Avian Liver: The Forgotten Organ


Simple Summary
Abstract
1. Introduction
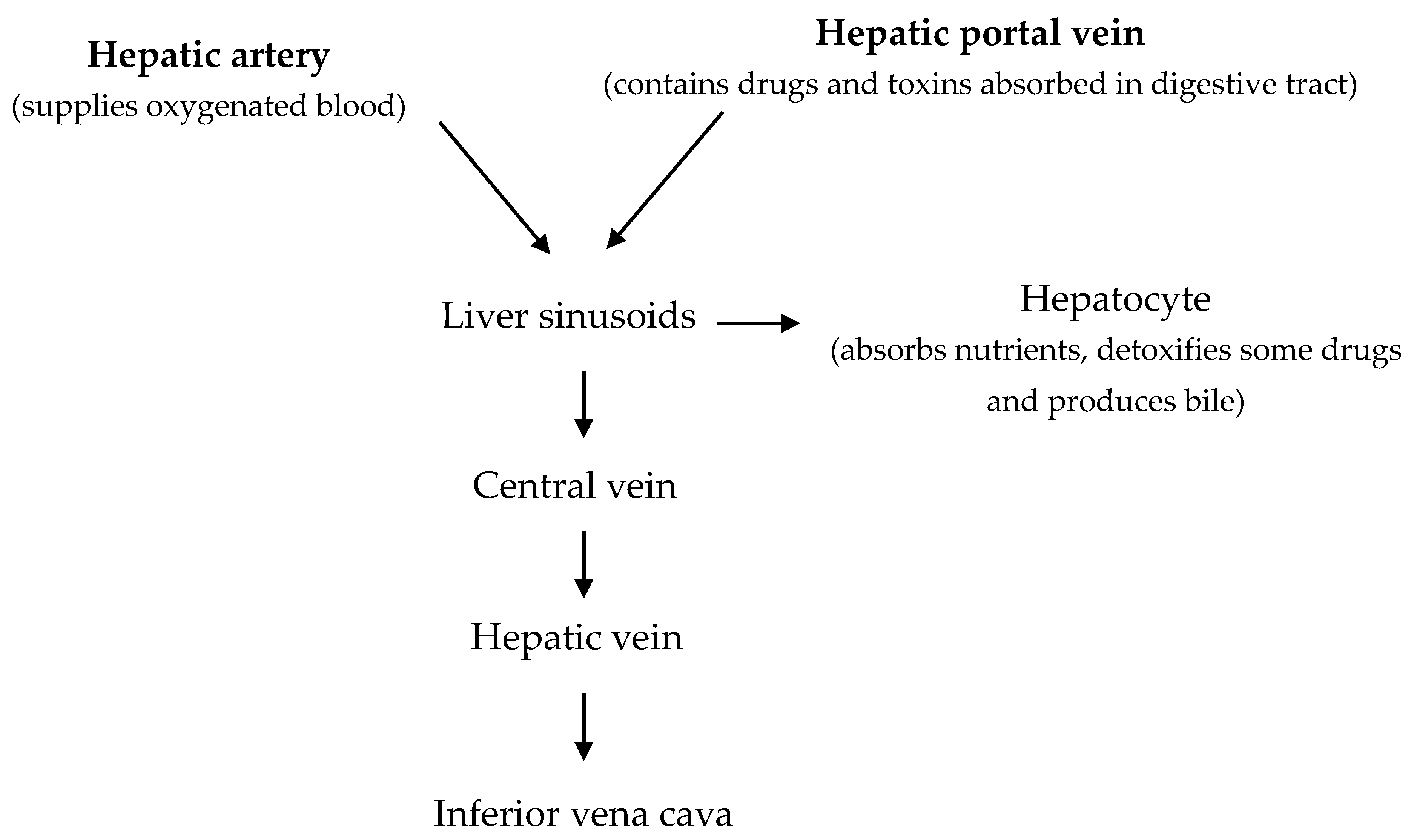
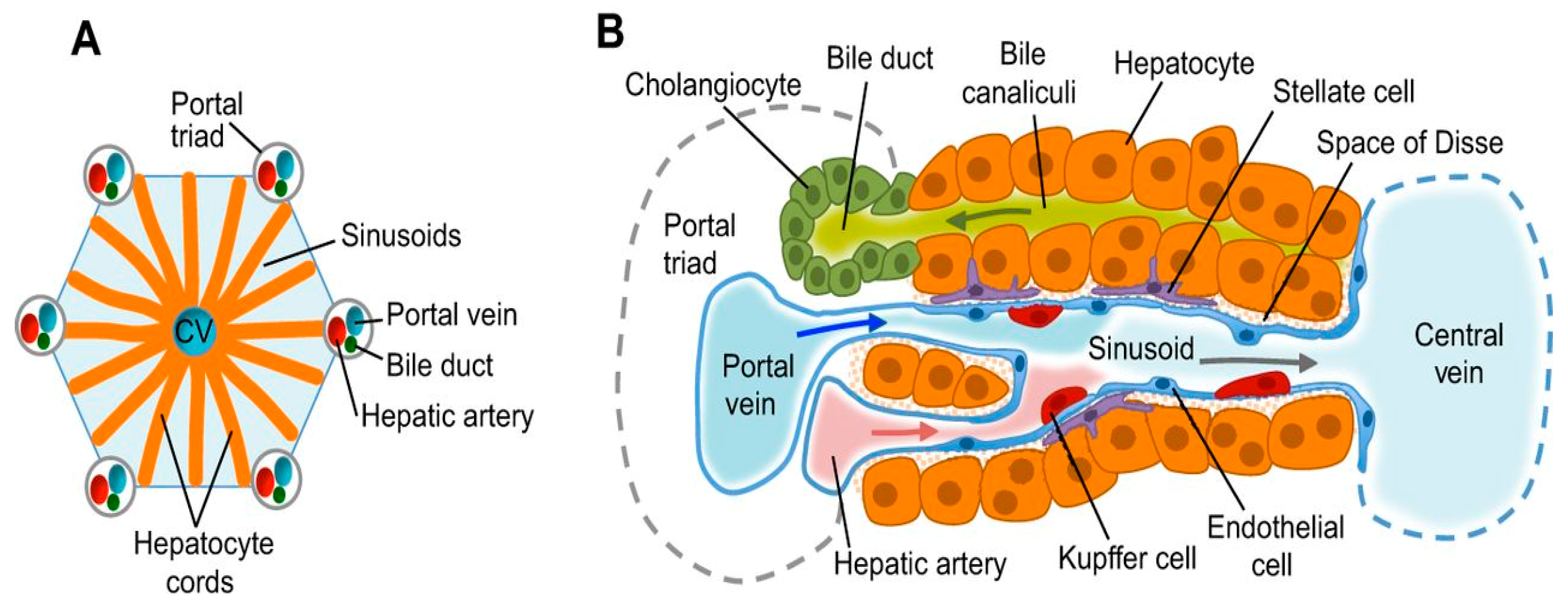
2. Structure and Functional Units in Avian Species
3. Composition and Size
4. Liver Functions
4.1. Fat Metabolism
4.1.1. Gall Bladder, Bile Formation and Secretion
4.1.2. Bile Composition and Functions
4.1.3. Lipogenesis and Lipoprotein Formation
4.2. Carbohydrate Metabolism
4.3. Protein Metabolism
4.4. Vitamin and Mineral Metabolism
4.5. Removal of Waste Products and Detoxification
5. Influence of Fat Sources on Liver Metabolism
6. Influence of Feed Restriction
7. Effects of Anti-Nutritional Factors
8. Effect of Dietary Fibre and Exogenous Carbohydrases
9. Effect of Feed Structure and Whole Grain Feeding
10. Effect of Nutrient Density
11. Liver Metabolic Disorder
Fatty Liver Haemorrhagic Syndrome (FLHS)
12. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Denbow, D.M. Gastrointestinal anatomy and physiology. In Sturkie’s Avian Physiology, 5th ed.; Whittow, G.C., Ed.; Academic Press: New York, NY, USA, 2000; pp. 299–325. [Google Scholar]
- Lucas, A.M.; Denington, E.M. Morphology of the chicken liver. Poult. Sci. 1956, 35, 793–806. [Google Scholar] [CrossRef]
- Kmiec, Z. Cooperation of liver cells in health and disease. Adv. Anat. Embryol. Cell Biol. 2001, 161, 1–151. [Google Scholar]
- Turk, D.E. The anatomy of the avian digestive tract as related to feed utilization. Poult. Sci. J. 1982, 60, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Akers, R.M.; Denbow, D.M. Digestive System. In Anatomy and Physiology of Domestic Animals, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; Chapter 17; pp. 483–527. [Google Scholar]
- Gordillo, M.; Evans, T.; Gouon-Evans, V. Orchestrating liver development. Development 2015, 142, 2094–2108. [Google Scholar] [CrossRef] [PubMed]
- Lumeij, J.T. Hepatology. In Avian Medicine: Principles and Application; Wingers Publishing, Inc.: Lake Worth, FL, USA, 1994; pp. 640–672. [Google Scholar]
- Bowes, V.A.; Julian, R.J. Organ weights of normal broiler chickens and those dying of sudden death syndrome. Can. Vet. J. 1988, 29, 153–156. [Google Scholar] [PubMed]
- Nir, I.; Nitsan, Z.; Mahagna, M. Comparative growth and development of the digestive organs and of some enzymes in broiler and egg type chicks after hatching. Br. Poult. Sci. 1993, 34, 523–532. [Google Scholar] [CrossRef]
- Ravindran, V.; Wu, Y.B.; Thomas, D.G.; Morel, P.C.H. Influence of whole wheat feeding on the development of gastrointestinal tract and performance of broiler chickens. Aust. J. Agric. Res. 2006, 57, 21–26. [Google Scholar] [CrossRef]
- Hermier, D. Lipoprotein metabolism and fattening in poultry. J. Nutr. 1997, 127, 805s–808s. [Google Scholar] [CrossRef]
- Tancharoenrat, P.; Ravindran, V.; Zaefarian, F.; Ravindran, G. Digestion of fat and fatty acids along the gastrointestinal tract of broiler chickens. Poult. Sci. 2014, 93, 371–379. [Google Scholar] [CrossRef]
- Bensadoun, A.; Rothfeld, A. The form of absorption of lipids in the chicken, Gallus domesticus. Proc. Soc. Exp. Biol. Med. 1972, 141, 814–817. [Google Scholar] [CrossRef]
- Griffin, H.; Grant, G.; Perry, M. Hydrolysis of plasma triacylglycerol-rich lipoproteins from immature and laying hens (Gallus domesticus) by lipoprotein lipase in vitro. Biochem. J. 1982, 206, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Cherian, G.; Holsonbake, T.B.; Goeger, M.P.; Bildfell, R. Dietary CLA alters yolk and tissue FA composition and hepatic histopathology of laying hens. Lipids 2002, 37, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.L.; Nesheim, M.C.; Young, R.J. Nutrition of the Chicken, 3rd ed.; M.L. Scott and Associates: Ithaca, NY, USA, 1982. [Google Scholar]
- Koeppen, B.M.; Stanton, B.A. Berne and Levy Physiology, 6th ed.; Mosby Elsevier: Philadelphia, PA, USA, 2008. [Google Scholar]
- Denbow, D.M. Gastrointestinal anatomy and physiology. In Sturkie’s Avian Physiology, 6th ed.; Scanes, C.G., Ed.; Academic Press: New York, NY, USA, 2015; pp. 337–366. [Google Scholar]
- Hurwitz, S.; Bar, A.; Katz, M.; Sklan, D.; Budowski, P. Absorption and secretion of fatty acids and bile acids in the intestine of the laying fowl. J. Nutr. 1973, 103, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Sklan, D.; Budowski, P.; Ascarelli, I.; Hurwitz, S. Lipid absorption and secretion in the chick: Effect of raw soybean meal. J. Nutr. 1973, 103, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Sklan, D.; Hurwitz, S.; Budowski, P.; Ascarelli, I. Fat digestion and absorption in chicks fed raw or heated soybean meal. J. Nutr. 1975, 105, 57–63. [Google Scholar] [CrossRef]
- Lavrenteva, G.F. Bile secretion in Chicken. Tru. Gor’ko. Sel’skokhoz. Inst. 1963, 13, 80–85. [Google Scholar]
- Andrews, C.J.H. Bile secretion in anesthetized Adelie penguin (Pygoscelis adeliae). J. Physiol. 1975, 246, 76P–77P. [Google Scholar]
- Duke, G.E. Alimentary canal: Secretion and digestion, special digestive functions, and absorption. In Avian Physiology; Sturkie, P.D., Ed.; Springer: New York, NY, USA, 1986; pp. 289–302. [Google Scholar]
