste receptor cells: Basic tastes and moreover. Curr. Pharm. Des. 2014, 20, 2684–2692. [Google Scholar] [CrossRef]
- Naline, E.; Philippe, N.; Guy, D.; Lucien, D.; Hayssam, T.; Domenico Regoli, B.; Advenier, C. Characterization of neurokinin effects and receptor selectivity in human isolated bronchi. Am. Rev. Respir. Dis. 1989, 140, 679–686. [Google Scholar] [CrossRef]
- Go, Y. Lineage-specific expansions and contractions of the bitter taste receptor gene repertoire in vertebrates. Mol. Biol. Evol. 2006, 23, 964–972. [Google Scholar] [CrossRef]
- Cheled-Shoval, S.L.; Druyan, S.; Uni, Z. Bitter, sweet and umami taste receptors and downstream signaling effectors: Expression in embryonic and growing chicken gastrointestinal tract. Poult. Sci. 2015, 94, 1928–1941. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Cheled-Shoval, S.L.; Reicher, N.; Niv, M.Y.; Uni, Z. Detecting thresholds for bitter, umami, and sweet tastants in broiler chicken using a 2-choice test method. Poult. Sci. 2017, 96, 2206–2218. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Gan, F.; Chen, X.X.; Liao, S.F.; Lv, C.H.; Ren, F.; Ye, G.P.; Pan, C.L.; Huang, D.; Shi, J.; Shi, X.L.; et al. Selenium-enriched probiotics improve antioxidant status, immune function, and selenoprotein gene expression of piglets raised under high ambient temperature. J. Agric. Food Chem. 2014, 62, 4502–4508. [Google Scholar] [CrossRef]
- Druyan, S.; Levi, E.; Shinder, D.; Stern, T. Reduced O2 concentration during CAM development-Its effect on physiological parameters of broiler embryos. Poult. Sci. 2012, 91, 987–997. [Google Scholar] [CrossRef]
- Behrens, M.; Prandi, S.; Meyerhof, W. Taste receptor gene expression outside the gustatory system. In Taste and Smell; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–34. [Google Scholar]
- Lee, R.J.; Cohen, N.A. Bitter and sweet taste receptors in the respiratory epithelium in health and disease. J. Mol. Med. 2014, 92, 1235–1244. [Google Scholar] [CrossRef]
- Di Pizio, A.; Kruetzfeldt, L.M.; Cheled-Shoval, S.; Meyerhof, W.; Behrens, M.; Niv, M.Y. Ligand binding modes from low resolution GPCR models and mutagenesis: Chicken bitter taste receptor as a test-case. Sci. Rep. 2017, 7, 8223. [Google Scholar] [CrossRef]
- Martin, B.; Dotson, C.D.; Shin, Y.K.; Ji, S.; Drucker, D.J.; Maudsley, S.; Munger, S.D. Modulation of taste sensitivity by GLP-1 signaling in taste buds. Ann. N. Y. Acad. Sci. 2009, 1170, 98–101. [Google Scholar] [CrossRef]
- Jang, H.J.; Kokrashvili, Z.; Theodorakis, M.J.; Carlson, O.D.; Kim, B.J.; Zhou, J.; Kim, H.H.; Xu, X.; Chan, S.L.; Juhaszova, M.; et al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc. Natl. Acad. Sci. USA 2007, 104, 15069–15074. [Google Scholar] [CrossRef]
