Analysis of 19 Minerals and Cortisol in Red Deer Hair in Two Different Areas of the Stelvio National Park: A Preliminary Study

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
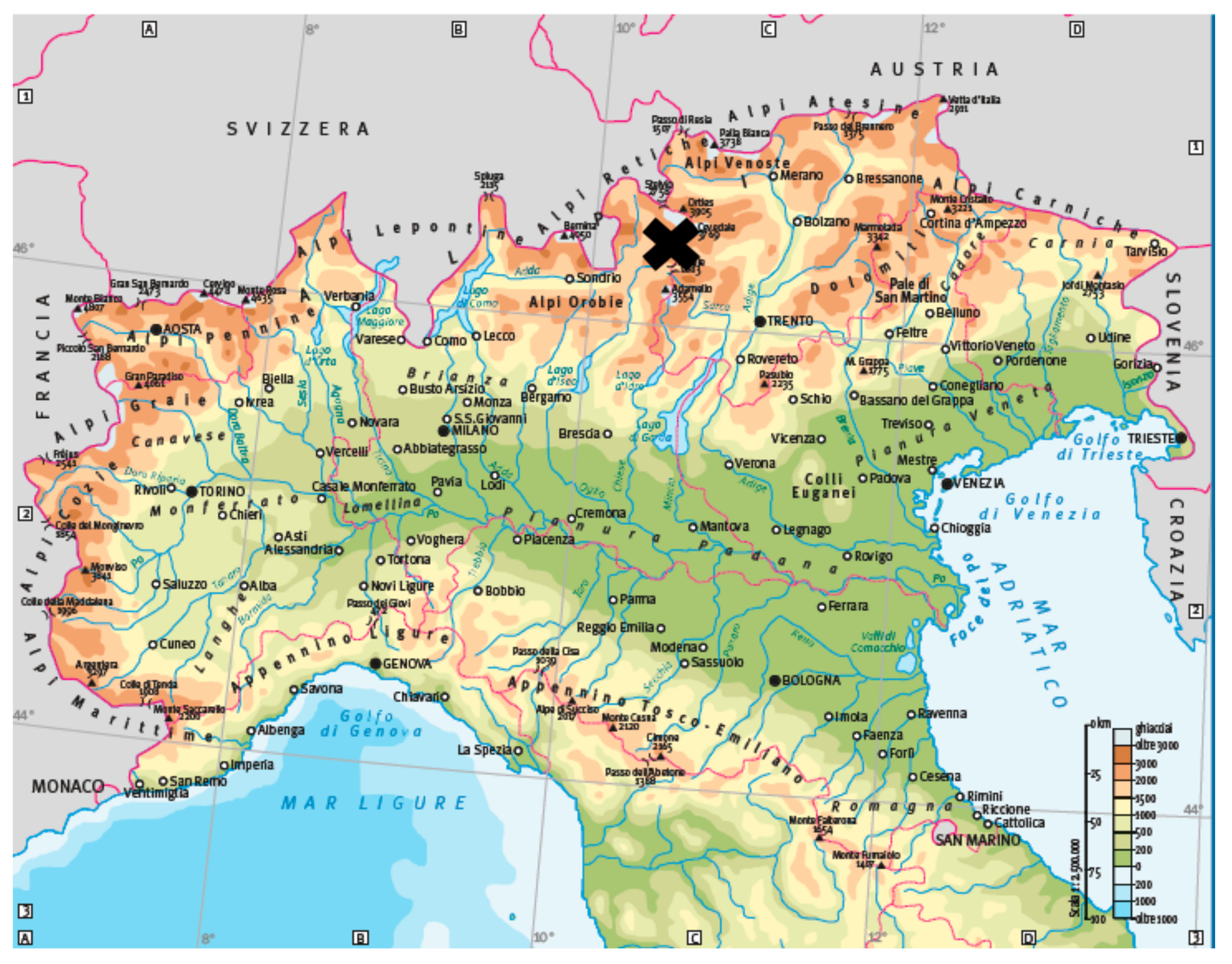
2.2. Study Area
2.3. Hair Sampling and Data Collection
2.4. Hair Washing Procedure
2.5. Hair Minerals Assay
2.6. Hair Cortisol Assay
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shankar, A.H.; Prasad, A.S. Zinc and immune function: The biological basis of altered resistance to infection. Am. J. Clin. Nutr. 1998, 68, 447–463. [Google Scholar] [CrossRef] [PubMed]
- Rabiee, A.R.; Lean, I.J.; Stevenson, M.A.; Socha, M.T. Effects of feeding organic trace minerals on milk production and reproductive performance in lactating dairy cows: A meta-analysis. J. Dairy Sci. 2010, 93, 4239–4251. [Google Scholar] [CrossRef]
- Enjalbert, F.; Lebreton, P.; Salat, O. Effects of copper, zinc and selenium status on performance and health in commercial dairy and beef herds: Retrospective study. J. Anim. Physiol. Anim. Nutr. 2006, 90, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Joksimovi, M.; Davidovi, V.; Bojani, M. The effects of some microelements supplementation—Selenium, zinc and copper into dairy cows feeds on their health and the role of selenium, zinc and copper in preserving the health status of dairy cows. Biotechnol. Anim. Husb. 2016, 32, 101–110. [Google Scholar] [CrossRef]
- Greenbaum, L.A. Micronutrient mineral deficiencies. In Nelson Textbook of Pediatrics, 18th ed.; Kliegman, R., Behrman, R., Jenson, H., Stanton, B., Saunders, W.B., Eds.; Co Philadelphia: Philadelphia, PA, USA, 2007. [Google Scholar]
- W.H.O. (Word Health Organization). Trace Elements in Human Nutrition and Health; W.H.O.: Geneva, Switzerland, 1996. [Google Scholar]
- Musso, C.G. Magnesium metabolism in health and disease. Int. Urol. Nephrol. 2009, 41, 357–362. [Google Scholar] [CrossRef]
- Peacock, M. Calcium metabolism in health and disease. Clin. J. Am. Soc. Nephrol. 2010, 5, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Flueck, W.T. Effect of trace elements on population dynamics: Selenium deficiency in free-ranging black-tailed deer. Ecology 1994, 75, 807–812. [Google Scholar] [CrossRef]
- Frank, A. “Mysterious” moose disease in Sweden. Similarities to copper deficiency and/or molybdenosis in cattle and sheep. Biochemical background of clinical signs and organ lesions. Sci. Total Environ. 1998, 209, 17–26. [Google Scholar] [CrossRef]
- Frank, A. A review of the “mysterious” wasting disease in Swedish moose (Alces alces L.) related to molybdenosis and disturbances in copper metabolism. Biol. Trace Elem. Res. 2004, 102, 143–159. [Google Scholar] [CrossRef]
- Grace, N.D.; Wilson, P.R. Trace element metabolism, dietary requirements, diagnosis and prevention of deficiencies in deer. N. Z. Vet. J. 2002, 50, 252–259. [Google Scholar] [CrossRef]
