Dehydrated Alfalfa and Fresh Grass Supply in Young Rabbits: Effect on Performance and Caecal Microbiota Biodiversity

 ,
,  ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
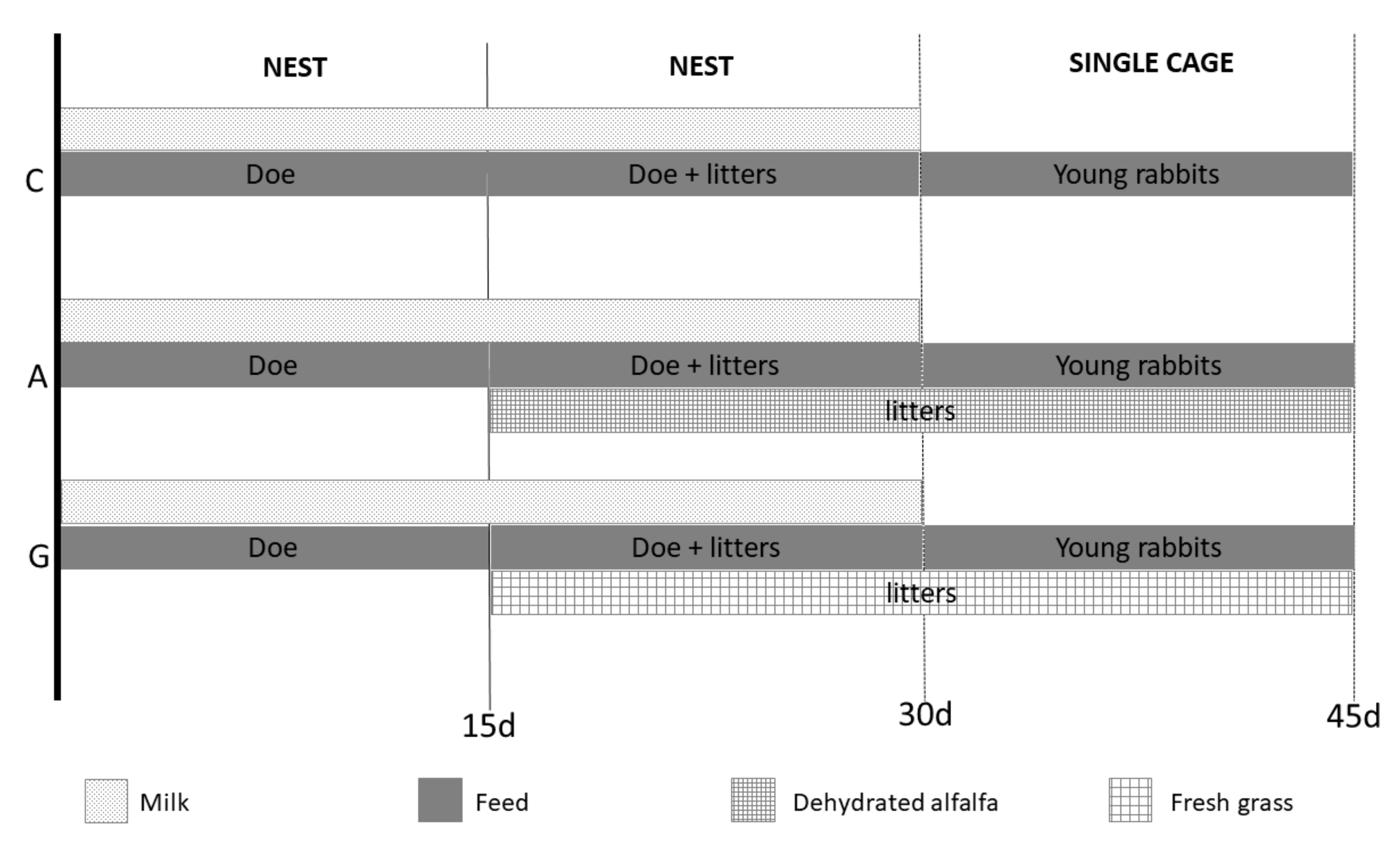
2.1. Animals and Experimental Design
- Control group (C); does and young rabbits were fed standard pelleted feed (alfalfa meal 38 g/100 g, barley 19 g/100 g, maize gluten feed 15 g/100 g, extruded soybean 14 g/100 g, wheat bran 7 g/100 g, and vitamin and mineral mix 7 g/100 g).
- A group; does and young rabbits were fed the same standard pelleted feed as group C but had additional access to dehydrated alfalfa (furnished by Carli, Pietracuta di San Leo; RN, Italy; in blocks of size 3 cm length × 2 cm width × 1 cm thickness).
- G group; does and young rabbits were fed the same standard pelleted feed as group C but had additional access to fresh grass (cut every day at an average height of 20 cm). The main species of the natural pasture were the following: Arrhenatherum elatius, Brachypodium spp., Dactylis glomerata, Lolium spp., Lotus cornicolatus, Onobrycis viciaefolia, Pisum sativum, and Capsella bursa-pastoris.