- Argenzio, R.A. Secretions of stomach and accessory glands. In Dukes Physiology of Domestic Animals, 12th ed.; Reece, W.O., Ed.; Cornell University Press: Ithaca, NY, USA; London, UK, 2004; pp. 405–418. [Google Scholar]
- Lisbona, F.; Jiménez, M.; Esteller, A.; Lopez, M.A. Basal Biliary secretion in conscious chicken and role of enterohepatic circulation. Comp. Biochem. Physiol. 1981, 69A, 341–344. [Google Scholar] [CrossRef]
- Haslewood, G.A.D. The Biological Importance of Bile Salts; North-Holland research monographs; North-Holland: Amsterdam, The Netherlands, 1978. [Google Scholar]
- Krogdahl, A. Digestion and absorption of lipid in poultry. J. Nutr. 1985, 115, 675–685. [Google Scholar] [CrossRef]
- Chen, M.; Gratzel, M.; Thomas, J.K. Kinetic studies in bile acid micelles. J. Am. Chem. Soc. 1975, 97, 2052–2057. [Google Scholar] [CrossRef]
- Lindsay, O.B.; March, B.E. Intestinal absorption of bile salts in the cockerel. Poult. Sci. 1967, 46, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.L.; Himes, J.A.; Cornelius, C.E. Bilirubin and biliverdin excretion by the chicken. Am. J. Physiol. 1974, 226, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.J. Physiology of digestive tract. In Physiology and Biochemistry of Domestic Fowl; Bell, D.J., Freeman, B.M., Eds.; Academic Press: London, UK, 1983; Volume 4, pp. 31–47. [Google Scholar]
- Elkin, R.G.; Wood, K.V.; Hagey, L.R. Biliary bile acid profiles of domestic fowl as determined by high performance liquid chromatography and fast atom bombardment mass spectrometry. Comp. Biochem. Physiol. B 1990, 96, 157–161. [Google Scholar] [CrossRef]
- Yeh, Y.H.; Hwang, D.F. High-performance liquid chromatographic determination for bile components in fish, chicken and duck. J. Chromatogr. B 2001, 751, 1–8. [Google Scholar] [CrossRef]
- Serafin, J.A.; Nesheim, M.C. Influence of dietary heat-labile factors in soybean meal upon bile acid pools and turnover in the chick. J. Nutr. 1970, 100, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Klasing, K.C. Nutritional strategies and adaptations. In Comparative Avian Nutrition; CAB International: New York, NY, USA, 1998; pp. 71–124. [Google Scholar]
- Richards, M.P.; Poch, S.M.; Coon, C.N.; Rosebrough, R.W.; Ashwell, C.M.; McMurtry, J.P. Feed Restriction Significantly Alters Lipogenic Gene Expression in Broiler Breeder Chickens. J. Nutr. 2003, 133, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Walzem, R.L.; Davis, P.A.; Hansen, R.J. Overfeeding increases very low-density lipoprotein diameter and causes the appearance of a unique lipoprotein particle in association with failed yolk deposition. J. Lipid Res. 1994, 35, 1354–1366. [Google Scholar]
- Alvarenga, R.R.; Zangeronimo, M.G.; Pereira, L.J.; Rodrigues, P.B.; Gomide, E.M. Lipoprotein metabolism in poultry. World’s Poult. Sci. J. 2011, 67, 431–440. [Google Scholar] [CrossRef]
- Speake, B.K.; Murray, A.M.; Noble, R.C. Transport and transformations of yolk lipids during development of the avian embryo. Prog. Lipid Res. 1998, 37, 1–32. [Google Scholar] [CrossRef]
- Griffin, H.; Hermier, D. Plasma lipoprotein metabolism and fattening in poultry. In Leanness in Domestic Birds; Butterworths: London, UK, 1988; pp. 175–201. [Google Scholar]
- Siuta-Mangano, P.; Howard, S.C.; Lennarz, W.J.; Lane, M.D. Synthesis, processing, and secretion of apolipoprotein B by the chick liver cell. J. Biol. Chem. 1982, 257, 4292–4300. [Google Scholar]
- Tarlow, D.M.; Watkins, P.A.; Reed, R.E.; Miller, R.S.; Zwergel, E.E.; Lane, M.D. Lipogenesis and the synthesis and secretion of very low-density lipoprotein by avian liver cells in nonproliferating monolayer culture. Hormonal effects. J. Cell Biol. 1977, 73, 332–353. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.M. The mobilisation of hepatic glycogen in Gallus domesticus at the end of incubation. Comp. Biochem. Physiol. 1969, 28, 1169–1176. [Google Scholar] [CrossRef]
- Goodridge, A.G. Metabolism of glucose-U-14C in vitro in adipose tissue from embryonic and growing chicks. Am. J. Physiol. 1986, 214, 897–901. [Google Scholar] [CrossRef] [PubMed]
- Chin, T.Y.; Quebbemann, A.J. Quantitation of renal uric acid synthesis in the chicken. Am. J. Physiol. 1987, 234, F446–F451. [Google Scholar] [CrossRef]
- Campbell, T.W. Avian Hematology and Cytology, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1995. [Google Scholar]
- Holick, M.F. Phylogenetic and evolutionary aspects of vitamin D from phytoplankton to humans. In Vertebrate Endocrinology: Fundamentals and Biomedical Implications; Pang, P.K.T., Schreibman, M.P., Eds.; Academic Press, Inc. (Harcourt Brace Jovanovich): Orlando, FL, USA, 1989. [Google Scholar]
- Konig, B.; Spielmann, J.; Haase, K.; Brandsch, C.; Kluge, H.; Stangl, G.I.; Eder, K. Effects of fish oil and conjugated linoleic acids on expression of target genes of PPARα and sterol regulatory element-binding proteins in the liver of laying hens. Br. J. Nutr. 2008, 100, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Royan, M.; Meng, G.Y.; Othman, F.; Sazili, A.Q.; Navidshad, B. Effects of conjugated linoleic acid, fish oil and soybean oil on PPARs (α & γ) mRNA expression in broiler chickens and their relation to body fat deposits. Int. J. Mol. Sci. 2011, 12, 8581–8595. [Google Scholar] [PubMed]