- Johnson, H.E.; Bleich, V.C.; Krausman, P.R. Mineral Deficiencies in Tule Elk, Owens Valley, California. J. Wildl. Dis. 2007, 43, 61–74. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, T.M.; Carroll, G.; Barboza, P.; Mueller, K.; Blake, J.; Woshner, V.; Willetto, C.; Willetto, C. Mineral and heavy metal status as related to a mortality event and poor recruitment in a moose population in alaska mortality event and poor recruitment in a moose population in Alaska. J. Wildl. Dis. 2001, 37, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Roug, A.; Swift, P.K.; Gerstenberg, G.; Woods, L.W.; Kreuder-johnson, C.; Torres, S.G.; Puschner, B. Comparison of trace mineral concentrations in tail hair, body hair, blood, and liver of mule deer (Odocoileus hemionus) in California. J. Vet. Diagn. Investig. 2016, 27, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Exp. Suppl. 2012, 101, 133–164. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P. An insight of environmental contamination of arsenic on animal health. Emerg. Contam. 2017, 3, 17–22. [Google Scholar] [CrossRef]
- Jones, R.L.; Weeks, H.P. Calcium, Magnesium, and Potassium in Hair of Deer from Areas of Contrasting Soil Productivity. Biol. Trace Elem. Res. 1998, 62, 155–166. [Google Scholar] [CrossRef]
- Jones, R.L. Zinc, Iron, and Sodium in Hair of Deer from Areas of Contrasting Soil Productivity. Biol. Trace Elem. Res. 2002, 86, 217–226. [Google Scholar] [CrossRef]
- Ramanzin, M.; Amici, A.; Casoli, C.; Esposito, L.; Lupi, P.; Marsico, G.; Mattiello, S.; Oliviero, O.; Ponzetta, M.P.; Russo, C.; et al. Meat from wild ungulates: Ensuring quality and hygiene of an increasing resource meat from wild ungulates. Ital. J. Anim. Sci. 2010, 9, 318–331. [Google Scholar] [CrossRef]
- Jovanovići, B.I.; Veličković, M.; Milanović, S.; Valčić, O.; Gvozdić, D.; Vranješ-Đurić, S. Supplemental selenium reduces the levels of biomarkers of oxidative and general stress in peripartum dairy cows. Acta Vet. 2015, 65, 191–201. [Google Scholar] [CrossRef]
- Pechova, A.; Pavlata, L. Chromium as an essential nutrient: A review. Vet. Med. 2007, 52, 1–18. [Google Scholar] [CrossRef]
- Gubareva, L.; Soloviev, A.; Bicheva, G.; Ermolova, L. Combined influence of hypo and hypermicroelementosis on functioning of cardiovascular and endocrine systems and anxiety level of adolescents. Hum. Ecol. 2017, 8, 29–36. [Google Scholar] [CrossRef]
- Vanaelst, B. The Effect of Chronic Stress on Body Composition and Mineral Status among Children. Use of Hair as a Biological Matrix. Master’s Thesis, Ghent University, Ghent, Belgium, 2013. [Google Scholar]
- Costantini, D.; Marasco, V.; Møller, A.P. A meta-analysis of glucocorticoids as modulators of oxidative stress in vertebrates. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2011, 181, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Vertuani, S.; Angusti, A.; Manfredini, S. The antioxidants and pro-antioxidants network: An overview. Curr. Pharm. Des. 2004, 10, 1677–1694. [Google Scholar] [CrossRef] [PubMed]
- Marieb, E.N.; Hoehn, K. Human Anatomy & Physiology, 10th ed.; Pearson: London, UK, 2010. [Google Scholar]
