Simple Summary
Serum biochemical parameters and hormones play a role in directly reflecting the physiological state and modulating the milk performance of dairy cows. However, the variability of the serum biochemical parameters and hormones in multiparous mid-lactation cows have not drawn much attention due to their supposedly stable states, especially for those cows under the same nutrition and management condition. The aim of this study was to evaluate the effects of days-in-milk (DIM, within the mid-lactation) and parity (ranging from 2–6) on serum biochemical parameters and hormone profiles based on a large cohort of dairy cows. The results showed that DIM and parity contribute to the variations in serum biochemical parameters and hormones related to protein status, energy supply, liver and kidney function, and oxidative stress of mid-lactation dairy cows, with the effect of DIM being dominant over parity. Our result suggested that the DIM periods and parity should be taken into consideration to optimize nutritional strategies in order to improve the milk performance traits more precisely.
Abstract
It is well known that serum biochemical parameters and hormones contribute greatly to the physiological and metabolic status of dairy cows. However, few studies have focused on the variation of these serum parameters in multiparous mid-lactation cows without the interference of diet and management. A total of 287 Holstein dairy cows fed the same diet and maintained under the same management regime were selected from a commercial dairy farm to evaluate the effects of days-in-milk (DIM) and parity on serum biochemical parameters and hormone profiles. Milk yield and milk protein content were affected by DIM and parity (p < 0.05). Milk protein yield showed a numerically decreasing trend with parity, and it was relatively constant in cows with parities between 2 and 4 but lower in cows with parity 6 (p = 0.020). Ten and five serum biochemical parameters related to protein status, energy metabolism, liver and kidney function, and oxidative stress were affected by DIM and parity, respectively (p < 0.05). Glucagon, insulin-like growth factor 1 concentration, and the revised quantitative insulin sensitivity check index were significantly different (p < 0.05) among cows with different DIM. Parity had no effect on hormone concentrations. An interaction between DIM and parity effect was only detected for glucagon concentration (p = 0.015), which showed a significantly increasing trend with DIM and overall decreasing trend with parity. In summary, DIM and parity played an important role in affecting the serum biochemical parameters and/or hormones of dairy cows, with serum parameters affected more by DIM than parity.
1. Introduction
Serum biochemical parameters and hormones directly reflect the physiological state of dairy cows [1] and may play a role in modulating milk production [2]. However, most studies focusing on factors affecting milk production have been based on differences in nutrition [2], management [3], or health [4], and only used a small number of cows. Recent studies have reported that healthy cows can have different milk production levels even when they are fed the same diet and managed under the same conditions [5]. However, it is unknown to what extent the serum biochemical parameters and hormones differ among these cows, and establishing further knowledge on this subject may help to evaluate the physiological state of dairy cows and to clarify the physiological regulation for variant milk performance traits.
The variations in serum biochemical parameters and hormones under the peripartum, early-lactation, and peak-lactation period have been investigated for years [1,3]. However, the variability of those parameters in mid-lactation cows has not drawn much attention due to their relatively stable physiological states in comparison to the other lactation stages. On the other hand, studies on parity effects have mainly focused on variation analysis between primiparous and multiparous cows [6]. Xue et al. (2018) reported that days-in-milk (DIM) and parity (ranging from 2–7) significantly affected the relative abundance of rumen bacteria in multiparous mid-lactation cows [7]. It is well documented that rumen microbial fermentation products can affect the concentration of metabolites in the blood such as blood urea nitrogen (BUN), glucose, and ketone [8]. Therefore, we hypothesized that DIM and parity could also affect serum biochemical parameters in mid-lactation cows. Serum hormones are usually used to reflect the real-time physiological status of dairy cows [1], so the variation analysis of hormone profiles under different DIM periods or parities can allow physiological shifts to be identified with significantly more precision.
The objective of this study was to investigate the effects of the DIM and parity on the serum biochemical parameters and hormone profiles of Holstein dairy cows under the same diet and management conditions.