- Campioli, E.; Rustichelli, C.; Avallone, R. n-3 dietary supplementation and lipid metabolism: Differences between vegetable- and fish-derived oils. J. Funct. Foods 2012, 4, 207–212. [Google Scholar] [CrossRef]
- Huang, C.H.; Lin, W.L.; Chu, J.H. Dietary lipid level influences fatty acid profiles, tissue composition, and lipid peroxidation of soft-shelled turtle, Pelodiscus sinensis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 142, 383–388. [Google Scholar] [CrossRef]
- Senkoylu, N.; Dale, N. Nutritional evaluation of a high-oil sunflower meal in broiler starter diets. J. Appl. Poult. Res. 2006, 15, 40–47. [Google Scholar] [CrossRef]
- Smink, W.; Gerrits, W.J.J.; Hovenier, R.; Geelen, M.J.H.; Verstegen, M.W.A.; Beynen, A.C. Effect of dietary fat sources on fatty acid deposition and lipid metabolism in broiler chickens. Poult. Sci. 2010, 89, 2432–2440. [Google Scholar] [CrossRef]
- Pinchasov, Y.; Nir, I. Effect of dietary polyunsaturated fatty acid concentration on performance, fat deposition, and carcass fatty acid composition in broiler chickens. Poult. Sci. 1992, 71, 1504–1512. [Google Scholar] [CrossRef] [PubMed]
- Ferrell, C.L. Contribution of visceral organs to animal energy expenditures. J. Anim. Sci. 1988, 66, 23–34. [Google Scholar]
- Pearson, A.W.; Greenwood, N.M.; Butler, E.J.; Fenwick, G.R. Biochemical changes in layer and broiler chickens when fed on a high-glucosinolate rapeseed meal. Br. Poult. Sci. 1983, 24, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Stangassinger, M.; Giesecke, D. Splanchnic metabolism of glucose and related energy substrates. In Control of Digestion and Metabolism in Ruminants; Milligan, L.P., Grovum, W.L., Dobson, A., Eds.; Prentice-Hall: Englewood Cliffs, NJ, USA, 1986; pp. 347–366. [Google Scholar]
- Knight, C.D.; Dibner, J.J. Nutritional programming in hatchling poultry: Why a good start is important. Poult. Dig. 1998, 57, 20–26. [Google Scholar]
- Canas, R.; Romero, J.J.; Baldwin, R.L. Maintenance energy requirements during lactation in rats. J. Nutr. 1982, 112, 1876–1880. [Google Scholar] [CrossRef] [PubMed]
- Lacy, M.P.; Van Krey, H.P.; Skewes, P.A.; Denbow, D.M. Effect of intrahepatic glucose infusion on feeding in heavy and light breed chicks. Poult. Sci. 1985, 64, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Lacy, M.P.; Van Krey, H.P.; Skewes, P.A.; Denbow, D.M. Food intake in the domestic fowl: Effect of intrahepatic lipid and amino acid infusions. Physiol. Behav. 1986, 36, 533–538. [Google Scholar] [CrossRef]
- Bartov, I. Lack of interactive effect of nicarbazin and dietary energy-to-protein ratio on performance and abdominal fat pad weight of broiler chicks. Poult. Sci. 1989, 68, 1535–1539. [Google Scholar] [CrossRef]
- Bartov, I. Effect of feed withdrawal on yield, fat content and fatty acid composition of various tissues in broilers. In Proceedings of the XIX World’s Poultry Congress, Amsterdam, The Netherlands, 19–24 September 1992; pp. 195–199. [Google Scholar]
- Bartov, I. Inter-relationship between the effects of dietary factors and feed withdrawal on the content and composition of liver fat in broiler chicks. Poult. Sci. 1996, 75, 632–641. [Google Scholar] [CrossRef]
- Fisher, C. Fat deposition in broilers. In Fats in Animal Nutrition; Wiseman, J., Ed.; Butterworths: London, UK, 1984; pp. 437–470. [Google Scholar]
- Imaizumi, K.; Sugano, M.; Wada, M. Nutritional regulation of lipid metabolism in rats. Part I. The effect of overnight fasting on the concentration and composition of liver and its microsomal lipids. Agric. Biol. Chem. 1972, 36, 234–241. [Google Scholar]
- Leveille, G.A.; Romsos, D.R.; Yu, Y.Y.; O’Hea, E.K. Lipid biosynthesis in the chick. A consideration of site of synthesis, influence of diet and possible regulatory mechanisms. Poult. Sci. 1975, 54, 1075–1091. [Google Scholar] [CrossRef]
- Leveille, G.A.; Tillotson, J.A.; Sauberlich, H.E. Fatty acid composition of plasma and liver lipid components as influenced by diet in the growing chick. J. Nutr. 1963, 81, 357–362. [Google Scholar] [PubMed]
- Fujii, M.; Odawara, K.; Ohyama, M.; Fukunaga, T.; Koga, K. Fatty acid compositions of triacylglycerols and phospholipids in hen liver lipids before and after sexual maturity. Poult. Sci. 1985, 64, 1371–1376. [Google Scholar] [CrossRef] [PubMed]
- Shapira, N.; Nachtomi, E.; Nir, I. Effects of isocaloric supplements of glucose or soyabean oil on lipids in tissues and plasma lipoproteins of starved and overfed chicks. Nutr. Metab. 1979, 23, 368–383. [Google Scholar] [CrossRef] [PubMed]
- Rosebrough, R.W.; Steele, N.C.; McMurtry, J.P.; Plavnik, I. Effect of early feed restriction in broilers. II. Lipid metabolism. Growth 1986, 50, 217–227. [Google Scholar] [PubMed]
- Fontana, E.A.; Weaver, W.D.; Denbow, D.M.; Watkins, B.A. Early feed restriction of broilers: Effects on abdominal fat pad, liver, and gizzard weights, fat deposition and carcass composition. Poult. Sci. 1993, 72, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Hocking, P.M.; Maxwell, M.H.; Mitchell, M.A. Relationship between the degree of food restriction and welfare indices in broiler breeder females. Br. Poult. Sci. 1996, 37, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Hocking, P.M.; Maxwell, M.H.; Mitchell, M.A. Welfare assessment of broiler breeder and layer females subjected to food restriction and limited access to water during rearing. Br. Poult. Sci. 1993, 34, 443–458. [Google Scholar] [CrossRef]