- McEwen, B.S. Allostasis and Allostatic Load: Implications for Neuropsychopharmacology. Neuropsychopharmacology 2000, 22, 108–124. [Google Scholar] [CrossRef]
- Moberg, G.P.; Mench, J.A. Biological response to stress: Implications for animal welfare. In The Biology of Animal Stress. Basic Implications for Animal Welfare, 1st ed.; CABI Publishing: Wallingford, UK, 2000; pp. 1–22. [Google Scholar]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the Stress Response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef] [PubMed]
- Macbeth, B.J.; Cattet, M.R.L.; Stenhouse, G.B.; Gibeau, M.L.; Janz, D.M. Hair cortisol concentration as a noninvasive measure of long-term stress in free-ranging grizzly bears (Ursus arctos): Considerations with implications for other wildlife. Can. J. Zool. 2010, 88, 935–949. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.K. Stress, Food, and Inflammation: Psychoneuroimmunology and Nutrition at the Cutting Edge. Psychosom. Med. 2010, 72, 365–369. [Google Scholar] [CrossRef]
- Yin, J.; Levanon, D.; Chen, J.D.Z. Inhibitory effects of stress on postprandial gastric myoelectrical activity and vagal tone in healthy subjects. Neurogastroenterol. Motil. 2004, 16, 737–744. [Google Scholar] [CrossRef]
- Mayer, E. The neurobiology of stress and gastrointestinal disease. Gut 2000, 47, 861–869. [Google Scholar] [CrossRef]
- Heshmati, H.M.; Riggs, B.L.; Burritt, M.F.; Mcalister, C.A.; Wollan, P.C.; Khosla, S. Effects of the circadian variation in serum cortisol on markers of bone turnover and calcium homeostasis in normal postmenopausal women. J. Clin. Endocrinol. Metab. 1998, 83, 751–756. [Google Scholar] [CrossRef]
- Caslini, C.; Comin, A.; Peric, T.; Prandi, A.; Pedrotti, L.; Mattiello, S. Use of hair cortisol analysis for comparing population status in wild red deer (Cervus elaphus) living in areas with different characteristics. Eur. J. Wildl. Res. 2016, 62, 713–723. [Google Scholar] [CrossRef]
- Kirschbaum, C.; Tietze, A.; Skoluda, N.; Dettenborn, L. Hair as a retrospective calendar of cortisol production. Increased cortisol incorporation into hair in the third trimester of pregnancy. Psychoneuroendocrinology 2009, 34, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Russell, E.; Koren, G.; Rieder, M.; Uum, S. Van Hair cortisol as a biological marker of chronic stress: Current status, future directions and unanswered questions. Psychoneuroendocrinology 2012, 37, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Peric, T.; Comin, A.; Corazzin, M.; Montillo, M.; Cappa, A.; Campanile, G.; Prandi, A. Short communication: Hair cortisol concentrations in Holstein-Friesian and crossbreed F1 heifers. J. Dairy Sci. 2013, 96, 3023–3027. [Google Scholar] [CrossRef] [PubMed]
- Peric, T.; Corazzin, M.; Romanzin, A.; Bovolenta, S.; Prandi, A.; Montillo, M.; Comin, A. Cortisol and DHEA concentrations in the hair of dairy cows managed indoor or on pasture. Livest. Sci. 2017, 202, 39–43. [Google Scholar] [CrossRef]
