Author Contributions
S.R. contributed to the methodology, validation, formal analysis, investigation, data curation, and original draft manuscript preparation. A.S. contributed to the methodology, investigation, review and editing, and visualization. A.J. contributed to the conceptualization, methodology, formal analysis, investigation, resources, reviewing and editing, project administration, and funding acquisition.
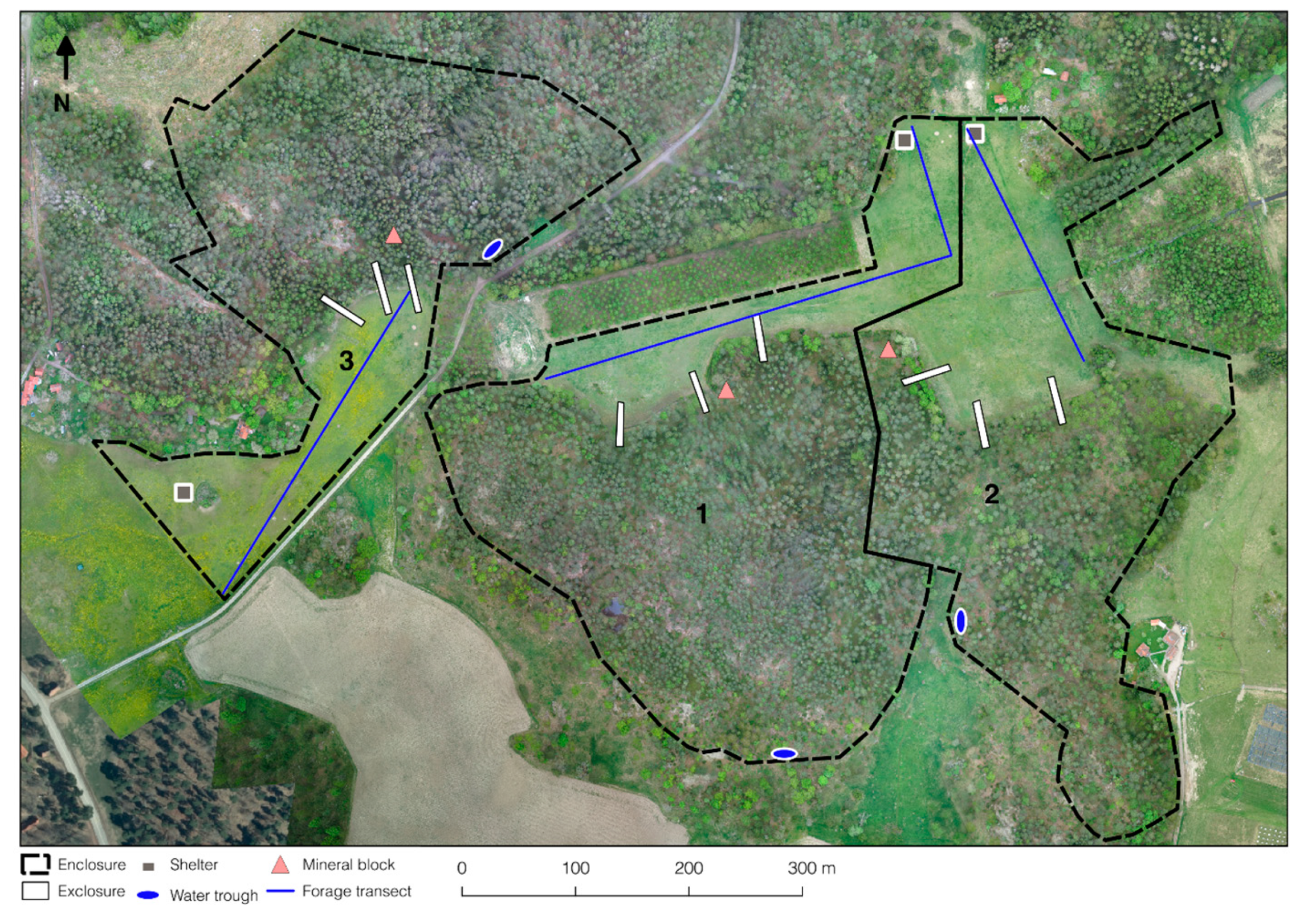
Figure 1.