- Dehkordi, O.; Rose, J.E.; Fatemi, M.; Allard, J.S.; Balan, K.V.; Young, J.K.; Fatima, S.; Millis, R.M.; Jayam-Trouth, A. Neuronal expression of bitter taste receptors and downstream signaling molecules in the rat brainstem. Brain Res. 2012, 1475, 1–10. [Google Scholar] [CrossRef]
- Dey, B.; Kawabata, F.; Kawabata, Y.; Nishimura, S.; Tabata, S. Bitter Taste Sensitivity and the Expression of Bitter Taste Receptors at Different Growth Stages of Chicks. J. Poult. Sci. 2018, 55, 0170188. [Google Scholar] [CrossRef]
- Dey, B.; Kawabata, F.; Kawabata, Y.; Yoshida, Y.; Nishimura, S.; Tabata, S. Identification of functional bitter taste receptors and their antagonist in chickens. Biochem. Biophys. Res. Commun. 2017, 482, 693–699. [Google Scholar] [CrossRef]
- KUDO, K.; SHIRAISHI, J.-I.; NISHIMURA, S.; BUNGO, T.; TABATA, S. The number of taste buds is related to bitter taste sensitivity in layer and broiler chickens. Anim. Sci. J. 2010, 81, 240–244. [Google Scholar] [CrossRef]
- Hyde, R.J.; Feller, R.P. Age and sex effects on taste of sucrose, NaCl, citric acid and caffeine. Neurobiol. Aging 1981, 2, 315–318. [Google Scholar] [CrossRef]
- Roland, W.S.; . Vincken, J.-P.; Buren, L.V.; Gruppen, H.; Smit, G. Soy isoflavones and other isoflavonoids activate the human bitter taste receptors hTAS2R14 and hTAS2R39. J. Agric. Food Chem. 2011, 59, 11764–11771. [Google Scholar] [CrossRef]
- Roland, W.S.; Buren, L.V.; Gruppen, H.; Driesse, M.; Gouka, R.J.; Smit, G.; Vincken, J.-P. Bitter taste receptor activation by flavonoids and isoflavonoids: Modeled structural requirements for activation of hTAS2R14 and hTAS2R39. J.Agric. Food Chem. 2013, 61, 10454–10466. [Google Scholar] [CrossRef]
- Roland, W.S.; Gouka, R.J.; Gruppen, H.; Driesse, M.; Buren, L.V.; Vincken, J.-P. 6-Methoxyflavanones as bitter taste receptor blockers for hTAS2R39. PLoS ONE 2014, 9, e94451. [Google Scholar] [CrossRef]
- Rasouli, E.; Jahanian, R. Improved performance and immunological responses as the result of dietary genistein supplementation of broiler chicks. Animal 2015, 9, 1473–1480. [Google Scholar] [CrossRef]
- Palanisamy, N.; Viswanathan, P.; Anuradha, C.V. Effect of genistein, a soy isof lavone, on whole body insulin sensitivity and renal damage induced by a high-fructose diet. Ren. Fail. 2008, 30, 645–654. [Google Scholar] [CrossRef]
- Behrens, M.; Meyerhof, W. Oral and extraoral bitter taste receptors. In Sensory and Metabolic Control of Energy Balance; Springer: Berlin/Heidelberg, Germany, 2011; Volume 52, pp. 87–99. [Google Scholar]
- Prandi, S.; Bromke, M.; Hübner, S.; Voigt, A.; Boehm, U.; Meyerhof, W.; Behrens, M. A subset of mouse colonic goblet cells expresses the bitter taste receptor Tas2r131. PLoS ONE 2013, 8, e82820. [Google Scholar] [CrossRef]
- Garcia-Esparcia, P.; Agatha Schlüter, M.; Carmona, M.; Moreno, J.; Ansoleaga, B.; Torrejón-Escribano, B.; Gustincich, S.; Pujol, A.; Ferrer, I. Functional genomics reveals dysregulation of cortical olfactory receptors in Parkinson disease: Novel putative chemoreceptors in the human brain. J. Neuropathol. Exp. Neurol. 2013, 72, 524–539. [Google Scholar] [CrossRef]