- Burnard, C.; Ralph, C.; Hynd, P.; Hocking, J.E.; Tilbrook, A. Hair cortisol and its potential value as a physiological measure of stress response in human and non-human animals. Anim. Prod. Sci. 2016, 57, 401–414. [Google Scholar] [CrossRef]
- Meyer, J.S.; Novak, M.A. Minireview: Hair Cortisol: A Novel Biomarker of Hypothalamic-Pituitary-Adrenocortical Activity. Endocrinology 2012, 153, 4120–4127. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.C.; White, C.D.; Van Uum, S.; Longstaffe, F.J. Integrating cortisol and isotopic analyses of archeological hair: Reconstructing individual experiences of health and stress. Am. J. Phys. Anthropol. 2015, 156, 577–594. [Google Scholar] [CrossRef]
- Nowak, B.; Kozłowski, H. Heavy metals in human hair and teeth: The correlation with metal concentration in the environment. Biol. Trace Elem. Res. 1998, 62, 213–228. [Google Scholar] [CrossRef]
- Nowak, B.; Chmielnicka, J. Relationship of lead and cadmium to essential elements in hair, teeth, and nails of environmentally exposed people. Ecotoxicol. Environ. Saf. 2000, 46, 265–274. [Google Scholar] [CrossRef]
- Díaz-Barriga, F.; Santos, M.A.; Yañez, L.; Cuellar, J.A.; Ostrosky-Wegman, P.; Montero, R.; Perez, A.; Ruiz, E.; Garcia, A.; Gomez, H. Biological monitoring of workers at a recently opened hazardous waste disposal site. J. Expo. Anal. Environ. Epidemiol. 1993, 3, 63–71. [Google Scholar] [PubMed]
- Shah, F.; Gul Kazi, T.; Imran Afridi, H.; Khan, S.; Fatima Kolachi, N.; Balal Arain, M.; Ahmed Baig, J. The influence of environmental exposure on lead concentrations in scalp hair of children in Pakistan. Ecotoxicol. Environ. Saf. 2011, 74, 727–732. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. Mercury Study Report to Congress; Office of Air Quality Planning and Standards and Office of Research and Development: Washington, DC, USA, 1997. [Google Scholar]
- Flynn, A.; Franzmann, A.W.; Arneson, P.D.; Western, C. Sequential Hair Shaft as an Indicator of Prior Mineralization in the Alaskan Moose. J. Anim. Sci. 1975, 41, 906–910. [Google Scholar] [CrossRef]
- Duffy, L.K.; Hallock, R.J.; Finstad, G.; Bowyer, R.T. Noninvasive Environmental Monitoring of Mercury in Alaskan Reindeer. Am. J. Environ. Sci. 2009, 1, 249–253. [Google Scholar] [CrossRef]
- Sobańska, M.A. Wild boar hair (Sus scrofa) as a non-invasive indicator of mercury pollution. Sci. Total Environ. 2005, 339, 81–88. [Google Scholar] [CrossRef]
- Martin, J.G.A.; Réale, D.J.G. Animal temperament and human disturbance: Implications for the response of wildlife to tourism. Behav. Process. 2008, 77, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.; Marquez, M.; GochfeldJ, M. Heavy metals in the hair of opossum from Palo Verde, Costa Rica. Arch. Environ. Contam. Toxicol. 1994, 27, 472–476. [Google Scholar] [CrossRef]
- Amici, A.; Danieli, P.P.; Russo, C.; Primi, R.; Ronchi, B. Concentrations of some toxic and trace elements in wild boar (Sus scrofa) organs and tissues in different areas of the Province of Viterbo (Central Italy). Ital. J. Anim. Sci. 2012, 11, 354–362. [Google Scholar] [CrossRef]
