Modelling Nutritional Requirements of Growing Pigs from Local Breeds Using InraPorc

 ,
,  , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Requirements for Modelling Study
2.2. Data Collection
2.2.1. Alentejana
2.2.2. Basque
2.2.3. Bísara
2.2.4. Apulo Calabrese
2.2.5. Cinta Senese
2.2.6. Iberian
2.2.7. Krškopolje Pig
2.2.8. Mangalitsa and Moravka
2.3. Growth Modelling
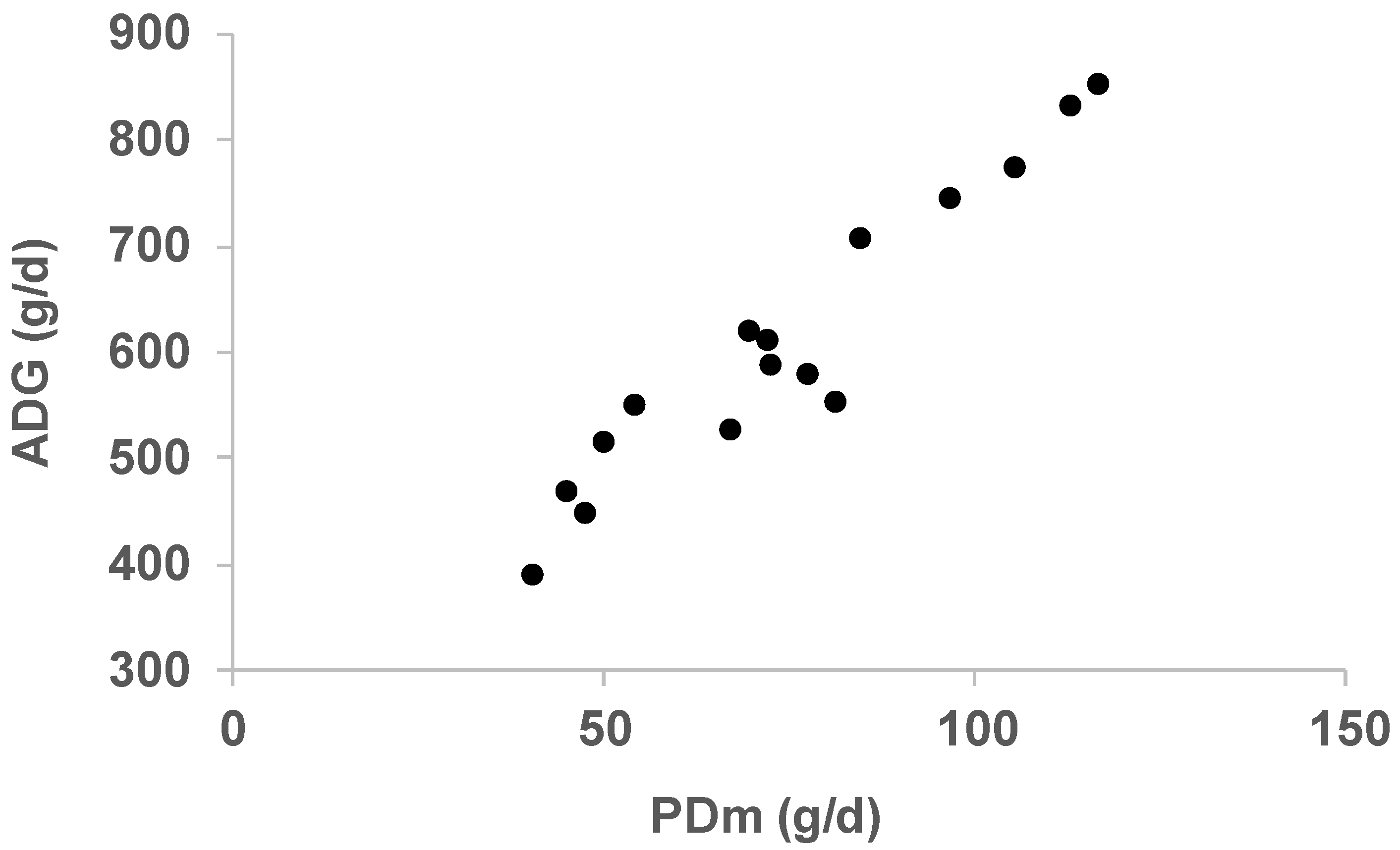
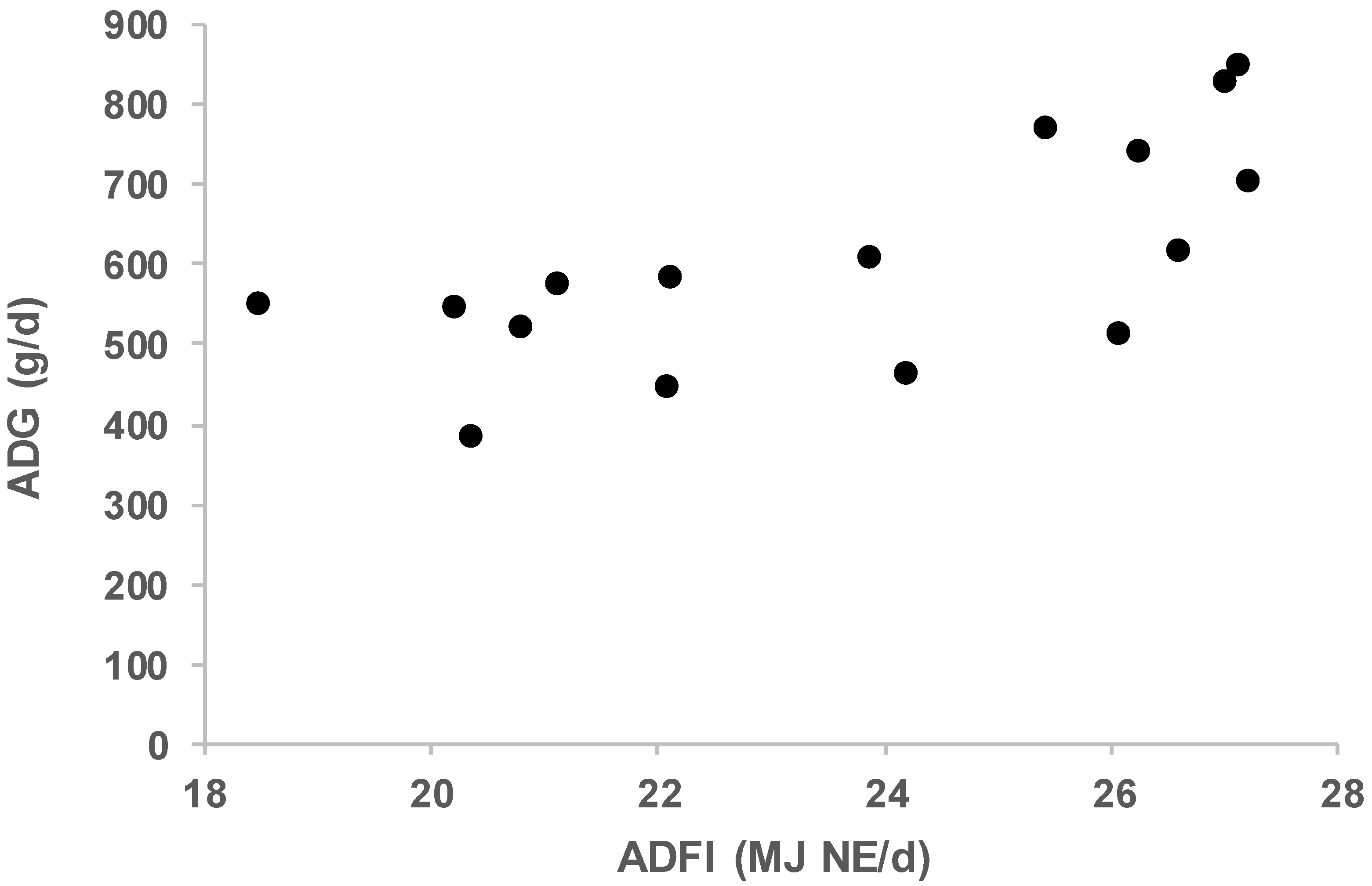
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial 1 | Trial 2 | ||||||
|---|---|---|---|---|---|---|---|
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d | Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d |
| 41.5–54.2 | 22 | 28 | 2.50 | 35.3–45.8 | 39 | 28 | 1.83 |
| 54.2–69.6 | 22 | 28 | 3.15 | 45.8–59.7 | 39 | 30 | 2.08 |
| 69.6–83.1 | 22 | 28 | 3.58 | 59.2–95.1 | 13 | 78 | 2.82 |
| 83.1–96.3 | 22 | 34 | 4.68 | ||||
| Group—Conventional | Group—Alternative | ||||||
|---|---|---|---|---|---|---|---|
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d | Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d |
| 34.8–74.7 | 20 | 64 | 2.23 | 35.3–75.4 | 20 | 60 | 2.43 |
| 74.7–109.1 | 20 | 70 | 2.51 | 75.4–111.5 | 20 | 70 | 2.64 |
| 109.1–139.9 | 20 | 80 | 2.41 | 111.5–146.3 | 20 | 76 | 2.85 |
| [12] 1 | Project TREASURE 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d | Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d | |
| Feed | Corn | |||||||
| 29.8–95.8 | 24 | 118 | 2.052 | 25.3–34.3 | 10 | 14 | 1.5 | 0 |
| 95.8–144.3 | 24 | 91 | 2.618 | 34.3–42.9 | 10 | 14 | 1.5 | 0 |
| 42.9–52.4 | 10 | 14 | 1.5 | 0.3 | ||||
| 52.4–59.4 | 10 | 14 | 1.5 | 0.3 | ||||
| 59.4–68.8 | 10 | 14 | 1.5 | 0.4 | ||||
| 68.8–74.6 | 10 | 14 | 1.5 | 0.4 | ||||
| 74.6–80.5 | 10 | 14 | 1.5 | 0.4 | ||||
| 80.5-–90.1 | 10 | 14 | 1.5 | 0.6 | ||||
| 90.1–96.7 | 10 | 14 | 1.5 | 0.6 | ||||
| 96.7–103.4 | 10 | 14 | 1.5 | 0.6 | ||||
| 103.4–110.6 | 10 | 14 | 1.5 | 0.6 | ||||
| 110.6–115.8 | 10 | 14 | 1.5 | 0.6 | ||||