2. Materials and Methods
2.1. Animals and Management
All experimental designs and protocols were approved by the Animal Care Committee (ZJU20170422) at Zhejiang University (Hangzhou, P. R. China), and the study was performed following the university’s guidelines (IDS1703) for animal research.
A total of 287 healthy Holstein dairy cows with DIM of 156 (± 33.5 SD) and parity of 3.03 (± 1.12 SD) were selected from the Hangjiang Dairy Farm (Hangzhou, China). Feeding and management of the dairy cows have been described previously [9]. Briefly, the cows were kept individually in well-ventilated tethered stalls in a barn, and had free access to drinking water. They were fed total mixed ration [concentrate: forage = 57: 43, Dry matter (DM) basis] and milked three times per day at 06:30, 14:00, and 20:00 h.
2.2. Sample Collection and Measurement
The diet ingredients and chemical compositions were collected and measured and are shown in Supplementary Table S1. Milk yields were recorded and milk samples were sampled to determine milk composition. Blood samples were collected using pro-coagulation tubes before morning feeding (04:00–06:00), and serum was obtained for analysis of serum biochemical parameters. The details for collection and treatment of the samples were described previously [9]. The determined serum biochemical parameters include BUN, total protein, albumin, glucose, non-esterified fatty acids (NEFA), total cholesterol, β-hydroxybutyrate (BHB), triglyceride, creatinine, total bilirubin, malonaldehyde (MDA), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), and total antioxidant capability (T-AOC).
A total of 500 μL of serum was subjected to hormone profiling using colorimetric commercial kits (Jiangsu Meibiao Biological Technology Co. Ltd., Yancheng, China) in a DG5033A microplate reader (Nanjing Huadong Electronics Group Medical Equipment Co. Ltd., Nanjing, China) according to the manufacturer’s protocol. The determined hormones include insulin, glucagon, insulin-like growth factor 1 (IGF-1), cortisol, ghrelin, and leptin. The revised quantitative insulin sensitivity check index (RQUICKI) was calculated using the method reported by Holtenius and Holtenius (2007) [10].
2.3. Grouping Based on Days-in-Milk and Parity
Experimental cows were divided into four groups based on their DIM: 90–119, 120–149, 150–179, and 180–219, as reported in Xue et al. (2018) [7]. To explore the parity effects, experimental cows were separated into five groups based on the parity range (parity: 2–6). All of the ‘groups’ of cows were housed individually in the same pen to avoid the interference of environment and management.
2.4. Statistical Analysis
Statistical analysis was performed using SAS 9.2 (SAS Institute Inc., Cary, NC, USA). Notably, variable outliers were identified as those beyond 3 standard deviations of the mean and were discarded [11]. The 95% confidence interval (CI) was calculated based on the SAS PROC MEANS model. The PROC UNIVARIATE model and PROC DISCRIM model were used to conduct the normality and homogeneity test, respectively. One-way ANOVA or a non-parametric test was utilized to analyze the DIM and parity contributions based on the normality and homogeneity test results. Bonferroni correction was utilized to correct the p values of multiple testing among different DIM or parity groups. Two-way ANOVA was performed to assess the interaction of DIM and parity effects. Statistical significance of the main effects was declared at p < 0.05, and tendencies were accepted at 0.05 < p < 0.10.
3. Results and Discussion
3.1. General Information
In this study, high individual variations existed in milk yield (coefficient of variation, CV = 17.1%) and milk protein yield (CV = 15.6%). The BUN, glucose, NEFA, total cholesterol, BHB, triglyceride, creatinine, total bilirubin, MDA, and T-AOC also had high variability (CV > 15%). All hormones except for RQUICKI showed a high individual variation (CV > 15%). Notably, although high variability existed in the above parameters, all variations are within the range of homeostatic status as Cozzi et al. (2011) reported [11].