- Mattiello, S.; Faustini, M.; Rosenthal, R.; Vigo, D.; Redaelli, W.; Maffeo, G. Minerali nel pelo del cervo (Cervus elaphus): Indagini preliminari su di una popolazione alpina. Obiettivi Doc. Vet. 1997, 11, 49–53. [Google Scholar]
- Długaszek, M.; Kopczyński, K. Correlations between elements in the fur of wild animals. Bull. Environ. Contam. Toxicol. 2014, 93, 25–30. [Google Scholar] [CrossRef]
- Crill, C.; Janz, D.M.; Kusch, J.M.; Santymire, R.M.; Heyer, G.P.; Shury, T.K.; Lane, J.E. Investigation of the utility of feces and hair as non-invasive measures of glucocorticoids in wild black-tailed prairie dogs (Cynomys ludovicianus). Gen. Comp. Endocrinol. 2019, 275, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Ventrella, D.; Elmi, A.; Barone, F.; Carnevali, G.; Govoni, N.; Bacci, M.L. Hair Testosterone and Cortisol Concentrations in Pre- and Post-Rut Roe Deer Bucks: Correlations with Blood Levels and Testicular Morphometric Parameters. Animals 2018, 6, 113. [Google Scholar] [CrossRef] [PubMed]
- Neuman-Lee, L.A.; Terletzky, P.A.; Atwood, T.C.; Gese, E.M.; Smith, G.D.; Greenfield, S.; Pettit, J.; French, S.S. Demographic and temporal variations in immunity and condition of polar bears (Ursus maritimus) from the southern Beaufort Sea. J. Exp. Zool. A Ecol. Integr. Physiol. 2017, 327, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, C.; Montillo, M.; Prandi, A.; Mkupasi, E.M.; Ngowi, H.A.; Johansen, M.V. Hair cortisol and dehydroepiandrosterone concentrations in naturally Taenia solium infected pigs in Tanzania. Gen. Comp. Endocrinol. 2017, 246, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Comin, A.; Veronesi, M.C.; Montillo, M.; Faustini, M.; Valentini, S.; Cairoli, F.; Prandi, A. Hair cortisol level as a retrospective marker of hypothalamic-pituitary-adrenal axis activity in horse foals. Vet. J. 2012, 194, 131–132. [Google Scholar] [CrossRef]
- Montillo, M.; Comin, A.; Corazzin, M.; Peric, T.; Faustini, M.; Veronesi, M.C.; Valentini, S.; Bustaffa, M.; Prandi, A. The Effect of temperature, rainfall, and light conditions on hair cortisol concentrations in newborn foals. J. Equine Vet. Sci. 2014, 34, 774–778. [Google Scholar] [CrossRef]
- Stradaioli, G.; Peric, T.; Montillo, M.; Comin, A.; Corazzin, M.; Veronesi, M.C.; Prandi, A. Hair cortisol and testosterone concentrations and semen production of Bos taurus bulls. Ital. J. Anim. Sci. 2017, 16, 631–639. [Google Scholar] [CrossRef]
- Bergamin, C.; Comin, A.; Corazzin, M.; Faustini, M.; Peric, T.; Scollo, A.; Gottardo, F.; Montillo, M.; Prandi, A. Cortisol, DHEA, and Sexual Steroid Concentrations in Fattening Pigs’ Hair. Animals 2019, 12, 345. [Google Scholar] [CrossRef]
- Comune di Valfurva. Bollettino di Informazione Municipale del Comune di Valfurva; No. 1; Comune di Valfurva: Valfurva, SO, Italy, 2011. [Google Scholar]