Aerial view of the three enclosures (1, 2, and 3) used in the study, showing position of shelters, water troughs, mineral blocks, exclosures, and pasture transects. Photo taken on 24 May 2016 at 150 m altitude.
Figure 1.
Aerial view of the three enclosures (1, 2, and 3) used in the study, showing position of shelters, water troughs, mineral blocks, exclosures, and pasture transects. Photo taken on 24 May 2016 at 150 m altitude.
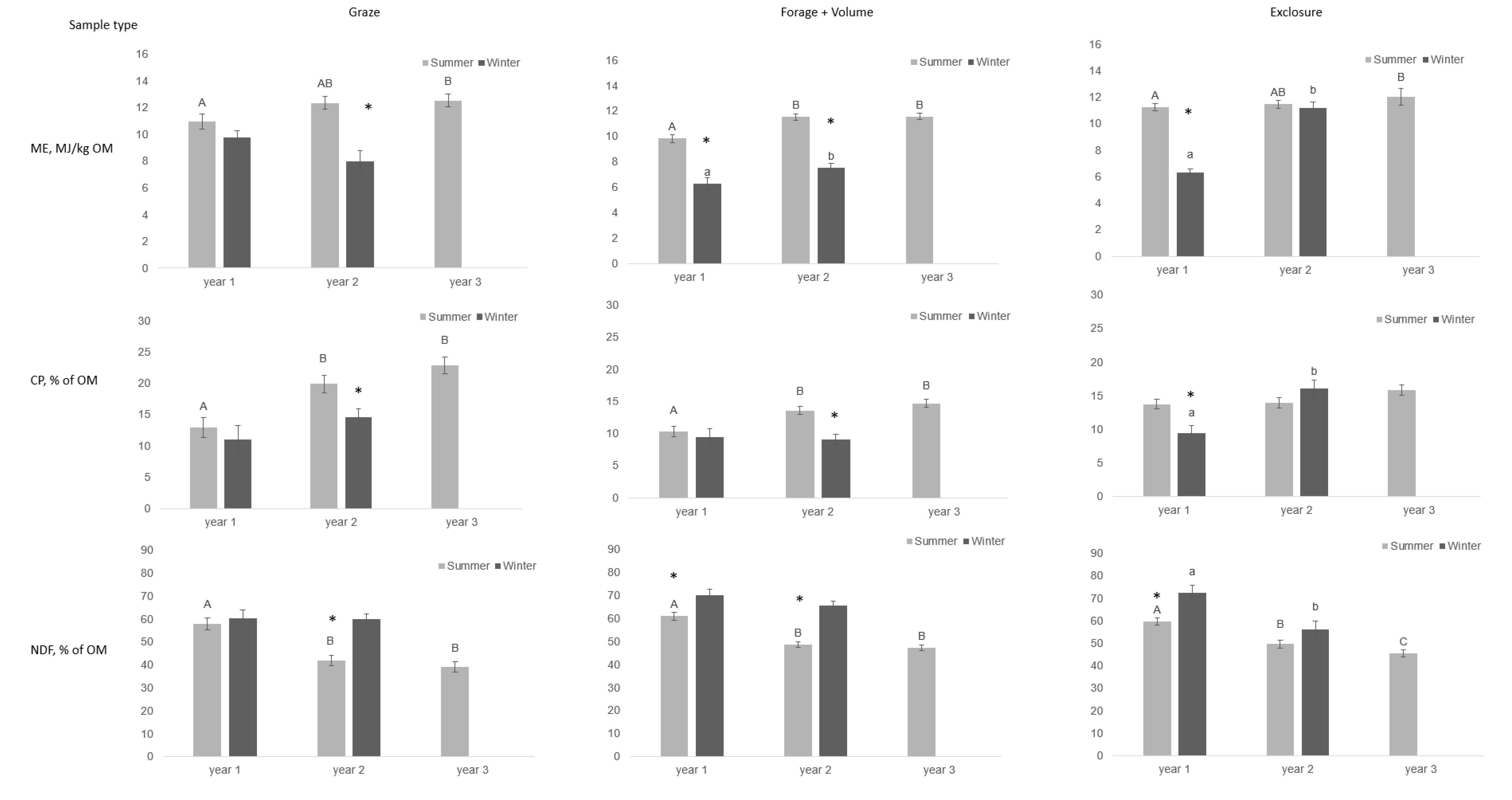
Figure 2.