| Group without soy 2 | Group with soy 3 | ||||||
|---|---|---|---|---|---|---|---|
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d | Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d |
| 40.2–72.2 | 36 | 42 | 2.3 | 33.8–62.7 | 36 | 42 | 2.1 |
| 72.2–115.2 | 36 | 56 | 3.3 | 62.7–105.4 | 36 | 56 | 3.3 |
| 115.2–139.1 | 36 | 44 | 3.7 | 105.4–35.2 | 36 | 44 | 3.5 |
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d |
|---|---|---|---|
| 38.0–46.45 | 11 | 29 | 1.23 |
| 46.45–57.95 | 11 | 56 | 1.82 |
| 57.95–67.77 | 11 | 84 | 2.00 |
| 67.77–85.27 | 11 | 119 | 2.40 |
| 85.27–95.45 | 11 | 149 | 2.65 |
| 95.45–106.23 | 11 | 183 | 2.80 |
| 106.23–123.9 | 11 | 212 | 3.37 |
| 123.9–143.35 | 11 | 246 | 3.32 |
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d |
|---|---|---|---|
| 16.1–35.9 | 29 | 68 | 0.91 |
| 35.9–103.2 | 20 | 135 | 2.08 |
| 103.2–159.5 | 20 | 70 | 3.28 |
| Reduced Protein Diet in Growing Phase 1 | Usual Protein Diet in Growing Phase 2 | ||||||
|---|---|---|---|---|---|---|---|
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d | Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d |
| 15.7–50.8 | 8 | 68.3 | 1.78 | 16.0–48.5 | 8 | 67.4 | 1.76 |
| 51.9–114.0 | 6 | 86.8 | 3.13 | 48.1–114.0 | 6 | 107 | 3.05 |
| Conventional Rearing System 2 Diet 1 (8.52 MJ NE/kg, 15.3% Protein) Diet 2 (8.82 MJ NE/kg, 14.0% Protein) | Ecological Rearing System 3 Diet 1 (8.69 MJ NE/kg, 15.0% Protein) Diet 2 (8.83 MJ NE/kg, 14.0% Protein) | ||||||
|---|---|---|---|---|---|---|---|
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d | Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d |
| 36.2–45.4 | 12 | 14 | 1.88 | 35.0–43.7 | 12 | 14 | 2.22 |
| 45.4–56.6 | 12 | 14 | 2.63 | 43.7–54.1 | 12 | 14 | 2.64 |
| 56.6–69.4 | 12 | 14 | 2.99 | 54.1–66.4 | 12 | 14 | 2.98 |
| 69.4–83.2 | 12 | 14 | 3.56 | 66.4–79.2 | 12 | 14 | 3.54 |
| 83.2–94.3 | 12 | 14 | 3.26 | 79.2–91.7 | 12 | 14 | 3.26 |
| 94.3–107.3 | 12 | 13 | 3.26 | 91.7–103.5 | 12 | 13 | 3.26 |