The serum biochemical parameters measured from 287 cows could serve as reference information in order to evaluate the metabolic status of multiparous mid-lactation Holstein cows. Although researchers have reported references regarding ruminant serum biochemical parameters, the information was too broad to distinguish the difference between dairy cows and beef cattle [12]. For dairy cows, the available references of blood biochemical parameters were only based on a relatively small cohort (n = 20) of dairy cows [13]. Thus, the reference intervals of milk performance and serum biochemical parameters based on the 287 lactating dairy cows are of significant value to provide guidance for metabolic evaluation and nutrition adjustment in the dairy industry.
3.2. Effects of Days-in-Milk on Serum Biochemical Parameters and Hormones
The DIM exerted significant effects on milk yield, milk protein content, milk urea nitrogen content (MUN), and somatic cell counts (SCC) (p < 0.001, Table 1). It has been well established that milk performance varies through the different lactation stages [5]. However, there is no well-described research on how milk performance traits vary within the whole mid-lactation period, which was covered in this study. No significant difference in parity existed in the four DIM groups.

Table 1.
Effects of days-in-milk on milk performance and blood biochemical parameters.
Concentrations of 10 serum biochemical parameters were significantly different among cows with varied DIM, including the BUN, total protein, glucose, NEFA, BHB, creatinine, total bilirubin, MDA, GSH-Px, and T-AOC (p < 0.05, Table 1). As reported by Puppel and Kuczyńska (2016) [14], the changes in BUN, total protein, creatinine, and total bilirubin can be used to evaluate the protein status of dairy cows. The milk protein yield of dairy cows did not change with different DIM (p = 0.351), but milk protein content tended to increase with DIM (p < 0.001). The higher BUN in the DIM 150–179 and DIM 180–219 groups (4.33 and 4.47 g/L, respectively) compared with the DIM 90–119 and DIM 120–149 groups (3.97 and 3.70 g/L, respectively; p < 0.01) may imply that more ammonia was metabolized to urea in the liver at these later DIM stages. Additionally, the lower total protein concentration in the DIM 150–179 and DIM 180–219 groups (62.7 and 59.2 g/L, respectively) than in the DIM 90–119 and DIM 120–149 groups (66.6 and 66.7 g/L, respectively; p < 0.01) indicated undesired protein efficiency in dairy cows at the later stage. The overall increasing glucose (p = 0.046) and NEFA (p = 0.005) concentrations with DIM, and the high BHB concentration in the DIM 90–119 group compared with those in DIM 150–179 and DIM 180–219 groups (p = 0.008) reflected the energy status changes in the mid-lactation dairy cows. As an indicator of energy balance, the changes in BHB concentration under different DIM periods align with the milk yield changes, as reported by Sakowski et al. (2012) [15]. Furthermore, the BHB has been reported to upregulate the expression of milk protein synthesis genes in bovine mammary epithelial cells [16]. However, the changes in BHB in this study were opposite to the milk protein variations in the different DIM groups. It warrants further study how the BHB is involved in milk protein synthesis. The dynamic changes in MDA concentration, GSH-Px, and T-AOC activity during the mid-lactation period indicated a fluctuation in oxidative stress related to milk synthesis. In the current study, MDA concentration was significantly higher in the first two DIM groups than that in the latter two groups (5.00, 4.72, 3.29, and 3.64 nmol/L, respectively; p < 0.001), which was partly consistent with the milk yield changes.
The effects of DIM on hormone profiles are shown in Figure 1a. First, glucagon concentration was significantly lower in the DIM 90–119 group than the DIM 120–149 group (88.2, 98.0, 95.2, and 95.2 pg/mL, respectively; p = 0.012), which is partly consistent with the change in milk protein content. However, Bobe et al. (2009) found that glucagon infusions decreased milk protein content and yield [17]. The inconsistent results may be derived from different glucagon doses and variant lactation stages, as Hippen et al. (1999) reported [18]. Moreover, both the IGF-1 concentration (p = 0.001) and RQUICKI (p < 0.001) showed a curvilinear change with increasing DIM. IGF-1 was reported to increase milk yield or milk protein synthesis in in vitro studies [19,20]. However, no corresponding result was found between IGF-1 and milk yield or milk protein content in this study. In our current study, RQUICKI was affected by the DIM periods, as previously reported [21]. The relatively higher RQUICKI in the DIM 90–179 group indicated higher insulin sensitivity to promote the glucose utilization for lactation and maintenance in earlier mid-lactation, which was proved by the relatively lower glucose concentration and higher milk yield in those DIM periods.