Content of metabolizable energy (ME) per kg organic matter (OM), crude protein (CP) as % of OM, and neutral detergent fiber (NDF) as % of OM in three types of pasture samples (graze, forage + volume, exclosure) collected monthly in three enclosures grazed by 12 Gotlandsruss between May 2014 and September 2016. One year is defined as start of summer season to end of winter season. Each enclosure contained three exclosures. An asterisk indicates significant difference between seasons within year (p < 0.05). Different lowercase letters (a, b) indicate differences between years within winter season, while different uppercase letters (A, B) indicate differences between years within summer season (p < 0.05).
Figure 2.
Content of metabolizable energy (ME) per kg organic matter (OM), crude protein (CP) as % of OM, and neutral detergent fiber (NDF) as % of OM in three types of pasture samples (graze, forage + volume, exclosure) collected monthly in three enclosures grazed by 12 Gotlandsruss between May 2014 and September 2016. One year is defined as start of summer season to end of winter season. Each enclosure contained three exclosures. An asterisk indicates significant difference between seasons within year (p < 0.05). Different lowercase letters (a, b) indicate differences between years within winter season, while different uppercase letters (A, B) indicate differences between years within summer season (p < 0.05).
Figure 3.
(A) Monthly variation and (B) seasonal and between-year variation in crude protein (CP) content (LSmeans ± SE) as % of organic matter (OM) in feces from Gotlandsruss grazing without supplementary feeding from May 2014 to September 2016. One year is defined as start of summer season to end of winter season.
Figure 3.
(A) Monthly variation and (B) seasonal and between-year variation in crude protein (CP) content (LSmeans ± SE) as % of organic matter (OM) in feces from Gotlandsruss grazing without supplementary feeding from May 2014 to September 2016. One year is defined as start of summer season to end of winter season.
Table 1.
Area (ha) of vegetation types within each enclosure and dominant plant species identified in a vegetation inventory performed in all three enclosures in May 2014 (study start).
Table 1.
Area (ha) of vegetation types within each enclosure and dominant plant species identified in a vegetation inventory performed in all three enclosures in May 2014 (study start).
| Vegetation Type, ha | Enclosure 1 | Enclosure 2 | Enclosure 3 |
|---|
| Grassland | 2.7 | 3.3 | 2.7 |
| Forest | 10.7 | 5.8 | 6.8 |
| Semi-forest a | 0 | 1.3 | 0.2 |
| Total area | 13.4 | 10.3 | 9.7 |
| Dominant Plant Species in Grassland, % | | | |
| Grasses | 60 | 57 | 26 |
| Yarrow (Achillea millefolium) | 10 | 14 | |
| Dandelion (Taraxacum spp.) | 7 | 10 | 47 |
| Hempnettle (Galeopsis tetrahit) | | 6 | |
| White clover (Trifolium repens) | | | 5 |
| Dominant Ground Cover in Forest, % | | | |
| Mosses | 46 | 29 | 55 |
| Grasses | 11 | 26 | 12 |
| Bilberry (Vaccinium myrtillus) | 12 | 7 | 10 |
| Lingonberry (Vaccinium vitis-idaea) | 5 | 4 | 3 |
Table 2.