- Zubair, A.K.; Leeson, S. Effect of early feed restriction and realimentation on heat production and changes in sizes of digestive organs of male broilers. Poult. Sci. 1994, 73, 529–538. [Google Scholar] [CrossRef]
- Susbilla, P.J.; Frankel, T.L.; Parkinson, G.; Gow, C.B. Weight of internal organs and carcass yield of early food restricted broilers. Br. Poult. Sci. 1994, 35, 677–685. [Google Scholar] [CrossRef]
- Liener, I.E. Toxic Constituents of Plant Foodstuffs; Academic Press: New York, NY, USA, 1980; p. 502. [Google Scholar]
- Ortiz, L.T.; Alzueta, C.; Trevino, J.; Castano, M. Effects of faba bean tannins on the growth and histological structure of the intestinal tract and liver of chicks and rats. Br. J. Nutr. 1994, 35, 743–754. [Google Scholar] [CrossRef]
- Emiola, I.A.; Ologhobo, A.D.; Gous, R.M. Performance and histological responses of internal organs of broiler chicken fed raw, dehulled, and aqueous and dry-heated kidney bean meals. Poult. Sci. 2007, 86, 1234–1240. [Google Scholar] [CrossRef]
- Eisenberg, F.; Bolden, A.H. Biosynthesis of inositol in rat testis homogenate. Biochem. Biophys. Res. Commun. 1963, 12, 72–77. [Google Scholar] [CrossRef]
- Lee, S.A.; Bedford, M.R. Inositol—An effective growth promotor? World’s Poult. Sci. J. 2016, 72, 743–760. [Google Scholar] [CrossRef]
- Hayashi, E.; Maeda, T.; Tomita, T. The effect of myo-inositol deficiency on lipid metabolism in rats. II. The mechanism of triacylglycerol accumulation in the liver of myo-inositol deficient rats. Biochim. Biophys. Acta 1974, 360, 146–155. [Google Scholar] [CrossRef]
- Croze, M.L.; Soulage, C.O. Potential role and therapeutic interests of myoinositol in metabolic diseases. Biochimie 2013, 95, 1811–1827. [Google Scholar] [CrossRef]
- Croze, M.L.; Géloën, A.; Soulage, C.O. Abnormalities in myo-inositol metabolism associated with type 2 diabetes in mice fed a high-fat diet: Benefits of a dietary myo-inositol supplementation. Br. J. Nutr. 2015, 113, 1862–1875. [Google Scholar] [CrossRef]
- Liu, N.; Ru, Y.; Wang, J.; Xu, T. Effect of dietary sodium phytate and microbial phytase on the lipase activity and lipid metabolism of broiler chickens. B. J. Nutr. 2010, 103, 862–868. [Google Scholar] [CrossRef]
- Katayama, T. Effect of Dietary Sodium Phytate on the Hepatic and Serum Levels of Lipids and on the Hepatic Activities of NADPH-generating Enzymes in Rats Fed on Sucrose. Biosci. Biotechnol. Biochem. 1995, 59, 1159–1160. [Google Scholar] [CrossRef]
- Viveros, A.; Brenes, A.; Centeno, C. Effects of microbial phytase supplementation on mineral utilization and serum enzyme activities in broiler chicks fed different levels of phosphorus. Poult. Sci. 2002, 81, 1172–1183. [Google Scholar] [CrossRef]
- Karadas, F.; Pirgozliev, V.; Acamovic, T.; Bedford, M.R. The effects of dietary phytase activity on the concentration of Coenzyme Q10 in the liver of young turkeys and broilers. Br. Poult. Abstr. 2005, 1, 1–2. [Google Scholar] [CrossRef]
- Karadas, F.; Pirgozliev, V.; Pappas, A.C.; Acamovic, T.; Bedford, M.R. Effects of different dietary phytase activities on the concentration of antioxidants in the liver of growing broilers. J. Anim. Physio. Anim. Nutr. 2010, 94, 519–526. [Google Scholar] [CrossRef]
- Sharma, N.K.; Choct, M.; Wu, S.B.; Smillie, R.; Morgan, N.; Omar, A.S.; Sharma, N.; Swick, R.A. Performance, litter quality and gaseous odour emissions of broilers fed phytase supplemented diets. Anim. Nutr. 2016, 2, 288–295. [Google Scholar] [CrossRef]
- Rougière, N.; Carré, B. Comparison of gastrointestinal transit times between chickens from D+ and D− genetic lines selected for divergent digestion efficiency. Animal 2010, 4, 1861–1872. [Google Scholar] [CrossRef]
- Mateos, G.G.; Lázaro, R.; Gracia, M. The feasibility of using nutritional modifications to replace drugs in poultry feeds. J. Appl. Poult. Res. 2002, 11, 437–452. [Google Scholar] [CrossRef]
- Mateos, G.G.; Jiménez-Moreno, E.; Serrano, M.P.; Lázaro, R. Poultry response to high levels of dietary fiber sources varying in physical and chemical characteristics. J. Appl. Poult. Res. 2012, 21, 156–174. [Google Scholar] [CrossRef]
- Hetland, H.; Svihus, B.; Krogdahl, Å. Effects of oat hulls and wood shavings on digestion in broilers and layers fed diets based on whole or ground wheat. Br. Poult. Sci. 2003, 44, 275–282. [Google Scholar] [CrossRef]
- Hetland, H.; Svihus, B.; Choct, M. Role of insoluble fiber on gizzard activity in layers. J. Appl. Poult. Res. 2005, 14, 38–46. [Google Scholar] [CrossRef]
- Amerah, A.M.; Ravindran, V.; Lentle, R.G.; Thomas, D.G. Influence of feed particle size on the performance, energy utilization, digestive tract development and digestive parameter of broiler starters. Poult. Sci. 2007, 86, 2615–2623. [Google Scholar] [CrossRef]
- González-Alvarado, J.M.; Jiménez-Moreno, E.; González-Sánchez, D.; Lázaro, R.; Mateos, G.G. Effect of inclusion of oat hulls and sugar beet pulp in the diet on productive performance and digestive traits of broilers from 1 to 42 days of age. Anim. Feed Sci. Technol. 2010, 162, 37–46. [Google Scholar] [CrossRef]