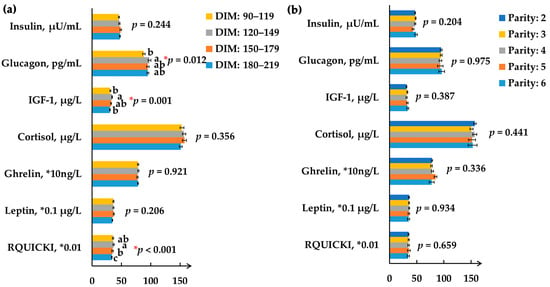
Figure 1.
Effect of days-in-milk (a) and parity (b) on serum hormone profiles. (a) Description of days-in-milk effects on serum hormone profiles; (b) Description of parity effects on serum hormone profiles. Different color bars represent varying days-in-milk or different parities. Bar height represents the mean value of each individual hormone; the error bars represent the standard error of mean. IGF-1: insulin-like growth factor 1; RQUICKI: revised quantitative insulin sensitivity check index [10]. a,b Means within a hormone with different superscripts significantly differ (p < 0.05).
3.3. Effects of Parity on Serum Biochemical Parameters and Hormones
Milk yield, milk protein content and yield, MUN, and SCC were affected by parity (p < 0.05, Table 2). Similarly, research related to the effects of parity on milk performance traits has been studied for years [5], and was not the focus of this research. No significant difference existed in DIM within any parity group (p > 0.05).
Table 2.
Effects of parity on milk performance and blood biochemical parameters.
In the current study, the concentrations of five serum biochemical parameters were affected by parity (p < 0.05, Table 2). Serum total protein concentration was higher in dairy cows with parity 5 than those with parity 2 (p = 0.005, Table 2), which aligns with the findings of Blum et al. (1983) [22]. The serum total cholesterol concentration was higher in cows with parity 2 than those with parity 3 and parity 6 (6.35, 5.86, 5.87, 5.81, and 5.24 mmol/L for parities 2 to 6, respectively; p = 0.001), which might reveal more lipid mobilization to provide energy for more milk production in parity-2 cows. Serum BHB concentration was relatively stable in cows with parities 2 to 5, but lower in parity-6 cows (0.50, 0.51, 0.49, 0.47, and 0.40 mmol/L for parities 2 to 6, respectively; p = 0.035). Under different parities, the consistent changes in the BHB concentration and milk yield revealed the significance of energy supply for milk synthesis. Creatinine concentration in the serum was highest in dairy cows with parity 2 and decreased until parity 5 (62.5, 59.1, 57.2, 54.3, and 58.5 μmol/L for parities 2 to 6, respectively; p = 0.004). The SOD concentration declined with increasing parity (p = 0.034). For all serum biochemical parameters, no interaction between DIM and parity effects was observed (p > 0.05).
All hormones that we detected were kept stable under the different parities (p > 0.05, Figure 1b). However, an interaction effect of DIM and parity on glucagon concentration (p = 0.015) was detected. No parity effect on the ghrelin level is in accordance with Honig et al. (2016), [23], which reflects that feed intake was enough to provide energy for lactation, and energy homeostasis was not the primary cause for variant milk performance traits of these experimental cows under different parity groups. The above results suggest that the hormones involved in energy and appetite are not the critical endocrine mediators to the observed variations in lactation performance with parity.