2.2. Productive Performance
2.3. Slaughtering and Sampling
2.4. Analytical Evaluations
Proximate Composition of Feed and Supplemental Diets
2.5. Caecal Microbiota Evaluation
2.5.1. DNA Extraction
2.5.2. 16S Ribosomal RNA (rRNA) Gene Sequencing
2.5.3. Sequence Analysis
2.6. Statistical Analyses
3. Results
3.1. Proximate Composition of Pelleted and Supplemental Feed
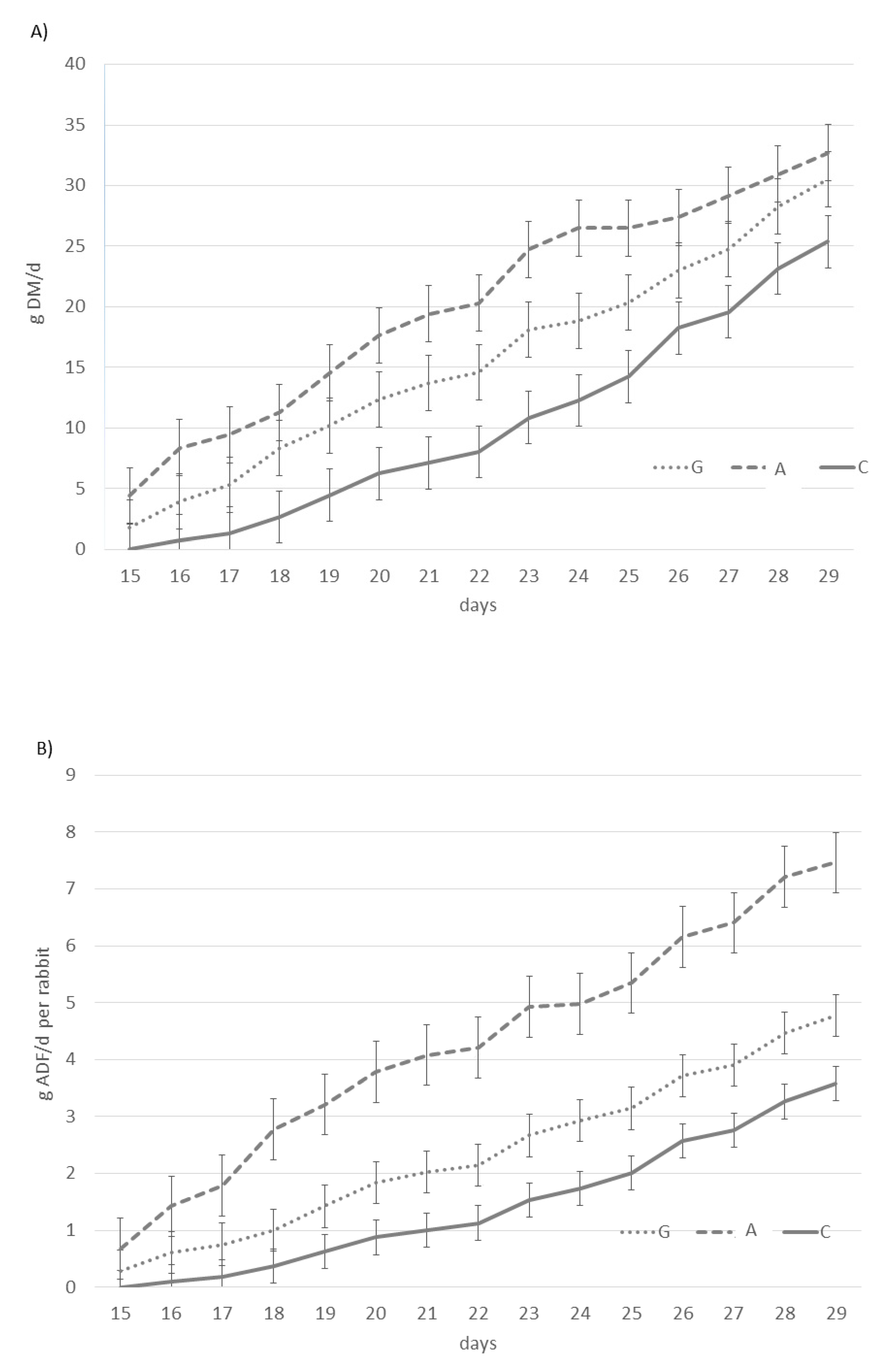
3.2. Productive Performance of Does, Litters, and Young Rabbits
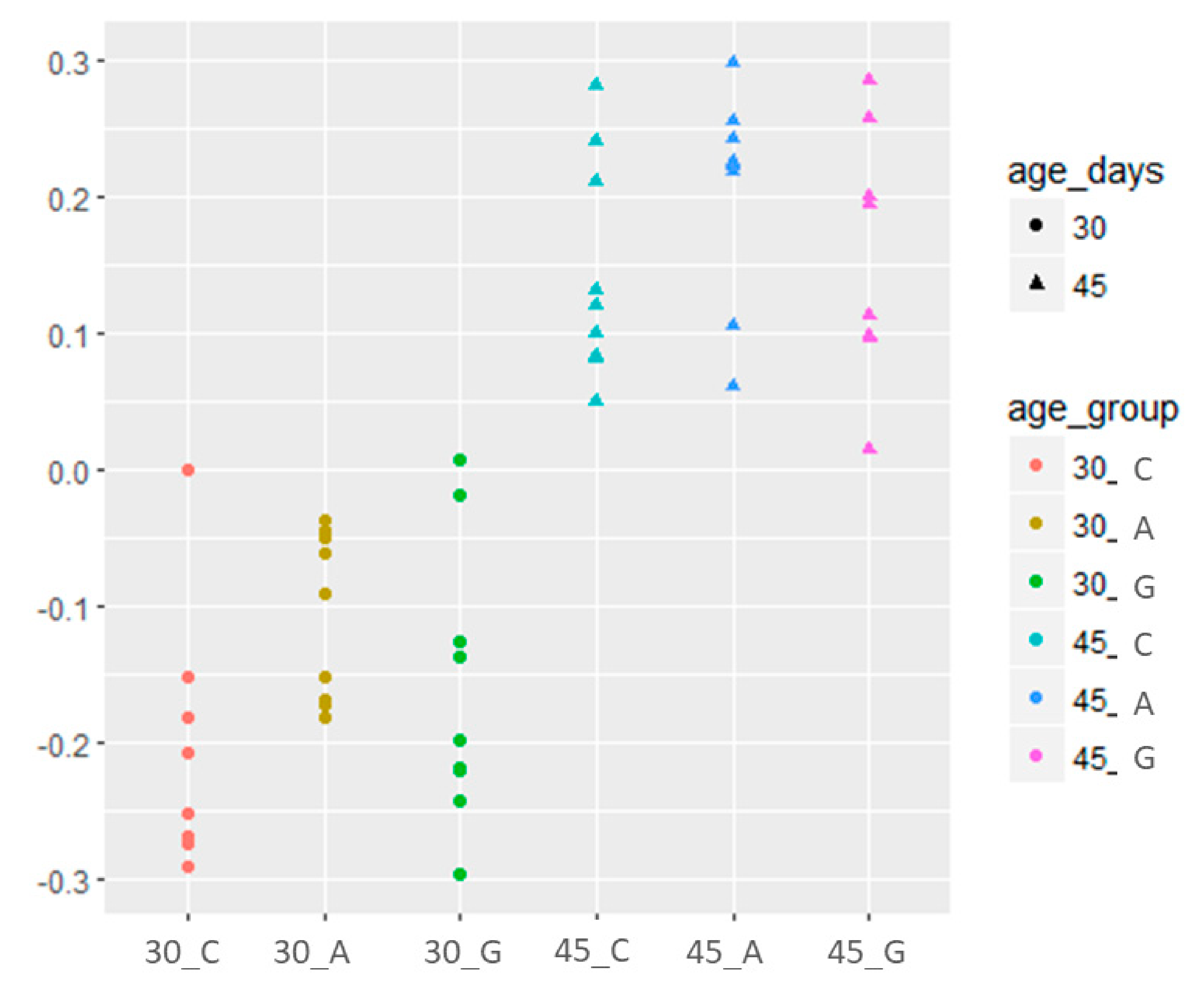
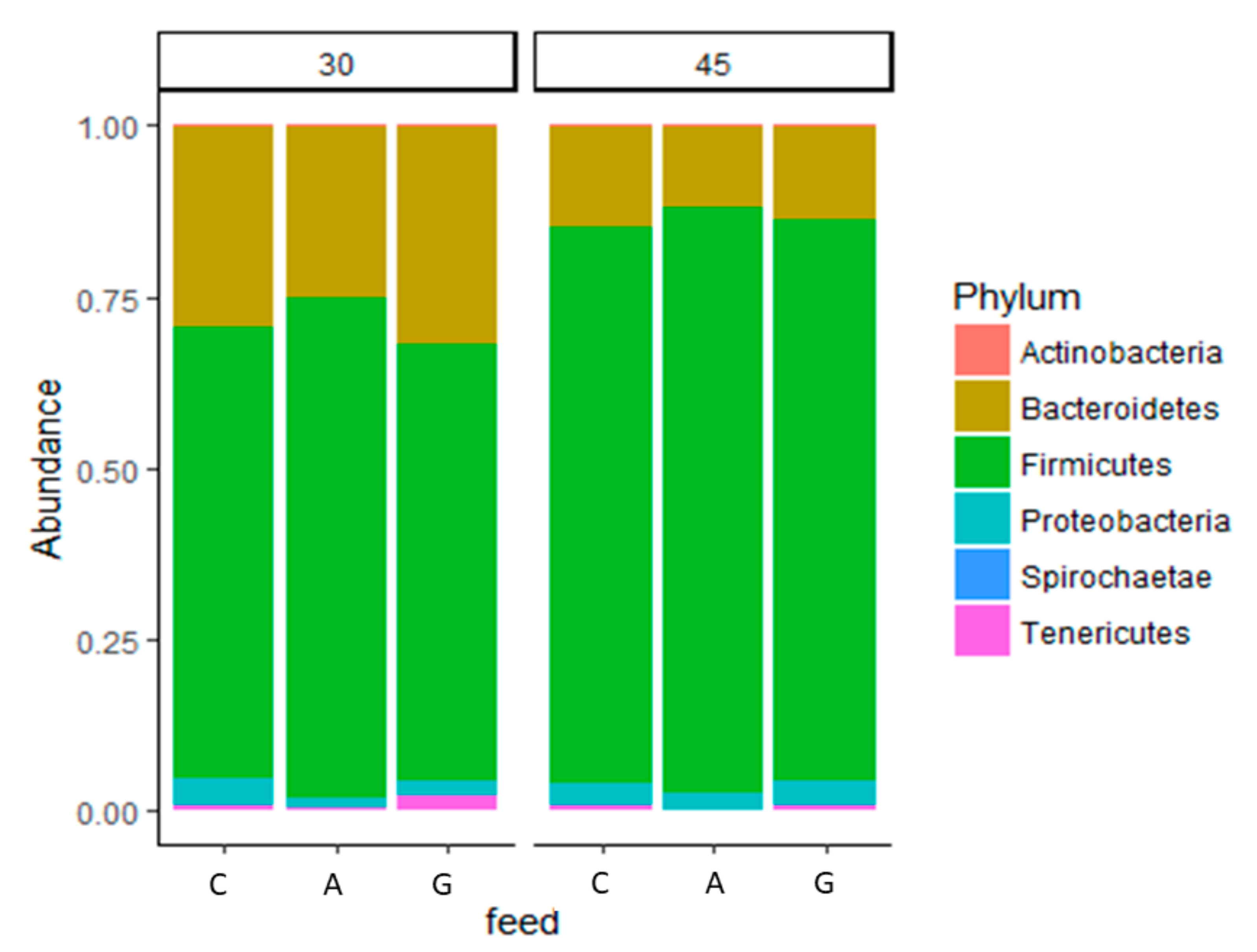
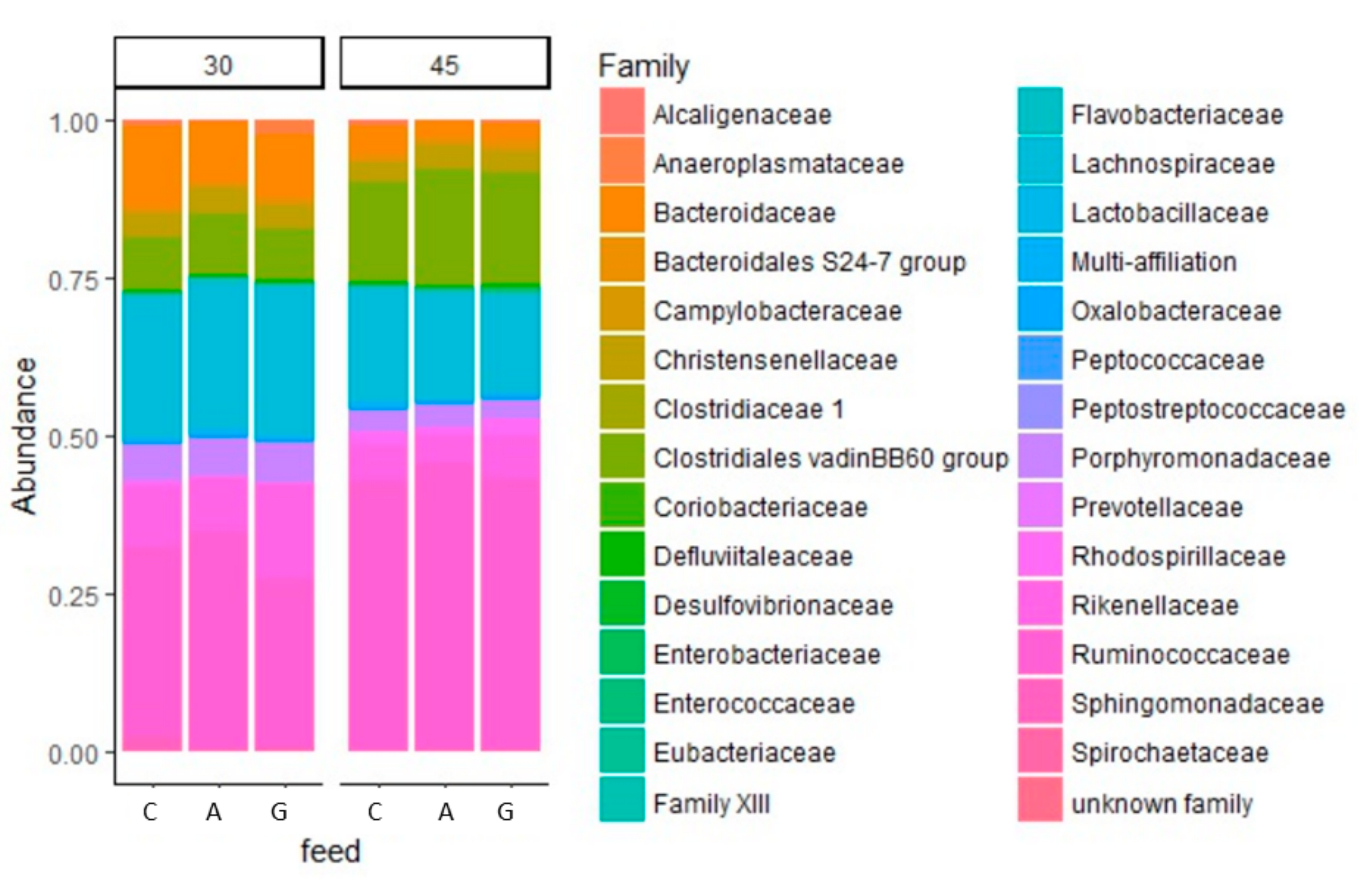
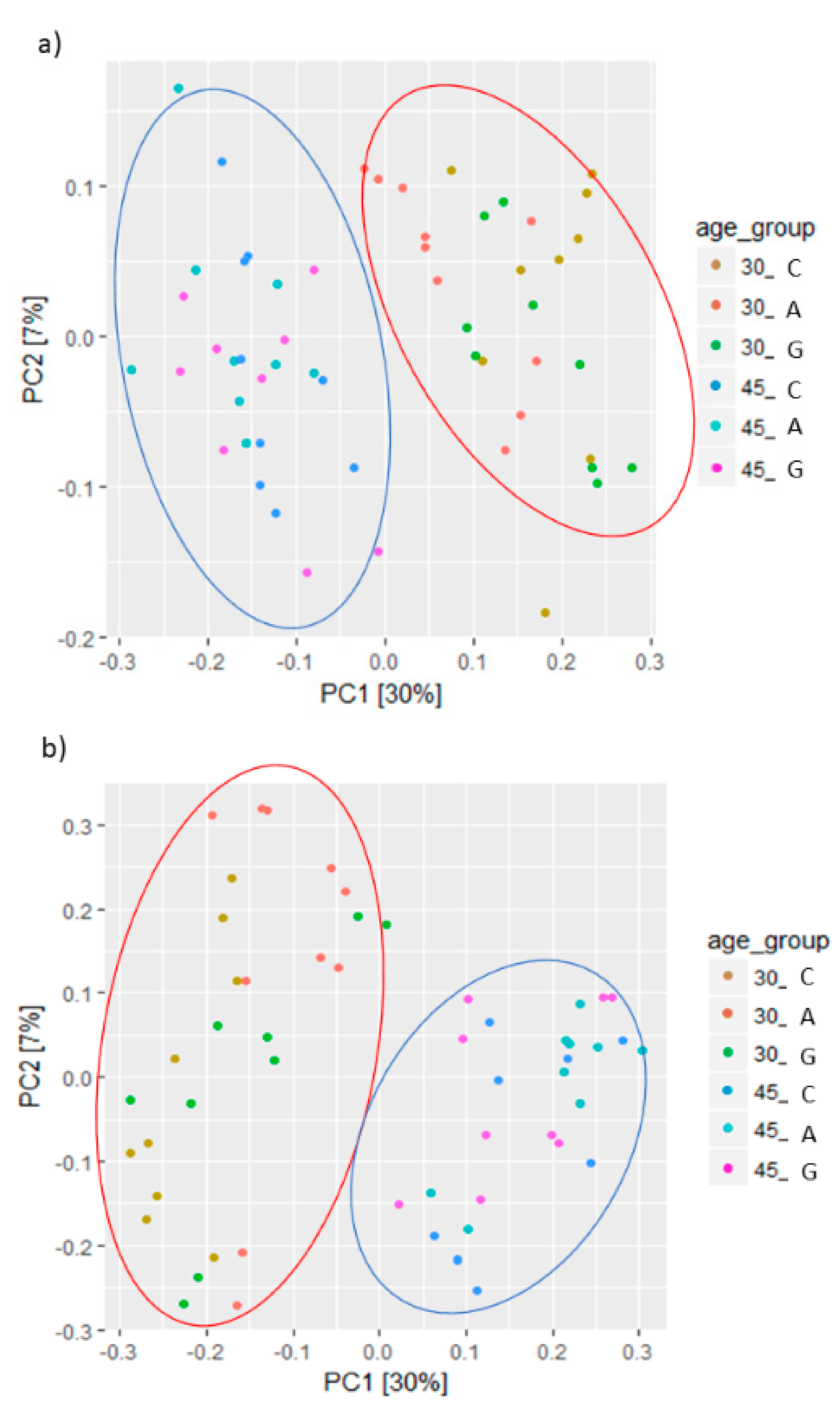
3.3. Caecal Microbiota Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed]
- Maertens, L.; Falcao-e-Cunha, L.; Marounek, M. Feed additives to reduce the use of antibiotics. In Recent Advances in Rabbit Sciences; European cooperation in the field of Scientific and Technical Research (COST): Brussels, Belgium, 2006; p. 259. [Google Scholar]
- Nogacka, A.M.; Salazar, N.; Arboleya, S.; Suárez, M.; Fernández, N.; Solís, G.; de Los Reyes-Gavilán, C.G.; Gueimonde, M. Early microbiota, antibiotics and health. Cell. Mol. Life Sci. 2018, 75, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310. [Google Scholar] [CrossRef] [PubMed]
- Argañaraz-Martínez, E.; Babot, J.D.; Apella, M.C.; Chaia, A.P. Physiological and functional characteristics of Propionibacterium strains of the poultry microbiota and relevance for the development of probiotic products. Anaerobe 2013, 23, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Verstegen, M.W.A.; Tamminga, S.; Williams, B.A. The role of the commensal gut microbial community in broiler chickens. World’s Poultry Sci. J. 2005, 61, 95–104. [Google Scholar] [CrossRef]
- Leser, T.D.; Amenuvor, J.Z.; Jensen, T.K.; Lindecrona, R.H.; Boye, M.; Møller, K. Culture-independent analysis of gut bacteria: The pig gastrointestinal tract microbiota revisited. Appl. Environ. Microbiol. 2002, 68, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Combes, S.; Fortun-Lamothe, L.; Cauquil, L.; Gidenne, T. Engineering the rabbit digestive ecosystem to improve digestive health and efficacy. Animal 2013, 7, 1429–1439. [Google Scholar] [CrossRef]