- Svihus, B. Limitation to wheat starch digestion in growing broiler chickens: A brief review. Anim. Prod. Sci. 2011, 51, 583–589. [Google Scholar] [CrossRef]
- Akiba, Y.; Matsumoto, T. The role of dietary fibers on hepatic lipid metabolism in chicks. Nutr. Rep. Int. 1982, 26, 223–230. [Google Scholar]
- Akiba, Y.; Matsumoto, T. Effects of force-feeding and dietary cellulose on liver lipid accumulation and lipid composition of liver and plasma in growing chicks. J. Nutr. 1978, 108, 739–748. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Kiarie, E.; Nyachoti, C.M. Growth performance, organ weights, and blood parameters of broilers fed diets containing expeller-extracted canola meal. Poult. Sci. 2011, 90, 2520–2527. [Google Scholar] [CrossRef]
- Parke, D.V.; Ioannides, C. The role of nutrition in toxicology. Annu. Rev. Nutr. 1981, 1, 207–234. [Google Scholar] [CrossRef]
- Bauman, P.F.; Smith, K.; Bray, T.M. The effect of dietary protein and sulfur amino acids on hepatic glutathione concentration and glutathione-dependent enzyme activities in the rat. Can. J. Physiol. Pharmacol. 1988, 66, 1048–1052. [Google Scholar] [CrossRef]
- Mossami, A. Effects of Different Inclusions of Oat Hulls on Performance, Carcass Yield and Gut Development in Broiler Chickens. Master’s Thesis, Swedish University of Agricultural Science, Uppsalla, Sweden, 2011. [Google Scholar]
- Zhu, H.L.; Hu, L.L.; Hou, Y.Q.; Zhang, J.; Ding, B.Y. Effects of enzyme supplementation on performance and digestive parameters of broilers fed corn-soybean diets. Poult. Sci. 2014, 93, 1704–1712. [Google Scholar] [CrossRef]
- Dworkin, L.D.; Levine, G.M.; Farber, J.J.; Spector, N.H. Small intestinal mass of the rat is partially determined by indirect effects on intraluminal nutrition. Gastroenterology 1976, 71, 626–630. [Google Scholar]
- Wang, Z.R.; Qiao, S.Y.; Lu, W.Q.; Li, D.F. Effects of enzyme supplementation on performance, nutrient digestibility, gastrointestinal morphology, and volatile fatty acid profiles in the hindgut of broilers fed wheat-based diets. Poult. Sci. 2005, 84, 875–881. [Google Scholar] [CrossRef]
- Brenes, A.; Smith, J.; Guenter, W.; Marquardt, R.R. Effect of enzyme supplementation on the performance and digestive tract size of broiler chickens fed wheat- and barley-based diets. Poult. Sci. 1993, 72, 1731–1739. [Google Scholar] [CrossRef]
- Wu, Y.B.; Ravindran, V. Influence of whole wheat inclusion and xylanase supplementation on the performance, digestive tract measurements and carcass characteristics of broiler chickens. Anim. Feed Sci. Technol. 2004, 116, 129–139. [Google Scholar] [CrossRef]
- Lázaro, R.; Latorre, M.A.; Medel, P.; Gracia, M.; Mateos, G.G. Feeding regimen and enzyme supplementationto rye-based diets for broilers. Poult. Sci. 2004, 83, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Józefiak, D.; Rutkowski, A.; Jensen, B.B.; Engberg, R.M. The effect of β-glucanase supplementation of barley- and oat-based diets on growth performance and fermentation in broiler chicken gastrointestinal tract. Br. Poult. Sci. 2006, 47, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Sayyazadeh, H.; Rahimi, G.; Rezaei, M. Influence of enzyme supplementation of maize, wheat and barley-based diets on the performance of broiler chickens. Pak. J. Biol. Sci. 2006, 9, 616–621. [Google Scholar]
- Al-marzooqi, W.; Leeson, S. Effect of dietary lipase enzyme on gut morphology, gastric motility, and long-term performance of broiler chicks. Poult. Sci. 2000, 79, 956–960. [Google Scholar] [CrossRef] [PubMed]
- Emadinia, A.; Toghyani, M.; Gheisari, A.; Tabeidian, S.A.; Ale Saheb Fosoul, S.S.; Mohammadrezaei, M. Effect of wet feeding and enzyme supplementation on performance and immune responses of broiler chicks. J. Appl. Anim. Res. 2014, 42, 32–37. [Google Scholar] [CrossRef]
- Parsaie, S.; Shariatmadari, F.; Zamiri, M.J.; Khajeh, K. Influence of wheat-based diets supplemented with xylanase, bile acid and antibiotics on performance, digestive tract measurements and gut morphology of broilers compared with a maize-based diet. Br. Poult. Sci. 2007, 48, 594–600. [Google Scholar] [CrossRef]
- Józefiak, D.; Rutkowski, A.; Jensen, B.B.; Engberg, M. Effects of dietary inclusion of triticale, rye and wheat and xylanase supplementation on growth performance of broiler chickens and fermentation in the gastrointestinal tract. Anim. Feed Sci. Technol. 2007, 132, 79–93. [Google Scholar] [CrossRef]
- Iji, P.A.; Saki, A.A.; Tivey, D.R. Intestinal structure and function of broiler chickens on diets supplemented with a mannan oligosaccharide. J. Sci. Food Agric. 2001, 81, 1186–1192. [Google Scholar] [CrossRef]
- Abdollahi, M.R.; Ravindran, V.; Wester, T.J.; Ravindran, G.; Thomas, D.V. Influence of feed form and conditioning temperature on performance, apparent metabolisable energy and ileal digestibility of starch and nitrogen in broiler starters fed wheat-based diet. Anim. Feed Sci. Technol. 2011, 168, 88–99. [Google Scholar] [CrossRef]