- Corlatti, L.; Gugiatti, A.; Pedrotti, L. Spring spotlight counts provide reliable indices to track changes in population size of mountain-dwelling red deer Cervus elaphus. Wildl. Biol. 2016, 22, 268–276. [Google Scholar] [CrossRef]
- Anonymous. Primo Documento Illustrativo del Piano del Parco Nazionale dello Stelvio; ERSAF/Regione Lombardia: Milano, Italy, 2018. [Google Scholar]
- Doglioni, G.; Flores, G. An Introduction to the Italian Geology; Lamisco: Potenza, Italy, 1997. [Google Scholar]
- Carro, M.; Pedrotti, L. Atlante del Parco Nazionale dello Stelvio; Consorzio del Parco Nazionale dello Stelvio: Bormio, Italy, 2010. [Google Scholar]
- ARPA (Agenzia Regionale per la Protezione Ambientale). Rapporto sullo Stato dell’Ambiente in Provincia di Sondrio—Anni 2007–2008; Agenzia Regionale per la Protezione dell’Ambiente della Lombardia—Tipografia Ignizio: Montagna in Valtellina, Sondrio, Italy, 2010. [Google Scholar]
- Mattioli, S.; De Marinis, A.M. Guida al Rilevamento Biometrico degli Ungulati; Istituto Superiore per la Protezione e la Ricerca Ambientale, Documenti Tecnici: Roma, Italy, 2009. [Google Scholar]
- Monson, R.A.; Stone, W.B.; Weber, B.L.; Spadaro, F.I. Comparison of Riney and total kidney fat techniques for evaluating the physical condition of white-tailed deer. N. Y. Fish Game J. 1974, 21, 67–72. [Google Scholar]
- Monaco, A.; Franzetti, B.; Pedrotti, L.; Toso, S. Linee Guida per la Gestione del Cinghiale; Ministero delle Politiche Agricole e Forestali—Istituto Nazionale per la Fauna Selvatica: Roma, Italy, 2003. [Google Scholar]
- Milner, J.M.; Bonenfant, C.; Mysterud, A.; Gaillard, J.M.; Csányi, S.; Stenseth, N.C. Temporal and spatial development of red deer harvesting in Europe: Biological and cultural factors. J. Appl. Ecol. 2006, 43, 721–734. [Google Scholar] [CrossRef]
- Carnevali, L.; Pedrotti, L.; Riga, F.; Toso, S. Banca Dati Ungulati: Status, distribuzione, consistenza, gestione e prelievo venatorio delle popolazioni di Ungulati in Italia. Rapporto 2001–2005. Biol. Cons. Fauna 2009, 117, 1–168. [Google Scholar]
- Mattiello, S.; Redaelli, W.; Miro, C.; Crimella, M.C.; Carenzi, C. Dairy Cattle Husbandry and Red Deer Utilization of a Summer Range in the Central Italian Alps. Mt. Res. Dev. 2003, 34, 161–168. [Google Scholar] [CrossRef]
- Gupta, N.; Yadav, K.K.; Kumar, V.; Kumar, S.; Chadd, R.P.; Kumar, A. Trace elements in soil-vegetables interface: Translocation, bioaccumulation, toxicity and amelioration—A review. Sci. Total Environ. 2019, 651, 2927–2942. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.; Mittal, M.; Mehta, A. Heavy metal induced oxidative stress & its possible reversal by chelation therapy. Indian J. Med. Res. 2008, 128, 501–523. [Google Scholar] [PubMed]
- Jan, A.T.; Ali, A.; Haq, Q. Glutathione as an antioxidant in inorganic mercury induced nephrotoxicity. J. Postgrad. Med. 2011, 57, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Pompilio, L.; Meriggi, A. Modelling wild ungulate distribution in alpine habitat: A case study. Ital. J. Zool. 2001, 68, 281–289. [Google Scholar] [CrossRef]