4. Conclusions
In conclusion, DIM and parity contribute to the variations in serum biochemical parameters and hormones related to protein status, energy supply, liver and kidney function, and oxidative stress of mid-lactation Holstein dairy cows, with the effect of DIM being dominant over parity. For multiparous mid-lactation cows, DIM and parity should also be considered to assist in optimizing the nutrition and management strategies according to their physiological metabolism.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-2615/9/5/230/s1, Table S1: Ingredients and nutrient compositions of the experimental diet.
Author Contributions
Conceptualization: J.L. and L.G.; methodology: X.W., M.X. and J.L.; formal analysis: X.W. and D.W.; data curation: X.W. and H.-Z.S.; writing—original draft preparation: X.W.; writing—review and editing: X.W., J.L., L.G., H.-Z.S., M.X. and D.W.; visualization: X.W.; supervision: J.L., L.G. and H.-Z.S.; funding acquisition: J.L. and L.G. All authors approved the manuscript.
Funding
This research was supported by grants from the National Natural Science Foundation of China (No. 31729004 to L.G. and J.L.) and the China Dairy Research System (CARS-36 to J.L.).
Acknowledgments
The authors thank the members of the Institute of Dairy Science at Zhejiang University (Hangzhou, China) for their assistance in the sampling.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Nowroozi-Asl, A.; Aarabi, N.; Rowshan-Ghasrodashti, A. Ghrelin and its correlation with leptin, energy related metabolites and thyroidal hormones in dairy cows in transitional period. Pol. J. Vet. Sci. 2016, 19, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Huhtanen, P.; Vanhatalo, A.; Varvikko, T. Effects of abomasal infusions of histidine, glucose, and leucine on milk production and plasma metabolites of dairy cows fed grass silage diets. J. Dairy Sci. 2002, 85, 204–216. [Google Scholar] [CrossRef]
- Wright, J.B.; Wall, E.H.; Mcfadden, T.B. Effects of increased milking frequency during early lactation on milk yield and udder health of primiparous Holstein heifers. J. Anim. Sci. 2013, 91, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Su, D.; Tian, H.; Li, X.; Li, Y.; Ran, L.; Hu, R.; Cheng, J. Liver Metabolic Perturbations of Heat-Stressed Lactating Dairy Cows. Asian Austral J. Anim. 2018, 31, 1244–1251. [Google Scholar] [CrossRef]
- Kim, T.I.; Mayakrishnan, V.; Park, J.H.; Ki, K.S.; Lim, D.H.; Kim, S.B.; Park, S.M.; Jeong, H.Y.; Park, B.Y. The effect of lactation number, stage, length, and milking frequency on milk yield in Korean Holstein dairy cows using automatic milking system. Asian Austral J. Anim. 2017, 30. [Google Scholar] [CrossRef]
- Bühler, S.; Frahm, J.; Tienken, R.; Kersten, S.; Meyer, U.; Huber, K.; Dänicke, S. Effects of energy supply and nicotinic acid supplementation on serum anti-oxidative capacity and on expression of oxidative stress-related genes in blood leucocytes of periparturient primi- and pluriparous dairy cows. J. Anim. Physiol. Anim. Nutr. 2018, 102, e87–e98. [Google Scholar] [CrossRef]
- Xue, M.; Sun, H.; Wu, X.; Guan, L.L.; Liu, J. Assessment of rumen microbiota from a large cattle cohort reveals the pan and core bacteriome contributing to varied phenotypes. Appl. Environ. Microbiol. 2018. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Penner, G.B.; Stumpff, F.; Gabel, G. RUMINANT NUTRITION SYMPOSIUM: Role of fermentation acid absorption in the regulation of ruminal pH. J. Anim. Sci. 2011, 89, 1092–1107. [Google Scholar] [CrossRef]