- Abdollahi, M.R.; Ravindran, V.; Wester, T.J.; Ravindran, G.; Thomas, D.V. Influence of pellet diameter and length on the quality of pellets and, performance, nutrient utilisation and digestive tract development of broilers fed wheat-based diets. Br. Poult. Sci. 2013, 54, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.R.; Ravindran, V.; Wester, T.J.; Ravindran, G.; Thomas, D.V. The effect of manipulation of pellet size (diameter and length) on pellet quality and performance, apparent metabolisable energy and ileal nutrient digestibility in broilers fed maize-based diets. Anim. Prod. Sci. 2013, 53, 114–120. [Google Scholar] [CrossRef]
- Abdollahi, M.R.; Ravindran, V.; Svihus, B. Influence of feed form on growth performance, ileal nutrient digestibility, and energy utilisation in broiler starters fed a sorghum-based diet. Livest. Sci. 2014, 165, 80–86. [Google Scholar] [CrossRef]
- Selle, P.H.; Liu, S.Y.; Cai, J.; Cowieson, A.J. Steam-pelleting temperatures and grain variety of finely-ground, sorghum-based broiler diets. I. Influence on growth performance, relative gizzard weights, nutrient utilisation, starch and nitrogen digestibility. Anim. Prod. Sci. 2014, 54, 339–346. [Google Scholar] [CrossRef]
- Marks, H.L.; Pesti, G.M. The roles of protein level and diet form in water consumption and abdominal fat pad deposition of broilers. Poult. Sci. 1984, 63, 1617–1625. [Google Scholar] [CrossRef] [PubMed]
- Plavnik, I.; Hurwitz, S. Effect of dietary protein, energy, and feed pelleting on the response of chicks to early feed restriction. Poult. Sci. 1989, 68, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Rezaeipour, V.; Gazani, S. Effects of feed form and feed particle size with dietary L-threonine supplementation on performance, carcass characteristics and blood biochemical parameters of broiler chickens. J. Anim. Sci. Technol. 2014, 56, 20. [Google Scholar] [CrossRef]
- Perez-Maldonado, R.A.; Mannion, P.F.; Farrel, D.J. The effect of steam and cold pelleting of four grain legumes at different inclusion levels in broiler starter diets. Proc. Aust. Poult. Sci. 1997, 9, 133–137. [Google Scholar]
- Preston, C.M.; McCracken, K.J.; McAllister, A. Effect of diet form and enzyme supplementation on growth, efficiency and energy utilisation of wheat-based diets for broilers. Br. Poult. Sci. 2000, 41, 324–331. [Google Scholar] [CrossRef]
- Scott, T.A. Evaluation of lighting programs, diet density, and short-term use of mash as compared to crumbled starter to reduce incidence of sudden death syndrome in broiler chicks to 35 days of age. Can. J. Anim. Sci. 2002, 82, 375–383. [Google Scholar] [CrossRef]
- González-Alvarado, J.M.; Jiménez-Moreno, E.; Lázaro, R.; Mateos, G.G. Effects of cereal, heat processing, and fiber on productive performance and digestive traits of broilers. Poult. Sci. 2007, 86, 1705–1715. [Google Scholar] [CrossRef] [PubMed]
- Mirghelenj, S.A.; Golian, A. Effects of feed form on development of digestive tract, performance and carcass traits of broiler chickens. J. Anim. Vet. Adv. 2009, 8, 1911–1915. [Google Scholar]
- Ghobadi, Z.; Karimi, A. Effect of feed processing and enzyme supplementation of wheat-based diets on performance of broiler chicks. J. Appl. Anim. Res. 2012, 40, 260–266. [Google Scholar] [CrossRef]
- Naderinejad, S.; Zaefarian, F.; Abdollahi, M.R.; Hassanabadi, A.; Kermanshahi, H.; Ravindran, V. Influence of feed form and particle size on performance, nutrient utilisation, and gastro intestinal tract development and morphometry in broiler starters fed maize-based diets. Anim. Feed Sci. Technol. 2016, 215, 92–104. [Google Scholar] [CrossRef]
- Benedetti, M.P.; Sartori, J.R.; Carvalho, F.B.; Pereira, L.A.; Fascina, V.B.; Stradiotti, A.C.; Pezzato, A.C.; Costa, C.; Ferreira, J.G. Corn texture and particle size in broiler diets. Rev. Bras. Cienc. Avic. 2011, 13, 227–234. [Google Scholar] [CrossRef]
- Favero, A.; Maiorka, A.; Dahlke, F.; Meurer, R.F.P.; Oliveira, R.S.; Sens, R.F. Influence of feed form and corn particle size on the live performance and digestive tract development of turkeys. J. Appl. Poult. Res. 2009, 18, 772–779. [Google Scholar] [CrossRef]
- Singh, Y.; Amerah, A.M.; Ravindran, V. Whole grain feeding: Methodologies and effects on performance, digestive tract development and nutrient utilisation of poultry. Anim. Feed Sci. Technol. 2014, 190, 1–18. [Google Scholar] [CrossRef]
- Nahas, J.; Lefrancois, M.R. Effects of feeding locally grown whole barley with or without enzyme addition and whole wheat on broiler performance and carcass trait. Poult. Sci. 2001, 80, 195–202. [Google Scholar] [CrossRef]
- Singh, Y.; Ravindran, V.; Wester, T.J.; Molan, A.L.; Ravindran, G. Influence of feeding coarse corn on performance, nutrient utilisation, digestive tract measurements, carcass characteristics, and cecal microflora counts of broilers. Poult. Sci. 2014, 93, 607–616. [Google Scholar] [CrossRef]
- Svihus, B.; Juvik, E.; Hetland, H.; Krogdahl, A. Causes for improvement in nutritive value of broiler chicken diets with whole wheat instead of ground wheat. Br. Poult. Sci. 2004, 45, 55–60. [Google Scholar] [CrossRef]
- Aghazadeh, A.M.; Taha Yazdi, M. Effect of butyric acid supplementation and whole wheat inclusion on the performance and carcass traits of broilers. S. Afr. J. Anim. Sci. 2012, 42, 241–248. [Google Scholar] [CrossRef]