- Combes, S.; Gidenne, T.; Cauquil, L.; Bouchez, O.; Fortun-Lamothe, L. Coprophagous behavior of rabbit pups affects implantation of cecal microbiota and health status. J. Anim. Sci. 2014, 92, 652–665. [Google Scholar] [CrossRef]
- Gidenne, T. Recent advances in rabbit nutrition: Emphasis on fibre requirements. A review. World Rabbit Sci. 2010, 8, 23–32. [Google Scholar]
- Carabaño, R.; Navarro, I.B.; Chamorro, S.; García, J.; Ruiz, A.G. New trends in rabbit feeding: Influence of nutrition on intestinal health. Span. J. Agric. Res. 2008, 1, 15–25. [Google Scholar] [CrossRef]
- Michelland, R.J.; Combes, S.; Monteils, V.; Cauquil, L.; Gidenne, T.; Fortun-Lamothe, L. Molecular analysis of the bacterial community in digestive tract of rabbit. Anaerobe 2010, 16, 61–65. [Google Scholar] [CrossRef]
- Combes, S.; Michelland, R.J.; Monteils, V.; Cauquil, L.; Soulié, V.; Tran, N.U.; Gidenne, T.; Fortun-Lamothe, L. Postnatal development of the rabbit caecal microbiota composition and activity. FEMS Microbiol. Ecol. 2011, 77, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Abecia, J.A.; Sosa, C.; Forcada, F.; Meikle, A. The effect of undernutrition on the establishment of pregnancy in the ewe. Reprod. Nutr. Dev. 2006, 46, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Gidenne, T.; Combes, S.; Fortun-Lamothe, L. Feed intake limitation strategies for the growing rabbit: Effect on feeding behaviour, welfare, performance, digestive physiology and health: A review. Animal 2012, 6, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Gallois, M.; Gidenne, T.; Fortun-Lamothe, L.; Le Huerou-Luron, I. An early stimulation of solid feed intake slightly influences the morphological gut maturation in the rabbit. Reprod. Nutr. Dev. 2005, 45, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Tumova, E.; Zita, L.; Skrivanova, V.; Fucikova, A.; Skrivan, M.; Buresova, M. Digestibility of nutrients, organ development and blood picture in restricted and ad libitum fed broiler rabbits. Archiv. Fur Geflugelkunde 2007, 71, 6–12. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 15th ed.; AOAC: Washington, DC, USA, 1995. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, rapidly, OTUs with galaxy solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and fast clustering method for amplicon-based studies. Peer J. 2014, 2, e386v381. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The silva ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- StataCorp. Stata Statistical Software: Release 14.0; StataCorp LP: College Station, TX, USA, 2015. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- RStudio Team. Rstudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Bagóné Vántus, V.; Kovács, M.; Zsolnai, A. The rabbit caecal microbiota: Development, composition and its role in the prevention of digestive diseases—A review on recent literature in the light of molecular genetic methods. Acta Agraria Kaposvan Riensis 2014, 18, 55–65. [Google Scholar]
- Castellini, C.; Cardinali, R.; Rebollar, P.G.; Dal Bosco, A.; Jimeno, V.; Cossu, M.E. Feeding fresh chicory (Chicoria intybus) to young rabbits: Performance, development of gastro-intestinal tract and immune functions of appendix and Peyer’s patch. Anim. Feed Sci. Technol. 2007, 134, 56–65. [Google Scholar] [CrossRef]
- Mousa, M.R.M. Effect of feeding acacia as supplements on the nutrient digestion, growth performance, carcass traits and some blood constituents of Awassi lambs under the conditions of North Sinai. Asian J. Anim. Sci. 2011, 5, 102–117. [Google Scholar] [CrossRef]
- Read, T.; Combes, S.; Gidenne, T.; Destombes, N.; Grenet, L.; Fortun-Lamothe, L. Stimulate feed intake before weaning and control intake after weaning to optimise health and growth performance. World Rabbit Sci. 2015, 23, 145–153. [Google Scholar] [CrossRef]
- Orengo, J.; Gidenne, T. Feeding behaviour and caecotrophy in the young rabbit before weaning: An approach by analysing the digestive contents. Appl. Anim. Behav. Sci. 2007, 102, 106–118. [Google Scholar] [CrossRef]
- Gidenne, T.; Garcia, J. Nutritional strategies improving the digestive health of the weaned rabbit. In Recent Advances in Rabbit Sciences; European cooperation in the field of Scientific and Technical Research (COST): Brussels, Belgium, 2006; p. 229. [Google Scholar]