- Seelig, M.S. Consequences of magnesium deficiency on the enhancement of stress reactions; preventive and therapeutic implications (a review). J. Am. Coll. Nutr. 1994, 13, 429–446. [Google Scholar] [CrossRef]
- Grases, G.; Sanchis, P.; Casero, A.; Perelló, J.; Isern, B.; Rigo, E. Anxiety and stress among science students. Study of calcium and magnesium alterations. Magnes. Res. 2006, 19, 102–106. [Google Scholar]
- Moore, R.J.; Friedl, K.E.; Tulley, R.T.; Askew, E.W. Maintenance of iron status in healthy men during an extended period of stress and physical activity. Am. J. Clin. Nutr. 1993, 58, 923–927. [Google Scholar] [CrossRef]
- Singh, A.; Smoak, B.L.; Patterson, K.Y.; LeMay, L.G.; Veillon, C.; Deuster, P.A. Biochemical indices of selected trace minerals in men: Effect of stress. Am. J. Clin. Nutr. 1991, 53, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Evers, S.E.; Avison, W.R.; Campbell, M.K. Higher zinc intake buffers the impact of stress on depressive symptoms in pregnancy. Nutr. Res. 2010, 30, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shen, H.; Chen, C.; Wang, W.; Yu, S.; Zhao, M.; Li, M. The effect of psychological stress on iron absorption in rats. BMC Gastroenterol. 2009, 9, 83. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| SE Area | NW Area | p-Value a | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | Unit | n | Mean | SEM | Min | Max | n | Mean | SEM | Min | Max | |
| Cortisol | pg/mg | 16 | 5.18 | 0.34 | 3.41 | 7.18 | 15 | 6.97 | 0.63 | 4.33 | 11.61 | 0.063 |
| Li | μg/g | 16 | 0.53 | 0.11 | 0.15 | 1.55 | 15 | 0.31 | 0.07 | 0.00 | 1.02 | 0.206 |
| B | μg/g | 8 | 0.87 | 0.18 | 0.26 | 1.78 | 5 | 1.88 | 0.87 | 0.11 | 4.16 | 0.770 |
| Al | μg/g | 15 | 242.98 | 41.38 | 89.30 | 726.14 | 15 | 137.62 | 20.69 | 11.39 | 286.42 | 0.026 * |
| Ti | μg/g | 16 | 4.24 | 0.43 | 0.29 | 6.59 | 15 | 3.95 | 0.57 | 0.82 | 8.47 | 0.477 |
| V | μg/g | 16 | 0.75 | 0.16 | 0.14 | 1.88 | 15 | 0.39 | 0.07 | 0.03 | 1.01 | 0.220 |
| Cr | μg/g | 16 | 0.53 | 0.05 | 0.16 | 0.93 | 15 | 0.42 | 0.07 | 0.17 | 1.28 | 0.053 |
| Fe | μg/g | 16 | 222.68 | 25.48 | 44.22 | 410.66 | 15 | 216.58 | 50.95 | 59.98 | 764.89 | 0.220 |
| Ni | μg/g | 16 | 1.32 | 0.28 | 0.27 | 3.97 | 14 | 0.64 | 0.18 | 0.14 | 2.67 | 0.022 * |
| Co | μg/g | 16 | 0.18 | 0.02 | 0.06 | 0.35 | 14 | 0.09 | 0.01 | 0.05 | 0.15 | 0.003 ** |
| Cu | μg/g | 16 | 5.03 | 0.29 | 2.94 | 7.40 | 15 | 4.07 | 0.19 | 2.55 | 5.25 | 0.007 ** |
| Zn | μg/g | 16 | 70.43 | 1.15 | 64.74 | 83.04 | 15 | 65.41 | 2.07 | 45.71 | 74.55 | 0.123 |
| As | μg/g | 16 | 1.39 | 0.17 | 0.44 | 2.88 | 15 | 0.72 | 0.10 | 0.18 | 1.56 | 0.003 ** |
| Ag | μg/g | 8 | 0.05 | 0.01 | 0.01 | 0.10 | 9 | 0.07 | 0.02 | 0.01 | 0.18 | 0.630 |
| Cd | μg/g | 7 | 2.32 | 0.42 | 0.60 | 3.50 | 12 | 0.34 | 0.12 | 0.00 | 1.00 | 0.001 ** |
| Sn | μg/g | 11 | 0.14 | 0.03 | 0.04 | 0.33 | 9 | 0.12 | 0.03 | 0.03 | 0.33 | 0.494 |
| Sb | μg/g | 14 | 0.03 | 0.01 | 0.01 | 0.09 | 10 | 0.07 | 0.02 | 0.01 | 0.21 | 0.639 |