- Mabelebele, M.; Gous, R.M.; Masey O’Neil, H.V.; Iji, P.A. Whole sorghum inclusion and feed form on performance and nutrient digestibility of broiler chickens. J. Appl. Anim. Nutr. 2018, 6, e5. [Google Scholar] [CrossRef]
- Salway, J.G. Metabolism at a Glance, 3rd ed.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2004. [Google Scholar]
- Lamot, D.M.; Sapkota, D.; Wijtten, P.J.A.; van den Anker, I.; Heetkamp, M.J.W.; Kemp, B.; van den Brand, H. Diet density during the first week of life: Effects on growth performance, digestive organ weight, and nutrient digestion of broiler chickens. Poult. Sci. 2018. [Google Scholar] [CrossRef]
- Buyse, J.; Geypens, B.; Malheiros, R.D.; Moraes, V.M.; Swennen, Q.; Decuypere, E. Assessment of age-related glucose oxidation rates of broiler chickens by using stable isotopes. Life Sci. 2004, 75, 2245–2255. [Google Scholar] [CrossRef] [PubMed]
- Crespo, R.; Shivaprasad, H.L. Developmental, metabolic, and other noninfectious disorders. In Diseases of Poultry, 11th ed.; Iowa State Press: Ames, IA, USA, 2003; pp. 1055–1102. [Google Scholar]
- Butler, E.J. Fatty liver disease in the domestic fowl: A review. Avian Pathol. 1976, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wolford, J.H.; Polin, D. Lipid accumulation and hemorrhage in livers of laying chickens. A study on fatty liver-hemorrhagic syndrome (FLHS). Poult. Sci. 1972, 51, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Wolford, J.H.; Murphy, D. Effect of diet on fatty liver-hemorrhagic syndrome incidence in laying chickens. Poult. Sci. 1972, 51, 2087–2094. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.C. Nutritional and metabolic aspects of fatty liver disease in poultry. Vet. Q. 1979, 1, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Meijering, A. Fatty liver syndrome in laying hens. An attempt to review. World’s Poult. Sci. J. 1980, 35, 79–94. [Google Scholar] [CrossRef]
- Butler, E. Lipid metabolism in the fowl under normal and abnormal circumstances. Proc. Nutr. Soc. 1975, 34, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Polin, D.; Wolford, J.H. Various types of diets, sources of energy, and positive energy balance in the induction of fatty liver hemorrhagic syndrome. Poult. Sci. 1976, 55, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Pearson, A.W.; Arkhipov, A.V.; Butler, E.J.; Laursen-Jones, A.P. Influence of dietary cereal and energy content on the accumulation of lipids in the liver in fatty liver haemorrhagic syndrome in the fowl. Res. Vet. Sci. 1978, 24, 72–76. [Google Scholar] [CrossRef]
- Haghighi-Rad, F.; Polin, D. Lipid: The unidentified factor for alleviating fatty liver syndrome. Poult. Sci. 1982, 61, 2075–2082. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.S.; Chang, C.H.; Wyatt, R.D. Influence of carbohydrate source on liver fat accumulation in hens. Poult. Sci. 1976, 55, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Maurice, D.V.; Jensen, L.S. Influence of diet composition on hepatic lipid accumulation and hemorrhages in caged layers. Poult. Sci. 1978, 57, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Olomu, J.M.; Robblee, A.R.; Clandinin, D.R.; Hardin, R.T. Effects of span rapeseed meal on productive performance, egg quality, composition of liver and hearts and incidence of “fatty livers” in laying hens. Can. J. Anim. Sci. 1975, 55, 71–75. [Google Scholar] [CrossRef]
- Bhatnagar, M.K.; Yamashiro, S.; David, L.L. Ultrastructural study of liver fibrosis in turkeys fed diets containing rapeseed meal. Res. Vet. Sci. 1980, 29, 260–265. [Google Scholar] [CrossRef]
- Martland, M.F.; Butler, E.J.; Fenwick, G.R. Rapeseed induced liver haemorrhage, reticulolysis and biochemical changes in laying hens: The effects of feeding high and low glucosinolate meals. Res. Vet. Sci. 1984, 36, 298–309. [Google Scholar] [CrossRef]
- Hemsley, L.A.; Edgar, J.; Smetana, P. A comparison of barley, oats, sorghum and wheat in the production of the fatty liver and kidney syndrome. Br. Vet. J. 1973, 129, lxv–lxvi. [Google Scholar] [CrossRef]
- Payne, C.G.; Gilchrist, P.; Pearson, J.A.; Hemsley, L.A. Involvement of biotin in the fatty liver and kidney syndrome of broilers. Br. Poult. Sci. 1974, 15, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Frigg, M. Bioavailability of biotin in cereals. Poult. Sci. 1976, 55, 2310–2318. [Google Scholar] [CrossRef]
- Bryden, W.L.; Cumming, R.B.; Balnave, D. The influence of vitamin A status on the response of chickens to aflatoxin B1 and changes in liver lipid metabolism associated with aflatoxicosis. Br. J. Nutr. 1979, 41, 529–540. [Google Scholar] [CrossRef]
- Jensen, L.S.; Chang, C.H.; Maurice, D.V. Liver lipid accumulation and performance of hens as affected by cage density and initial body weight. Poult. Sci. 1976, 55, 1926–1932. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.J.; Walzem, R.L. Avian fatty liver hemorrhagic syndrome: A comparative review. Adv. Vet. Sci. Comp. Med. 1993, 37, 451–468. [Google Scholar] [PubMed]