- Gidenne, T.; Debray, L.; Fortun-Lamothe, L.; Le Huërou-Luron, I. Maturation of the intestinal digestion and of microbial activity in the young rabbit: Impact of the dietary fibre: Starch ratio. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 148, 834–844. [Google Scholar] [CrossRef]
- Xiccato, G.; Trocino, A.; Majolini, D.; Fragkiadakis, M.; Tazzoli, M. Effect of decreasing dietary protein level and replacing starch with soluble fibre on digestive physiology and performance of growing rabbits. Animal 2011, 5, 1179–1187. [Google Scholar] [CrossRef]
- Perez, J.M.; Gidenne, T.; Bouvarel, I.; Arveux, P.; Bourdillon, A.; Briens, C.; Mirabito, L. Replacement of digestible fibre by starch in the diet of the growing rabbit. II. Effects on performances and mortality by diarrhoea. Annales de Zootech 2000, 49, 369–377. [Google Scholar] [CrossRef]
- Gidenne, T.; Arveux, P.; Madec, O. The effect of the quality of dietary lignocellulose on digestion, zootechnical performance and health of the growing rabbit. Anim. Sci. 2001, 73, 97–104. [Google Scholar] [CrossRef]
- Maitre, I.; Lebas, F.; Arveux, P.; Bourdillon Supperay, J.; Saint Cast, Y. Taux de lignocellulose (ADF de Van Soest) et performances de croissance du lapin de chair. In Proceedings of the 5Èmes Journées de la Recherche Cunicoles, Paris, France, 12–13 December 1990; Volume 2, pp. 56.1–56.11. [Google Scholar]
- Gidenne, T. Caeco-colic digestion in the growing rabbit: Impact of nutritional factors and related disturbances. Livest. Prod. Sci. 1997, 51, 73–88. [Google Scholar] [CrossRef]
- Padilha, M.T.S.; Licois, D.; Gidenne, T.; Carrè, B.; Fonty, G. Relationships between microflora and caecal fermentation in rabbits before and after weaning. Reprod. Nutr. Dev. 1995, 35, 375–386. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, H.; Sun, D.; Liu, L.; Xu, X. Effects of doe-litter separation on intestinal bacteria, immune response and morphology of suckling rabbits. World Rabbit Sci. 2018, 26, 71–79. [Google Scholar] [CrossRef]
- Bäuerl, C.; Collado, M.C.; Zúñiga, M.; Blas, E.; Martínez, G.P. Changes in cecal microbiota and mucosal gene expression revealed new aspects of epizootic rabbit enteropathy. PLoS ONE 2014, 9, e105707. [Google Scholar] [CrossRef] [PubMed]