| 107.3–111.8 | 12 | 6 | 3.26 | 103.5–115.4 | 12 | 6 | 3.35 |
| 111.8–116.4 | 12 | 12 | 3.45 | 115.4–117.5 | 12 | 12 | 3.35 |
| 116.4–120.4 | 12 | 14 | 3.45 | 117.5–124.3 | 12 | 14 | 3.35 |
| Mangalitsa | Moravka | ||||||
|---|---|---|---|---|---|---|---|
| Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d | Weight, kg | No. of Pigs | Days on Trial | Feed Intake kg/Pig/d |
| 24.3–35.9 | 12 | 35 | 1.43 | 29.9–46.2 | 10 | 35 | 1.61 |
| 35.9–44.8 | 12 | 19 | 1.97 | 46.2–55.6 | 10 | 19 | 2.24 |
| 44.8–59.2 | 12 | 28 | 2.5 | 55.6–71.5 | 10 | 28 | 2.74 |
| 59.2–79.2 | 12 | 29 | 2.67 | 71.5–89.1 | 10 | 29 | 3.10 |
| 79.2–97.0 | 12 | 30 | 3.08 | 89.1–103.6 | 10 | 30 | 3.50 |
| 97.0–104.1 | 12 | 31 | 2.58 | 103.6–118.9 | 10 | 31 | 2.82 |
| 104.1–115.5 | 12 | 26 | 2.79 | 118.9–131.3 | 10 | 26 | 2.97 |
References
- Emmans, G.C.; Kyriazakis, I. Models of pig growth: Problems and proposed solutions. Livest. Prod. Sci. 1997, 51, 119–129. [Google Scholar] [CrossRef]
- Van Milgen, J.; Noblet, J.; Dourmad, J.Y.; Labussière, E.; Garcia-Launay, F.; Brossard, L. Precision pork production: Predicting the impact of nutritional strategies on carcass quality. Meat Sci. 2012, 92, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Van Milgen, J.; Valancogne, A.; Dubois, S.; Dourmad, J.Y.; Sève, B.; Noblet, J. InraPorc: A model and decision support tool for the nutrition of growing pigs. Anim. Feed Sci. Technol. 2008, 143, 387–405. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine, 11th revised ed.; The National Academies Press: Washington, DC, USA, 2012; p. 420. [Google Scholar]
- Fernández-Fígares, I.; Lachica, M.; Nieto, R.; Rivera-Ferre, M.G.; Aguilera, J.F. Serum profile of metabolites and hormones in obese (Iberian) and lean (Landrace) growing gilts fed balanced or lysine deficient diets. Livest. Sci. 2007, 110, 73–81. [Google Scholar] [CrossRef]