- Weitzenburger, D.; Vits, A.; Hamann, H.; Distl, O. Effect of furnished small group housing systems and furnished cages on mortality and causes of death in two layer strains. Br. Poult. Sci. 2005, 46, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Shini, S.; Stewart, G.; Shini, A.; Bryden, W. Mortality rates and causes of death in laying hens kept in cage and alternative housing systems. In Proceedings of the 12th European Poultry Conference, Verona, Italy, 10–14 September 2006; p. 601. [Google Scholar]
- Pearson, A.W.; Butler, E.J. Environmental temperature as a factor in the aetiology of fatty liver-haemorrhagic syndrome in the fowl. Res. Vet. Sci. 1978, 25, 133–138. [Google Scholar] [CrossRef]
- Ivy, C.A.; Nesheim, M.C. Factors influencing the liver fat content of laying hens. Poult. Sci. 1973, 52, 281–291. [Google Scholar] [CrossRef]
- Akiba, Y.; Takahashi, K.; Kimura, M.; Hirama, S.; Matsumoto, T. The influence of environmental temperature, thyroid status and a synthetic oestrogen on the induction of fatty livers in chicks. Br. Poult. Sci. 1983, 24, 71–80. [Google Scholar] [CrossRef]
- Ayo, J.O.; Obidi, J.A.; Rekwot, P.I. Effects of heat stress on the well-being, fertility, and hatchability of chickens in the northern guinea savannah zone of Nigeria: A review. ISRN Vet. Sci. 2011. [Google Scholar] [CrossRef]
- Clark, C.E.; Das, G.P. Effect of high environmental temperature on internal organs of chickens. Poult. Sci. 1974, 53, 859–863. [Google Scholar] [CrossRef]
- Lee, K.; Flegal, C.J.; Wolford, J.H. Factors affecting liver fat accumulation and liver hemorrhages associated with fatty liver-hemorrhagic syndrome in laying chickens. Poult. Sci. 1975, 54, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Pearson, A.W.; Butler, E.J. The oestrogenised chick as an experimental model for fatty liver-haemorrhagic syndrome in the fowl. Res. Vet. Sci. 1978, 24, 82–86. [Google Scholar] [CrossRef]
- Walzem, R.L.; Hansen, R.J.; Williams, D.L.; Hamilton, R.L. Estrogen induction of VLDLy assembly in egg-laying hens. J. Nutr. 1999, 129, 467s–472s. [Google Scholar] [CrossRef] [PubMed]
- Scheele, C.W. Pathological changes in metabolism of poultry related to increasing production levels. Vet. Q. 1997, 19, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Polin, D.; Wolford, J.H. Role of oestrogen as a cause of fatty liver hemorrhagic syndrome. J. Nutr. 1977, 107, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Couch, J.R. Fatty livers in laying hens—A condition which may occur as a result of increased strain. Feedstuffs 1956, 28, 46–54. [Google Scholar]
- Stake, P.E.; Fredrickson, T.N.; Bourdeau, C.A. Induction of fatty liver-hemorrhagic syndrome in laying hens by exogenous beta-estradiol. Avian Dis. 1981, 25, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Abplanalp, H.; Napolitano, D. Genetic predisposition for fatty liver rupture in White Leghorn hens of a highly inbred line. Poult. Sci. 1988, 67 (Suppl. l), 52. [Google Scholar]
- Squires, E.J.; Leeson, S. Aetiology of fatty liver syndrome in laying hens. Br. Vet. J. 1988, 144, 602–609. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fat Source | Weight (g/kg BW) | DM (g/kg) | Protein (g/kg) | Ash (g/kg) | Fat (g/kg) |
|---|---|---|---|---|---|
| Soybean oil | 26.9 | 241 | 690 | 59 | 125 |
| Tallow | 26.3 | 234 | 688 | 60 | 130 |
| Bile Acid | Chicken | Duck |
|---|---|---|
| Cholic acid | 9.6 ± 0.5 | 45.2 ± 2.3 |
| Chenodeoxycholic acid | 25.2 ± 2.2 | 28.2 ± 1.6 |
| Ursodeoxycholic acid | n.d. | 43.5 ± 2.1 |
| Deoxycholic acid | n.d. | 31.6 ± 1.9 |
| Lithocholic acid | 68.7 ± 2.1 | 37.5 ± 2.1 |
| Taurocholic acid | 152.6 ± 3.1 | 16.8 ± 1.5 |
| Taurochenodeoxycholic acid | n.d. | 97.5 ± 3.4 |
| Taurolithocholic acid | 35.9 ± 0.6 | n.d. |
| Glycolithocholic acid | 228.4 ± 1.6 | n.d. |
| Reference | Age (day) | Grain Type | Whole Grain Inclusion (g/kg) | Weight Gain | Liver Weight |
|---|---|---|---|---|---|
| % Changes | |||||
| [137] | 7–38 | Barley | 100 1, 150 2 | +0.6 | +6.5 |
| 150 1, 150 2 | +2.4 | +8.4 | |||
| [137] | 7–38 | Wheat | 100 1, 200 2 | +4.2 | +7.8 |
| 100 1, 350 2 | +5.1 | +3.7 | |||
| 200 1, 200 2 | +1.3 | +3.7 | |||
| 200 1, 350 2 | +2.0 | +9.3 | |||
| [10] | 1–14 | Wheat | 100 | −9.7 | +11.9 |
| [140] | 21–42 | Wheat | 300 | −3.8 | +8.5 |
| [138] | 11–35 | Maize | 150 | +3.3 | +1.3 |
| 300 | +7.0 | +16.2 | |||
| 450 | +12.0 | +8.3 | |||
| 600 | +8.7 | +5.7 | |||
| [141] | 1–10 | Sorghum | 250 | +8.7 | +5.7 |
| 500 | +4.4 | −3.4 | |||
| 750 | +5.6 | +8.4 | |||
| [141] | 1–35 | Sorghum | 250 | −0.87 | +5.3 |
| 500 | −6.5 | +5.7 | |||
| 750 | −5.2 | +6.0 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaefarian, F.; Abdollahi, M.R.; Cowieson, A.; Ravindran, V. Avian Liver: The Forgotten Organ. Animals 2019, 9, 63. https://doi.org/10.3390/ani9020063
Zaefarian F, Abdollahi MR, Cowieson A, Ravindran V. Avian Liver: The Forgotten Organ. Animals. 2019; 9(2):63. https://doi.org/10.3390/ani9020063
Chicago/Turabian StyleZaefarian, Faegheh, Mohammad Reza Abdollahi, Aaron Cowieson, and Velmurugu Ravindran. 2019. "Avian Liver: The Forgotten Organ" Animals 9, no. 2: 63. https://doi.org/10.3390/ani9020063
APA StyleZaefarian, F., Abdollahi, M. R., Cowieson, A., & Ravindran, V. (2019). Avian Liver: The Forgotten Organ. Animals, 9(2), 63. https://doi.org/10.3390/ani9020063