- Rhee, K.J.; Sethupathi, P.; Driks, A.; Lanning, D.K.; Knight, K.L. Role of commensal bacteria in development of gut-associated lymphoid tissues and preimmune antibody repertoire. J. Immunol. 2004, 172, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- Hanson, N.B.; Lanning, D.K. Microbial induction of B and T cell areas in rabbit appendix. Dev. Comp. Immunol. 2008, 32, 980–991. [Google Scholar] [CrossRef] [PubMed]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.D.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Koeck, D.E.; Pechtl, A.; Zverlov, V.V.; Schwarz, W.H. Genomics of cellulolytic bacteria. Curr. Opin. Biotechnol. 2014, 29, 171–183. [Google Scholar] [CrossRef]
- Fortun-Lamothe, L.; Boullier, S. A review on the interactions between gut microflora and digestive mucosal immunity. Possible ways to improve the health of rabbits. Livest. Sci. 2007, 107, 1–18. [Google Scholar] [CrossRef]
- Morrow, A.; Lagomarcino, A.; Schibler, K.; Taft, D.; Yu, Z.; Wang, B.; Altaye, M.; Wagner, M.; Gevers, D.; Ward, D.V.; et al. Early microbial and metabolomic signatures predict later onset of necrotizing enterocolitis in preterm infants. Microbiome 2013, 1, 13. [Google Scholar] [CrossRef]
- Comito, D.; Romano, C. Dysbiosis in the pathogenesis of pediatric inflammatory bowel diseases. Int. J. Inflamm. 2012, 2012, 687143. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximate Composition | Pelleted Feed | Dehydrated Alfalfa | Fresh Grass |
|---|---|---|---|
| Dry Matter | 89.00 | 88.00 | 35.61 |
| Crude protein | 20.45 | 13.01 | 11.57 |
| Ether extract | 4.63 | 2.09 | 2.50 |
| Ash | 11.57 | 8.06 | 25.02 |
| Crude fiber | 26.99 | 38.01 | 30.61 |
| Neutral detergent fiber (NDF) | 24.02 | 51.75 | 25.46 |
| Acid detergent fiber (ADF) | 14.12 | 38.72 | 15.77 |
| Acid digestible lignin (ADL) | 2.91 | 7.10 | 3.22 |
| Hemicellulose | 9.91 | 13.03 | 9.70 |
| Cellulose | 11.21 | 31.72 | 12.55 |
| Productive Performances | Unit | Experimental Groups 1 | Sig. 2 | ||
|---|---|---|---|---|---|
| C | A | G | |||
| Rabbit born per doe | n litters | 9 | 9 | 9 | - |
| Weaned rabbits (30 days) | n | 8 | 8 | 8 | ns |