- Barea, R.; Nieto, R.; Aguilera, J.F. Effects of the dietary protein content and the feeding level on protein and energy metabolism in Iberian pigs growing from 50 to 100 kg body weight. Animal 2007, 1, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Nieto, R.; Lara, L.; Barea, R.; García-Valverde, R.; Aguinaga, M.A.; Conde-Aguilera, J.A.; Aguilera, J.F. Response analysis of the Iberian pig growing from birth to 150 kg body weight to changes in protein and energy supply. J. Anim. Sci. 2012, 90, 3809–3820. [Google Scholar] [CrossRef] [PubMed]
- Vautier, B.; Quiniou, N.; van Milgen, J.; Brossard, L. Accounting for variability among individual pigs in deterministic growth models. Animal 2013, 7, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Čandek-Potokar, M.; Nieto, R. European Local Pig Breeds–Diversity and Performance. A Study of Project Treasure; IntechOpen: London, UK, 2019; p. 303. [Google Scholar] [CrossRef]
- Freitas, A.B. Influência do Nível e Regime Alimentar em Pré-Acabamento Sobre o Crescimento e Desenvolvimento do porco Alentejano e suas Repercussões Sobre o Acabamento em Montanheira e com Alimento Comercial. Ph.D. Thesis, Universidade de Evora, Evora, Portugal, 1998. [Google Scholar]
- Lebret, B.; Dourmad, J.Y.; Mourot, J.; Pollet, P.Y.; Gondret, F. Production performance, carcass composition, and adipose tissue traits of heavy pigs: Influence of breed and production system. J. Anim. Sci. 2014, 92, 3543–3556. [Google Scholar] [CrossRef] [PubMed]
- Santos e Silva, J.; Ferreira-Cardoso, J.; Bernardo, A.; Pires da Costa, J.S. Conservation and development of the Bísaro pig. Characterisation and zootechnical evaluation of the breed for production and genetic management. In Proceedings of the Joint Session of the EAAP Commissions on Pig Production, Animal Genetics and Animal Nutrition, Zurich, Switzerland, 25 August 1999; Wenk, C., Fernandez, J.A., Dupuis, M., Eds.; Wageningen Press: Wageningen, The Netherlands, 2000; pp. 85–92. [Google Scholar]
- Rossi, A.; Ferrari, P.; Bossio, M.B.; Moncao, F.; Fusaro, A. Growth performance and meat quality of outdoor reared Calabrese pigs. In Proceedings of the 6th International Symposium on the Mediterranean Pig, Messina Capo d’Orlando, Italy, 11–13 October 2007; pp. 214–223. [Google Scholar]