Content of dry matter (DM), metabolizable energy (ME) per kg organic matter (OM), crude protein (CP) as % of OM, and neutral detergent fiber (NDF) as % of OM in three different types of pasture samples collected in three enclosures grazed by horses, and in three exclosures per enclosure, monthly between May 2014 and September 2016, except for December 2014–March 2015 and January-April 2016.
Table 2.
Content of dry matter (DM), metabolizable energy (ME) per kg organic matter (OM), crude protein (CP) as % of OM, and neutral detergent fiber (NDF) as % of OM in three different types of pasture samples collected in three enclosures grazed by horses, and in three exclosures per enclosure, monthly between May 2014 and September 2016, except for December 2014–March 2015 and January-April 2016.
| Sample | Enclosures | Exclosures | p |
|---|
| Forage | Volume | Graze |
|---|
| ME, MJ/kg OM | 10.0 ± 0.2 a | 10.0 ± 0.2 a | 10.9 ± 0.2 b | 10.5 ± 0.2 b | 0.0002 |
| CP, % of OM | 11.9 ± 0.4 a | 11.6 ± 0.5 a | 17.0 ± 0.4 b | 13.7 ± 0.4 c | <0.0001 |
| NDF, % of OM | 57.2 ± 0.7 a | 56.7 ± 0.9 a | 51.0 ± 0.7 b | 56.8 ± 0.8 a | <0.0001 |
| DM, % | 34 ± 1 a | 34 ± 1 a | 29 ± 1 b | 27 ± 1 b | <0.0001 |
Table 3.
Mean grass sward height and content of dry matter (DM) and metabolizable energy (ME) in the enclosures grazed by horses and in the exclosures within each enclosure mown monthly from May 2014 to September 2016. Exclosures were mown at the same spots as sward height measurements were made.
Table 3.
Mean grass sward height and content of dry matter (DM) and metabolizable energy (ME) in the enclosures grazed by horses and in the exclosures within each enclosure mown monthly from May 2014 to September 2016. Exclosures were mown at the same spots as sward height measurements were made.
| | Enclosures | Exclosures | p Means |
|---|
| En1 | En2 | En3 | Mean | En1 | En2 | En3 | Mean |
|---|
| Grass sward height, cm | 5.5 ± 0.1 a | 7.7 ± 0.2 b | 5.5 ± 0.2 a | 5.3 ± 0.1 | 4.3 ± 0.4 a | 4.9 ± 0.4 a,b | 6.0 ± 0.4 b | 5.6 ± 0.4 | <0.05 |
| DM, kg/ha | 957 ± 114 a | 1393 ± 114 b | 741 ± 114 a | 886 ± 73 | 525 ± 111 | 487 ± 111 | 721 ± 111 | 326 ± 76 | <0.0001 |
| ME, MJ/ha | 8494 ± 895 a | 11,204 ± 928 b | 7351 ± 895 a | 7470 ± 636 | 5208 ± 1171 | 4912 ± 1171 | 7760 ± 1171 | 3048 ± 701 | <0.0001 |
Table 4.
Mean summer season grass sward height and content of dry matter (DM) and metabolizable energy (ME) per year in the enclosures grazed by horses and in the exclosures within each enclosure mown monthly from May 2014 to September 2016. Exclosures were mown at the same spots as sward height measurements were made.
Table 4.
Mean summer season grass sward height and content of dry matter (DM) and metabolizable energy (ME) per year in the enclosures grazed by horses and in the exclosures within each enclosure mown monthly from May 2014 to September 2016. Exclosures were mown at the same spots as sward height measurements were made.
| | Enclosures | Exclosures |
|---|
| 2014 | 2015 | 2016 | 2014 | 2015 | 2016 |
|---|
| Grass sward height, cm | 8.9 ± 0.21 a | 5.9 ± 0.2 b | 5.2 ± 0.2 c | - | 7.1 ± 0.4 a | 5.4 ± 0.4 b |
| DM, kg/ha | 1560 ± 160 a | 863 ± 176 b | 770 ± 176 b | 714 ± 96 a | 760 ± 105 a | 260 ± 105 b |
| ME, MJ/ha | 13,858 ± 1293 a | 8133 ± 1416 b | 8176 ± 1466 b | 7201 ± 1044 a | 7869 ± 1144 a | 2810 ± 1144 b |
Table 5.