| Rabbits at 45 days | “ | 8 | 8 | 8 | ns |
| Individual weight at birth | g | 53.10 ± 3.10 | 55.12 ± 2.34 | 51.34 ± 2.50 | ns |
| Individual weight at 15 days | “ | 240.0 ± 8.2 | 243.00 ± 5.78 | 239.30 ± 7.12 | ns |
| Individual weight at 30 days | “ | 489.0 ± 121.5 | 500.50 ± 136.59 | 485.50 ± 74.23 | ns |
| Individual weight at 45 days | “ | 1058.00 ± 83.104 | 1050.44 ± 138.17 | 1019.1 ± 145.71 | ns |
| Daily weight gain 0–15 days | g/day/rabbit | 12.46 ± 3.12 | 12.53 ± 2.87 | 12.53 ± 1.16 | ns |
| Daily weight gain 15–30 days | “ | 16.6 ± 5.32 | 17.13 ± 6.87 | 16.4 ± 8.81 | ns |
| Daily weight gain 30–45 days | “ | 37.93 ± 10.22 | 36.66 ± 6.66 | 35.60 ± 4.65 | ns |
| Pre-weaning mortality (0–30 days) | % | 12.5 | 12.5 | 12.5 | ns |
| Milk production 0–15 days | g/day/doe | 148.02 ± 11.83 | 143.34 ± 10.54 | 145.35 ± 11.86 | ns |
| Milk intake 0–15 days | g/day/rabbit | 18.50 ± 2.16 | 17.91 ± 3.54 | 17.79 ± 1.86 | ns |
| Pelleted feed intake 15–30 days | “ | 11.55 ± 1.02 a | 6.56 ± 0.53 b | 7.43 ± 0.65 b | * |
| Supplemental intake 15–30 days | “ | - | 10.18 ± 4.25 b | 16.72 ± 3.99 a | * |
| Total feed intake (15–30 days) | “ | 11.55 ± 1.02 c | 16.74 ± 4.18 b | 24.15 ± 3.87 a | * |
| NDF intake (15–30 days) | “ | 2.47 ± 0.33 c | 6.04 ± 0.32 a | 3.09 ± 0.14 b | * |
| ADF intake (15–30 days) | “ | 1.45 ± 0.21 c | 4.29 ± 0.26 a | 2.38 ± 0.13 b | * |
| ADL intake (15–30 days) | “ | 0.29 ± 0.04 c | 1.05 ± 0.04 a | 0.38 ± 0.01 b | * |
| Cellulose intake (15–30 days) | “ | 1.15 ± 0.12 c | 3.49 ± 0.19 a | 1.83 ± 0.06 b | * |
| Pelleted feed intake 30–45 days | “ | 150.81 ± 56.28 a | 143.26 ± 57.60 b | 146.45 ± 32.62 ab | * |
| Supplemental intake 30–45 days | “ | - | 11.56 ± 3.68 b | 25.12 ± 2.55 a | * |
| Feed efficiency | 3.98 | 3.90 | 4.10 | ns | |
| NDF intake (30–45 days) | g/day/rabbit | 32.24 ± 2.15 b | 35.92 ± 2.89 a | 33.61 ± 2.84 b | * |
| ADF intake (30–45 days) | “ | 18.95 ± 1.08 c | 21.96 ± 1.31 a | 19.83 ± 1.07 b | * |
| ADL intake (30–45 days) | “ | 3.90 ± 0.34 b | 4.43 ± 0.40 a | 4.08 ± 0.01 b | * |
| Cellulose intake (30–45 days) | “ | 15.04 ± 1.00 b | 17.53 ± 0.97 a | 15.75 ± 0.98 b | * |