| Ba | μg/g | 15 | 2.24 | 0.29 | 0.81 | 4.66 | 15 | 2.04 | 0.45 | 0.20 | 7.09 | 0.419 |
| Tl | μg/g | 16 | 1.99 | 0.67 | 0.00 | 6.59 | 15 | 1.31 | 0.56 | 0.00 | 7.58 | 0.874 |
| Pb | μg/g | 12 | 0.75 | 0.16 | 0.08 | 1.69 | 11 | 0.35 | 0.08 | 0.01 | 1.03 | 0.110 |
| SE Area | NW Area | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Unit | n | Mean | SEM | n | Mean | SEM | p-Value a |
| Calves | ||||||||
| body length | cm | 7 | 126.00 | 4.17 | 8 | 134.87 | 3.21 | 0.111 |
| foot length | cm | 7 | 42.86 | 0.68 | 8 | 44.81 | 0.72 | 0.073 |
| height at withers | cm | 7 | 90.86 | 1.10 | 8 | 94.56 | 1.22 | 0.044 * |
| jaw length | mm | 6 | 207.14 | 1.94 | 8 | 206.22 | 3.96 | 0.856 |
| carcass weight | kg | 7 | 30.00 | 2.027 | 8 | 33.31 | 3.00 | 0.391 |
| kidney fat index | KFI | 6 | 24.24 | 6.66 | 8 | 22.37 | 3.38 | 0.792 |
| Yearlings | ||||||||
| body length | cm | 4 | 148.00 | 3.76 | 2 | 145.00 | 5.00 | 0.665 |
| foot length | cm | 4 | 48.12 | 1.23 | 2 | 47.75 | 0.75 | 0.853 |
| height at withers | cm | 4 | 104.50 | 4.11 | 2 | 105.50 | 1.50 | 0.880 |
| jaw length | mm | 4 | 243.10 | 6.37 | 2 | 225.20 | 9.70 | 0.187 |
| carcass weight | kg | 4 | 50.75 | 3.04 | 2 | 44.00 | 5.00 | 0.286 |
| kidney fat index | KFI | 4 | 35.94 | 9.55 | 2 | 23.26 | 6.66 | 0.442 |
| Adult | ||||||||
| body length | cm | 5 | 162.50 | 1.05 | 5 | 166.20 | 3.20 | 0.304 |
| foot length | cm | 5 | 49.70 | 0.55 | 5 | 49.30 | 0.51 | 0.606 |
| height at withers | cm | 5 | 108.20 | 0.80 | 5 | 106.20 | 2.08 | 0.396 |
| jaw length | mm | 5 | 268.73 | 4.17 | 5 | 262.11 | 3.15 | 0.241 |
| carcass weight | kg | 5 | 58.80 | 3.83 | 5 | 59.90 | 3.38 | 0.835 |
| kidney fat index | KFI | 5 | 34.20 | 7.42 | 5 | 59.10 | 15.30 | 0.181 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montillo, M.; Caslini, C.; Peric, T.; Prandi, A.; Netto, P.; Tubaro, F.; Pedrotti, L.; Bianchi, A.; Mattiello, S. Analysis of 19 Minerals and Cortisol in Red Deer Hair in Two Different Areas of the Stelvio National Park: A Preliminary Study. Animals 2019, 9, 492. https://doi.org/10.3390/ani9080492
Montillo M, Caslini C, Peric T, Prandi A, Netto P, Tubaro F, Pedrotti L, Bianchi A, Mattiello S. Analysis of 19 Minerals and Cortisol in Red Deer Hair in Two Different Areas of the Stelvio National Park: A Preliminary Study. Animals. 2019; 9(8):492. https://doi.org/10.3390/ani9080492
Chicago/Turabian StyleMontillo, Marta, Chiara Caslini, Tanja Peric, Alberto Prandi, Paola Netto, Franco Tubaro, Luca Pedrotti, Alessandro Bianchi, and Silvana Mattiello. 2019. "Analysis of 19 Minerals and Cortisol in Red Deer Hair in Two Different Areas of the Stelvio National Park: A Preliminary Study" Animals 9, no. 8: 492. https://doi.org/10.3390/ani9080492
APA StyleMontillo, M., Caslini, C., Peric, T., Prandi, A., Netto, P., Tubaro, F., Pedrotti, L., Bianchi, A., & Mattiello, S. (2019). Analysis of 19 Minerals and Cortisol in Red Deer Hair in Two Different Areas of the Stelvio National Park: A Preliminary Study. Animals, 9(8), 492. https://doi.org/10.3390/ani9080492