- Ayuso, M.; Óvilo, C.; Fernández, A.; Nunez, Y.; Isabel, B.; Dazac, A.; López-Bote, C.J.; Rey, A.I. Effects of dietary vitamin A supplementation or restriction and its timing on retinol and α-tocopherol accumulation and gene expression in heavy pigs. Anim. Feed Sci. Technol. 2015, 202, 62–74. [Google Scholar] [CrossRef]
- Barea, R.; Nieto, R.; Vitari, F.; Domeneghini, C.; Aguilera, J.F. Effects of pig genotype (Iberian v. Landrace x Large White) on nutrient digestibility, relative organ weight and small intestine structure at two stages of growth. Animal 2011, 5, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Nieto, R.; Miranda, A.; García, M.A.; Aguilera, J.F. The effect of dietary protein content and feeding level on the rate of protein deposition and energy utilization in growing Iberian pigs from 15 to 50 kg body weight. Br. J. Nutr. 2002, 88, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Čandek-Potokar, M.; Batorek Lukač, N.; Tomažin, U.; Nieto, R. Performances de croissance des races locales de porcs selon la phase de production: Une étude analytique du projet TRESAURE. In Proceedings of the 51èmes Journées de la Recherche Porcine, Paris, France, 5–6 February 2019; pp. 205–210. [Google Scholar]
- Čandek-Potokar, M.; Batorek Lukač, N.; Tomažin, U.; Škrlep, M.; Nieto, R. Analytical review of productive performance of local pig breeds. In European Local Pig Breeds–Diversity and Performance. A Study of Project Treasure; Čandek Potokar, M., Nieto, R., Eds.; IntechOpen: London, UK, 2019; pp. 281–303. [Google Scholar] [CrossRef]




| Breed 1 | Data Source | No. of | Days | Initial | Final | Diet(s) Composition | Feed Intake, | |
|---|---|---|---|---|---|---|---|---|
| Pigs | on Trial | BW, kg | BW, kg | Energy, MJ NE/kg | Protein, % | kg/Pig/d | ||
| Alentejana_1 | [10] | 28–34 | 118 | 41.5 | 96.3 | 9.14 | 15.7 | 2.59 |