| Phyla | Experimental Groups 1 | CV 2 | p3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 30 Days | 45 Days | ||||||||
| C | A | G | C | A | G | Age | Feed | ||
| Firmicutes | 65.78 c | 72.07 ab | 62.38 c | 78.87 b | 83.64 a | 79.98 b | 6.65 | *** | * |
| Bacteroidetes | 28.15 a | 24.83 a | 30.41 a | 14.15 b | 12.75 b | 13.26 b | 4.32 | *** | ns |
| Proteobacteria | 4.50 a | 1.59 b | 1.92 b | 3.02 ab | 2.30 ab | 3.48 ab | 1.04 | * | * |
| Tenericutes | 0.76 b | 1.19 b | 4.22 a | 3.02 a | 1.10 b | 2.28 ab | 0.82 | * | ns |
| Actinobacteria | 0.78 ab | 0.29 b | 1.05 a | 0.85 ab | 0.17 b | 0.96 ab | 0.63 | * | ns |
| Spirochaetae | 0.03 | 0.03 | 0.02 | 0.09 | 0.04 | 0.04 | 0.05 | ns | ns |
| Firmicutes/Bacteroidetes | 2.34 d | 2.90 c | 2.05 d | 5.57 b | 6.56 a | 6.03 ab | 1.02 | ** | * |
| Ruminococcaceae/Lachnospiraceae | <0.001 d | 1.58 b | 0.03 d | 0.77 c | 24.32 a | 0.07 d | 3.12 | ** | * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattioli, S.; Dal Bosco, A.; Combes, S.; Moscati, L.; Crotti, S.; Cartoni Mancinelli, A.; Cotozzolo, E.; Castellini, C. Dehydrated Alfalfa and Fresh Grass Supply in Young Rabbits: Effect on Performance and Caecal Microbiota Biodiversity. Animals 2019, 9, 341. https://doi.org/10.3390/ani9060341
Mattioli S, Dal Bosco A, Combes S, Moscati L, Crotti S, Cartoni Mancinelli A, Cotozzolo E, Castellini C. Dehydrated Alfalfa and Fresh Grass Supply in Young Rabbits: Effect on Performance and Caecal Microbiota Biodiversity. Animals. 2019; 9(6):341. https://doi.org/10.3390/ani9060341
Chicago/Turabian StyleMattioli, Simona, Alessandro Dal Bosco, Sylvie Combes, Livia Moscati, Silvia Crotti, Alice Cartoni Mancinelli, Elisa Cotozzolo, and Cesare Castellini. 2019. "Dehydrated Alfalfa and Fresh Grass Supply in Young Rabbits: Effect on Performance and Caecal Microbiota Biodiversity" Animals 9, no. 6: 341. https://doi.org/10.3390/ani9060341
APA StyleMattioli, S., Dal Bosco, A., Combes, S., Moscati, L., Crotti, S., Cartoni Mancinelli, A., Cotozzolo, E., & Castellini, C. (2019). Dehydrated Alfalfa and Fresh Grass Supply in Young Rabbits: Effect on Performance and Caecal Microbiota Biodiversity. Animals, 9(6), 341. https://doi.org/10.3390/ani9060341