- Meyer-Gerspach, A.; Wölnerhanssen, B.; Beglinger, C. Gut sweet taste receptors and their role in metabolism. In How Gut and Brain Control Metabolism; Karger Publishers: Berlin, Germany, 2014; pp. 123–133. [Google Scholar]
- Shah, A.S.; Ben-Shahar, Y.; Moninger, T.O.; Kline, K.J.; Welsh, M.J. Motile cilia of human airway epithelia are chemosensory. Science 2009, 325, 1131–1134. [Google Scholar] [CrossRef]
- Tizzano, M.; Cristofoletti, M.; Sbarbati, A.; Finger, T.E. Expression of taste receptors in solitary chemosensory cells of rodent airways. BMC Pulm. Med. 2011, 11, 3. [Google Scholar] [CrossRef]
- Behrens, M.; Brockhoff, A.; Kuhn, C.; Bufe, B.; Winnig, M.; Meyerhof, W. The human taste receptor hTAS2R14 responds to a variety of different bitter compounds. Biochem. Biophys. Res. Commun. 2004, 319, 479–485. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Diet | |
|---|---|---|
| D 1 to 28 | D 28 to 56 | |
| Ingredient (%) | ||
| Corn | 61.52 | 74.00 |
| Soybean meal | 29.00 | 12.00 |
| Soybean oil | 2.44 | 2.60 |
| Corn gluten meal | 2.00 | 7.32 |
| Dicalcium phosphate | 1.68 | 1.02 |
| Premix | 1.50 | 1.00 |
| Limestone | 1.15 | 1.05 |
| Lysine sulfate | 0.51 | 0.80 |
| Methionine | 0.20 | 0.21 |
| Total | 100 | 100 |
| Calculation of nutrients | ||
| Metabolizable energy, MJ/kg | 11.92 | 12.13 |
| Crude protein, % | 21.00 | 19.00 |
| Lysine, % | 1.10 | 0.97 |
| Methionine, % | 0.46 | 0.40 |
| Methionine + cystine, % | 0.80 | 0.72 |
| Calcium, % | 1.00 | 0.90 |
| Available phosphorus, % | 0.70 | 0.65 |
| Target | Accession No | Forward Primer | Reverse Primer | Product Size (bp) |
|---|---|---|---|---|
| ggTas2R1 | AB249766.1 | GGTGCCATCAAGACAGTCTTCTC | ACAGGCAGCCACTACAACAACA | 135 |
| ggTas2R2 | AB249767.1 | GCGATGATTCCATGGCTGC | CGTTGACCTGCAGAGGTAGG | 102 |
| ggTas2R7 | NM_001080719.1 | TGGCAGAGCAGCACAACACAAC | TACAAGACGCAGCCACAATGAA | 111 |
| β-actin | NM_205518.1 | CCAGCCATGTATGTAGCCATCCAG | ACGGCCAGCCAGATCCAGAC | 162 |
| Stages | Treatments | Heart (g) | Spleen (g) | Lungs (g) | Kidneys (g) | Bursa Fabricius (g) | Live Body Weight (g) |
|---|---|---|---|---|---|---|---|
| Starter | Control | 0.64 ± 0.04b | 0.32 ± 0.05a | 0.52 ± 0.01b,c | 0.72 ± 0.17a | 0.21 ± 0.30a | 70.94 ± 0.79b |
| DB-Low Dose | 0.66 ± 0.02b | 0.25 ± 0.03b | 0.52 ± 0.02b,c | 0.70 ± 0.10b | 0.09 ± 0.03b | 76.34 ± 0.41a | |
| DB-Medium Dose | 0.51 ± 0.04c | 0.22 ± 0.01c | 0.63 ± 0.04a | 0.68 ± 0.08b,c | 0.09 ± 0.04b | 63.64 ± 0.88c | |
| DB-High Dose | 0.43 ± 0.02d | 0.19 ± 0.03c,d | 0.47 ± 0.09d | 0.65 ± 0.08c | 0.09 ± 0.02b | 60.02 ± 0.98d | |
| GEN | 0.69 ± 0.07a | 0.34 ± 0.04a | 0.57 ± 0.01b | 0.73 ± 0.05a | 0.09 ± 0.01b | 63.70 ± 0.99c | |