| Alentejana_2 | [10] | 13–39 | 136 | 35.3 | 95.1 | 8.51 | 15.2 | 2.45 |
| Basque_conv | [11] | 20 | 214 | 34.8 | 139.9 | 9.59 | 18.0 | 2.39 |
| 8.96 | 14.7 | |||||||
| Basque_alter | [11] | 20 | 206 | 35.3 | 146.3 | 9.59 | 18.0 | 2.68 |
| 8.96 | 14.7 | |||||||
| Bísara_1 | [12] | 24 | 209 | 29.8 | 144.3 | 9.51 | 15.0 | 2.34 |
| Bísara_2 | TREASURE project | 20 | 168 | 25.4 | 115.8 | 9.75 | 16.7 | 1.5 kg + corn (0.0–0.6 kg) |
| 9.43 | 15.3 | |||||||
| Corn: 9.90 | 8.6 | |||||||
| A.Calabrese_1 | [13] | 36 | 142 | 40.2 | 139.1 | 9.34 | 15.4 | 3.14 |
| 9.69 | 14.2 | |||||||
| A.Calabrese_2 | [13] | 36 | 142 | 33.8 | 135.2 | 9.22 | 15.5 | 3.00 |
| 9.70 | 14.2 | |||||||
| Cinta Senese | TREASURE project | 11 | 246 | 38.0 | 143.4 | 9.64 | 12.4 | 2.48 |
| 9.71 | 10.3 | |||||||
| Iberian_1 | [14] | 20–29 | 273 | 16.1 | 159.5 | 10.0 | 17.8 | 2.10 |
| 9.5 | 15.8 | |||||||
| 10.4 | 13.5 | |||||||
| Iberian_2a | [15] | 6–8 | 155 | 15.7 | 114 | 9.23 | 12.6 | 2.54 |
| Iberian_2b | [15] | 6–8 | 174 | 16.0 | 114 | 9.08 | 16.6 | 2.55 |
| Krškopolje_con | TREASURE project | 12 | 115 | 36.2 | 120.4 | 8.52 | 15.3 | 3.23 |
| 8.82 | 14.0 | |||||||
| Krškopolje_eco | TREASURE project | 12 | 115 | 35.0 | 124.3 | 8.69 | 15.0 | 3.20 |
| 8.83 | 14.0 | |||||||
| Mangalitsa | TREASURE project | 12 | 198 | 24.3 | 115.5 | 9.98 | 13.7 | 2.64 |
| 10.00 | 14.4 | |||||||
| Moravka | TREASURE project | 10–12 | 198 | 29.9 | 131.3 | 9.98 | 13.7 | 2.94 |
| 10.00 | 14.4 | |||||||
| Profile 2 | InraPorc Parameters | Performance 40–100 kg BW | |||||||
|---|---|---|---|---|---|---|---|---|---|
| NE50, MJ NE | NE100, MJ NE | BGompertz, /d | PDm, g/d | Age at 40 kg, d | Age at 100 kg, d | ADG, g/d | ADFI, MJ NE/d | SID Lys. req., g/d | |
| Alentejana_1 | 19.0 | 29.4 | 0.0143 | 45 | 154 | 281 | 468 | 24.2 | 5.8 |
| Alentejana_2 | 17.9 | 27.4 | 0.0070 | 47 | 148 | 282 | 449 | 22.1 | 6.0 |
| Basque_conv | 21.5 | 22.2 | 0.0096 | 72 | 114 | 212 | 613 | 23.9 | 8.4 |
| Basque_alter | 23.6 | 27.9 | 0.0096 | 72 | 117 | 219 | 588 | 22.1 | 8.3 |