Content of dry matter (DM), metabolizable energy (ME) per kg organic matter (OM), crude protein (CP) as % of OM, and neutral detergent fiber (NDF) content as % of OM in four types of pasture samples collected in three enclosures (En1–En3) grazed all year round by Gotlandsruss and in three exclosures per enclosure. Samples were collected monthly between May 2014 and September 2016, except for December 2014–March 2015 and January–April 2016. LSmeans ± SE, p-values indicate the general effect of enclosure.
Table 5.
Content of dry matter (DM), metabolizable energy (ME) per kg organic matter (OM), crude protein (CP) as % of OM, and neutral detergent fiber (NDF) content as % of OM in four types of pasture samples collected in three enclosures (En1–En3) grazed all year round by Gotlandsruss and in three exclosures per enclosure. Samples were collected monthly between May 2014 and September 2016, except for December 2014–March 2015 and January–April 2016. LSmeans ± SE, p-values indicate the general effect of enclosure.
| Sample Type | En1 | En2 | En3 | p |
|---|
| Exclosure | | | | |
| ME, MJ/kg OM | 10.8 ± 0.2 | 10.9 ± 0.2 | 11.2 ± 0.2 | 0.2925 |
| CP, % of OM | 14.9 ± 0.4 | 14.3 ± 0.4 | 13.9 ± 0.4 | 0.1955 |
| NFD, % of OM | 55.0 ± 1.1 a | 54.7 ± 1.1 a,b | 51.8 ± 1.1 b | 0.0924 |
| DM, % | 27 ± 1 | 29 ± 1 | 26 ± 1 | 0.2722 |
| Enclosure Graze | | | | |
| ME, MJ/kg OM | 10.4 ± 0.2 a | 10.7 ± 0.2 a | 11.5 ± 0.2 b | 0.0029 |
| CP, % of OM | 16.9 ± 0.8 a,b | 16.0 ± 0.8 a | 18.8 ± 0.8 b | 0.0427 |
| NFD, % of OM | 52.7 ± 1.4 a | 53.9 ± 1.3 a | 45.6 ± 1.3 b | 0.0001 |
| DM, % | 30 ± 1 a,b | 31 ± 1 a | 27 ± 1 b | 0.0159 |
| Enclosure Forage + Volume | | | | |
| ME, MJ/kg OM | 10.1 ± 0.1 a | 9.3 ± 0.1 b | 10.6 ± 0.1 c | <0.0001 |
| CP, % of OM | 11.5 ± 0.4 a | 11.9 ± 0.4 a,b | 12.5 ± 0.4 b | 0.1405 |
| NFD, % of OM | 54.7 ± 1.1 a | 59.9 ± 1.1 b | 54.2 ± 1.1 a | 0.0007 |
| DM, % | 34 ± 1 a,b | 37 ± 1 a | 31 ± 1 b | 0.0083 |
Table 6.
Content of dry matter (DM), organic matter (OM), and crude protein (CP) as % of DM and as % of OM in feces from Gotlandsruss, divided equally between three enclosures (En1–En3) and grazing all year round.
Table 6.
Content of dry matter (DM), organic matter (OM), and crude protein (CP) as % of DM and as % of OM in feces from Gotlandsruss, divided equally between three enclosures (En1–En3) and grazing all year round.
| | En1 | En2 | En3 | p |
|---|
| DM, % | 20 ± 0.3 a | 19 ± 0.3 b | 21 ± 0.3 a | <0.0001 |
| OM, % | 79 ± 0.5 a | 80 ± 0.5 a | 75 ± 0.5 b | <0.0001 |
| CP, % of DM | 8.4 ± 0.2 a | 9.1 ± 0.2 b | 9.7 ± 0.2 c | <0.0001 |
| CP, % of OM | 10.8 ± 0.3 a | 11.5 ± 0.3 b | 13.1 ± 0.3 c | <0.0001 |
Table 7.