| Grower | Control | 2.09 ± 0.28b,c | 0.58 ± 0.24a | 1.75 ± 0.17b | 2.46 ± 0.08a,b | 0.55 ± 0.02b | 255.00 ± 1.33a |
| DB-Low Dose | 2.13 ± 0.13b | 0.56 ± 0.15b | 1.31 ± 0.13c | 2.14 ± 0.09b | 0.57 ± 0.03a | 220.50 ± 2.14b | |
| DB-Medium Dose | 1.71 ± 0.24c | 0.37 ± 0.13c | 1.84 ± 0.23a | 2.13 ± 0.04b | 0.47 ± 0.02c | 197.10 ± 3.12d | |
| DB-High Dose | 1.63 ± 0.27d | 0.31 ± 0.14d | 1.29 ± 0.07d | 2.11 ± 0.06c,d | 0.48 ± 0.02c | 197.40 ± 2.13d | |
| GEN | 2.37 ± 0.19a | 0.45 ± 0.19b,c | 1.79 ± 0.11b | 2.56 ± 0.04a | 0.41 ± 0.02d | 208.30 ± 1.91c | |
| Finisher | Control | 6.41 ± 0.02b | 2.24 ± 0.17b | 4.75 ± 0.11b,c | 8.58 ± 0.03a | 2.26 ± 0.09b,c | 888.20 ± 2.12a |
| DB-Low Dose | 6.92 ± 0.02a | 2.36 ± 0.25a | 3.97 ± 1.13c | 7.87 ± 0.02b | 2.60 ± 0.05a | 687.00 ± 2.13c | |
| DB-Medium Dose | 5.29 ± 0.03c | 1.79 ± 0.19c | 4.86 ± 1.31a | 7.77 ± 0.06c | 2.18 ± 0.08d,e | 725.30 ± 1.93b | |
| DB-High Dose | 5.27 ± 0.08c,d | 1.59 ± 0.32d | 3.66 ± 0.16d | 7.22 ± 0.02d | 2.21 ± 0.03c | 686.50 ± 1.75d | |
| GEN | 6.68 ± 0.03a,b | 2.32 ± 0.16a | 4.77 ± 0.50a,b | 8.62 ± 0.04a | 2.20 ± 0.02c | 620.50 ± 2.12e | |
| P-Value | Starter | <0.001 | <0.001 | <0.001 | <0.001 | 0.004 | 0.0047 |
| Grower | 0.005 | 0.004 | 0.005 | 0.004 | 0.005 | 0.0038 | |
| Finisher | 0.001 | 0.005 | 0.001 | 0.003 | 0.004 | 0.0044 | |
| Feed (Treatments) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Feed × Stages | 0.183 | 0.181 | 0.111 | 0.094 | 0.917 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdard, E.; Lv, Z.; Jiang, J.; Wei, Q.; Shi, Z.; Malyar, R.M.; Yu, D.; Shi, F. Responsiveness Expressions of Bitter Taste Receptors Against Denatonium Benzoate and Genistein in the Heart, Spleen, Lung, Kidney, and Bursa Fabricius of Chinese Fast Yellow Chicken. Animals 2019, 9, 532. https://doi.org/10.3390/ani9080532
Hamdard E, Lv Z, Jiang J, Wei Q, Shi Z, Malyar RM, Yu D, Shi F. Responsiveness Expressions of Bitter Taste Receptors Against Denatonium Benzoate and Genistein in the Heart, Spleen, Lung, Kidney, and Bursa Fabricius of Chinese Fast Yellow Chicken. Animals. 2019; 9(8):532. https://doi.org/10.3390/ani9080532
Chicago/Turabian StyleHamdard, Enayatullah, Zengpeng Lv, Jingle Jiang, Quanwei Wei, Zhicheng Shi, Rahmani Mohammad Malyar, Debing Yu, and Fangxiong Shi. 2019. "Responsiveness Expressions of Bitter Taste Receptors Against Denatonium Benzoate and Genistein in the Heart, Spleen, Lung, Kidney, and Bursa Fabricius of Chinese Fast Yellow Chicken" Animals 9, no. 8: 532. https://doi.org/10.3390/ani9080532
APA StyleHamdard, E., Lv, Z., Jiang, J., Wei, Q., Shi, Z., Malyar, R. M., Yu, D., & Shi, F. (2019). Responsiveness Expressions of Bitter Taste Receptors Against Denatonium Benzoate and Genistein in the Heart, Spleen, Lung, Kidney, and Bursa Fabricius of Chinese Fast Yellow Chicken. Animals, 9(8), 532. https://doi.org/10.3390/ani9080532