| Bísara_1 | 18.1 | 24.5 | 0.0073 | 77 | 127 | 230 | 580 | 21.1 | 8.8 |
| Bísara_2 | 17.2 | 19.3 | 0.0100 | 81 | 121 | 229 | 554 | 18.5 | 9.1 |
| A.Calabrese_1 | 20.8 | 33.3 | 0.0123 | 97 | 160 | 241 | 746 | 26.2 | 10.8 |
| A.Calabrese_2 | 19.7 | 32.7 | 0.0163 | 105 | 150 | 228 | 774 | 25.4 | 11.7 |
| Cinta Senese | 15.7 | 27.8 | 0.0041 | 41 | 125 | 280 | 389 | 20.3 | 5.2 |
| Iberian_1 | 16.3 | 28.9 | 0.0024 | 67 | 143 | 258 | 526 | 20.8 | 7.8 |
| Iberian_2a | 24.0 | 30.4 | 0.0079 | 84 | 110 | 195 | 707 | 27.2 | 9.7 |
| Iberian_2b | 23.6 | 29.1 | 0.0083 | 69 | 113 | 210 | 619 | 26.6 | 8.2 |
| Krškopolje_con | 20.7 | 35.2 | 0.0183 | 116 | 120 | 190 | 854 | 27.1 | 12.8 |
| Krškopolje_eco | 23.2 | 30.7 | 0.0119 | 113 | 122 | 194 | 834 | 27.0 | 12.4 |
| Mangalitsa | 23.1 | 28.4 | 0.0101 | 50 | 206 | 323 | 515 | 26.0 | 6.2 |
| Moravka | 22.8 | 31.1 | 0.0087 | 54 | 173 | 282 | 550 | 20.2 | 6.6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brossard, L.; Nieto, R.; Charneca, R.; Araujo, J.P.; Pugliese, C.; Radović, Č.; Čandek-Potokar, M. Modelling Nutritional Requirements of Growing Pigs from Local Breeds Using InraPorc. Animals 2019, 9, 169. https://doi.org/10.3390/ani9040169
Brossard L, Nieto R, Charneca R, Araujo JP, Pugliese C, Radović Č, Čandek-Potokar M. Modelling Nutritional Requirements of Growing Pigs from Local Breeds Using InraPorc. Animals. 2019; 9(4):169. https://doi.org/10.3390/ani9040169
Chicago/Turabian StyleBrossard, Ludovic, Rosa Nieto, Rui Charneca, José Pedro Araujo, Carolina Pugliese, Čedomir Radović, and Marjeta Čandek-Potokar. 2019. "Modelling Nutritional Requirements of Growing Pigs from Local Breeds Using InraPorc" Animals 9, no. 4: 169. https://doi.org/10.3390/ani9040169
APA StyleBrossard, L., Nieto, R., Charneca, R., Araujo, J. P., Pugliese, C., Radović, Č., & Čandek-Potokar, M. (2019). Modelling Nutritional Requirements of Growing Pigs from Local Breeds Using InraPorc. Animals, 9(4), 169. https://doi.org/10.3390/ani9040169