Correlation coefficient (r, p-value) between nutrient concentrations in graze samples and in fecal samples from 12 Gotlandsruss kept in three enclosures between May 2014 and September 2016 without supplementary feeding. DM = dry matter, OM = organic matter, CP = crude protein, VOS = digestibility coefficient of organic matter, NDF = neutral detergent fiber.
Table 7.
Correlation coefficient (r, p-value) between nutrient concentrations in graze samples and in fecal samples from 12 Gotlandsruss kept in three enclosures between May 2014 and September 2016 without supplementary feeding. DM = dry matter, OM = organic matter, CP = crude protein, VOS = digestibility coefficient of organic matter, NDF = neutral detergent fiber.
| Graze Samples | | Fecal Samples |
|---|
| | DM | OM | CP, % of DM | CP, % of OM |
|---|
| DM | r | 0.37703 | ns | −0.30271 | −0.28796 |
| p | 0.0032 | 0.0198 | 0.0270 |
| VOS | r | −0.58502 | −0.32107 | 0.62955 | 0.64978 |
| p | <0.0001 | 0.0132 | <0.0001 | <0.0001 |
| NDF | r | 0.36083 | 0.53560 | −0.69221 | −0.75195 |
| p | 0.0050 | <0.0001 | <0.0001 | <0.0001 |
| OM | r | −0.41346 | 0.36627 | ns | ns |
| p | 0.0011 | 0.0043 |
| CP, % of DM | r | −0.35722 | −0.48196 | 0.60511 | 0.66090 |
| p | 0.0055 | 0.0001 | <0.0001 | <0.0001 |
| CP, % of OM | r | ns | −0.55235 | 0.49911 | 0.57349 |
| p | <0.0001 | <0.0001 | <0.0001 |
| NDF, % of OM | r | 0.48350 | 0.46697 | −0.75952 | −0.80312 |
| p | 0.0001 | 0.0002 | <0.0001 | <0.0001 |
Table 8.
Correlation coefficient (r, p-value) between nutrient concentrations in forage + volume samples and in fecal samples from 12 Gotlandsruss kept in three enclosures between May 2014 and September 2016 without supplementary feeding. DM = dry matter, OM = organic matter, CP = crude protein, VOS = digestibility coefficient of organic matter, NDF = neutral detergent fiber.
Table 8.
Correlation coefficient (r, p-value) between nutrient concentrations in forage + volume samples and in fecal samples from 12 Gotlandsruss kept in three enclosures between May 2014 and September 2016 without supplementary feeding. DM = dry matter, OM = organic matter, CP = crude protein, VOS = digestibility coefficient of organic matter, NDF = neutral detergent fiber.
| Volume + Forage Samples | Fecal Samples |
|---|
| | DM | OM | CP, % of DM | CP, % of OM |
|---|
| DM, % | r | 0.58646 | ns | −0.45184 | −0.44157 |
| p | <0.0001 | 0.0003 | 0.0005 |
| VOS | r | −0.62653 | −0.44629 | 0.75445 | 0.76465 |
| p | <0.0001 | 0.0004 | <0.0001 | <0.0001 |
| NDF, % | r | 0.57023 | 0.48945 | −0.84249 | −0.86310 |
| p | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| OM, % | r | ns | ns | ns | ns |
| p |
| CP, % of DM | r | −0.38627 | −0.43487 | 0.64359 | 0.67240 |
| p | 0.0025 | 0.0006 | <0.0001 | <0.0001 |
| CP, % of OM | r | −0.38872 | −0.42891 | 0.64552 | 0.67353 |
| p | 0.0023 | 0.0007 | <0.0001 | <0.0001 |
| NDF, % of OM | r | 0.56563 | 0.50105 | −0.84228 | −0.86426 |
| p | <0.0001 | <0.0001 | <0.0001 | <0.0001 |






{kind=link}
{kind=link}
{kind=link}