- Wu, X.; Sun, H.; Xue, M.; Wang, D.; Guan, L.L.; Liu, J. Serum metabolome profiling revealed potential biomarkers for milk protein yield in dairy cows. J. Proteom. 2018, 184, 54–61. [Google Scholar] [CrossRef]
- Holtenius, P.; Holtenius, K. A model to estimate insulin sensitivity in dairy cows. Acta Vet. Scand. 2007, 49, 1–3. [Google Scholar] [CrossRef]
- Cozzi, G.; Ravarotto, L.; Gottardo, F.; Stefani, A.L.; Contiero, B.; Moro, L.; Brscic, M.; Dalvit, P. Short communication: Reference values for blood parameters in Holstein dairy cows: Effects of parity, stage of lactation, and season of production. J. Dairy Sci. 2011, 94, 3895–3901. [Google Scholar] [CrossRef]
- Russell, K.E.; Roussel, A.J. Evaluation of the Ruminant Serum Chemistry Profile. Vet. Clin. N. Am. Food Anim. Pract. 2007, 23, 403–426. [Google Scholar] [CrossRef]
- Moore, F. Serum chemistry profiles in dairy cows—A herd management tool? Vet. Med. 1997, 92, 986–991. [Google Scholar]
- Puppel, K.; Kuczyńska, B. Metabolic profiles of cow’s blood; a review. J. Sci. Food Agric. 2016, 96, 4321–4328. [Google Scholar] [CrossRef]
- Sakowski, T.; Kuczyńska, B.; Puppel, K.; Metera, E.; Słoniewski, K.; Barszczewski, J. Relationships between physiological indicators in blood, and their yield, as well as chemical composition of milk obtained from organic dairy cows. J. Sci. Food Agric. 2012, 92, 2905. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, S.; Qi, L.; Sheng, R.; Zhao, Y. Effects of beta-Hydroxybutyric Acid on Expressions of Genes Involved in Milk Protein Synthesis in Bovine Mammary Epithelial Cells. Chin. J. Anim. Nutr. 2014, 26, 3836–3842. [Google Scholar] [CrossRef]
- Bobe, G.; Hippen, A.R.; She, P.; Lindberg, G.L.; Young, J.W.; Beitz, D.C. Effects of glucagon infusions on protein and amino acid composition of milk from dairy cows 1. J. Dairy Sci. 2009, 92, 130–138. [Google Scholar] [CrossRef]
- Hippen, A.R.; She, P.; Young, J.W.; Beitz, D.C.; Lindberg, G.L.; Richardson, L.F.; Tucker, R.W. Metabolic responses of dairy cows and heifers to various intravenous dosages of glucagon. J. Dairy Sci. 1999, 82, 1128–1138. [Google Scholar] [CrossRef]
- Prosser, C.G.; Davis, S.R.; Farr, V.C.; Moore, L.G.; Gluckman, P.D. Effects of close-arterial (external pudic) infusion of insulin-like growth factor-II on milk yield and mammary blood flow in lactating goats. J. Endocrinol. 1994, 142, 93–99. [Google Scholar] [CrossRef]
- Burgos, S.A.; Cant, J.P. IGF-1 stimulates protein synthesis by enhanced signaling through mTORC1 in bovine mammary epithelial cells. Domest. Anim. Endocrinol. 2010, 38, 211–221. [Google Scholar] [CrossRef]
- Kerestes, M.; Faigl, V.; Kulcsár, M.; Balogh, O.; Földi, J.; Fébel, H.; Chilliard, Y.; Huszenicza, G. Periparturient insulin secretion and whole-body insulin responsiveness in dairy cows showing various forms of ketone pattern with or without puerperal metritis. Domest. Anim. Endocrinol. 2009, 37, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.W.; Kunz, P.L.; Leuenberger, H.; Gautschi, K.; Keller, M. Thyroid hormones, blood plasma metabolites and hematological parameters in relationship to milk yield in dairy cows. Anim. Prod. 1983, 36, 93–104. [Google Scholar] [CrossRef]
- Honig, H.; Ofer, L.; Elbaz, M.; Kaim, M.; Shinder, D.; Gershon, E. Seasonal and parity effects on ghrelin levels throughout the estrous cycle in dairy cows. Gen. Comp. Endocrinol. 2016, 235, 64–69. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).