Salivary Vasopressin as a Potential Non–Invasive Biomarker of Anxiety in Dogs Diagnosed with Separation–Related Problems

 , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Study Setting
2.2. Study Protocol
2.3. Parameters Recorded: Behavioral Responses
2.4. Parameters Recorded: Endocrine Responses
2.5. Statistical Analysis
3. Results
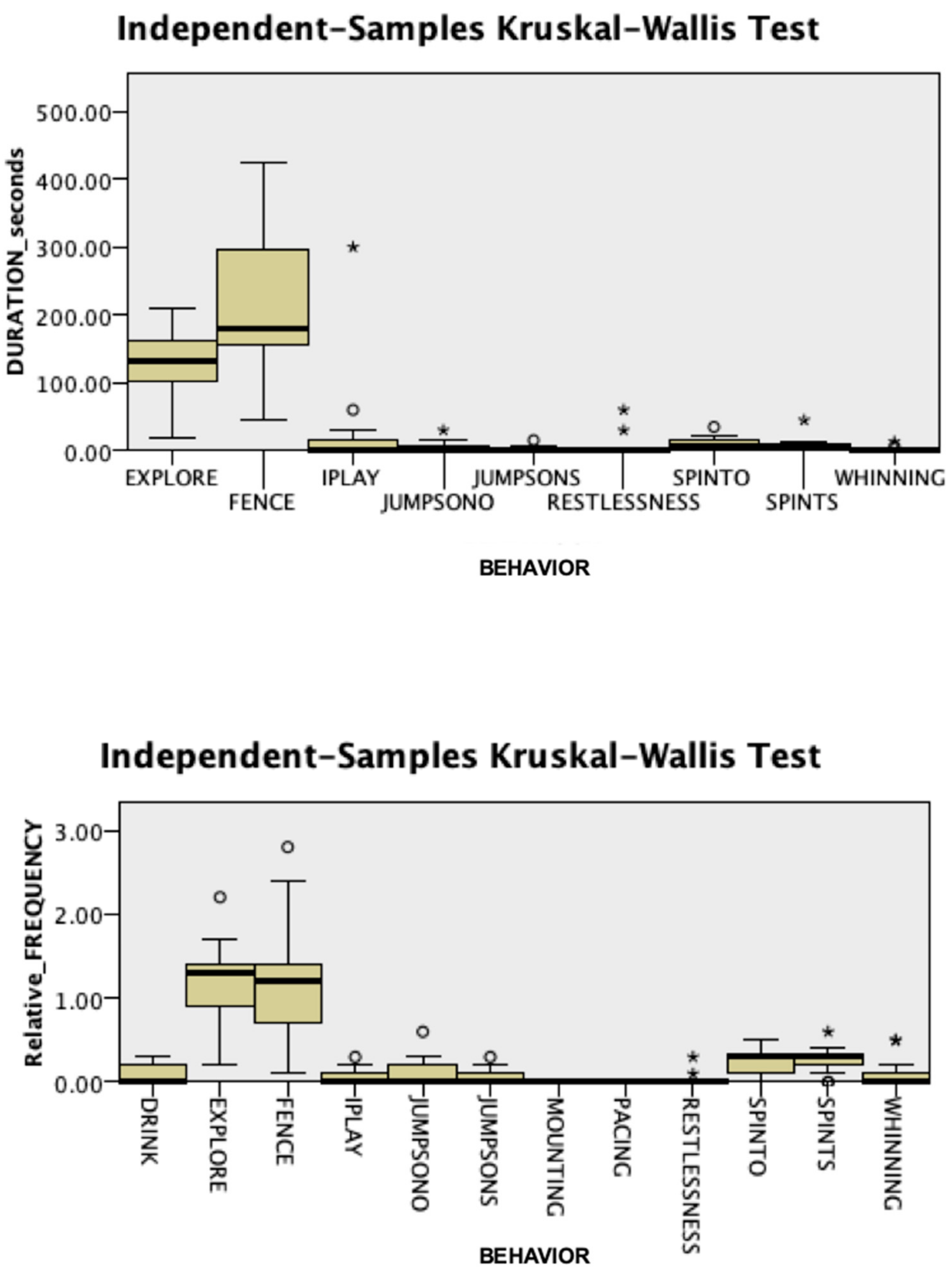
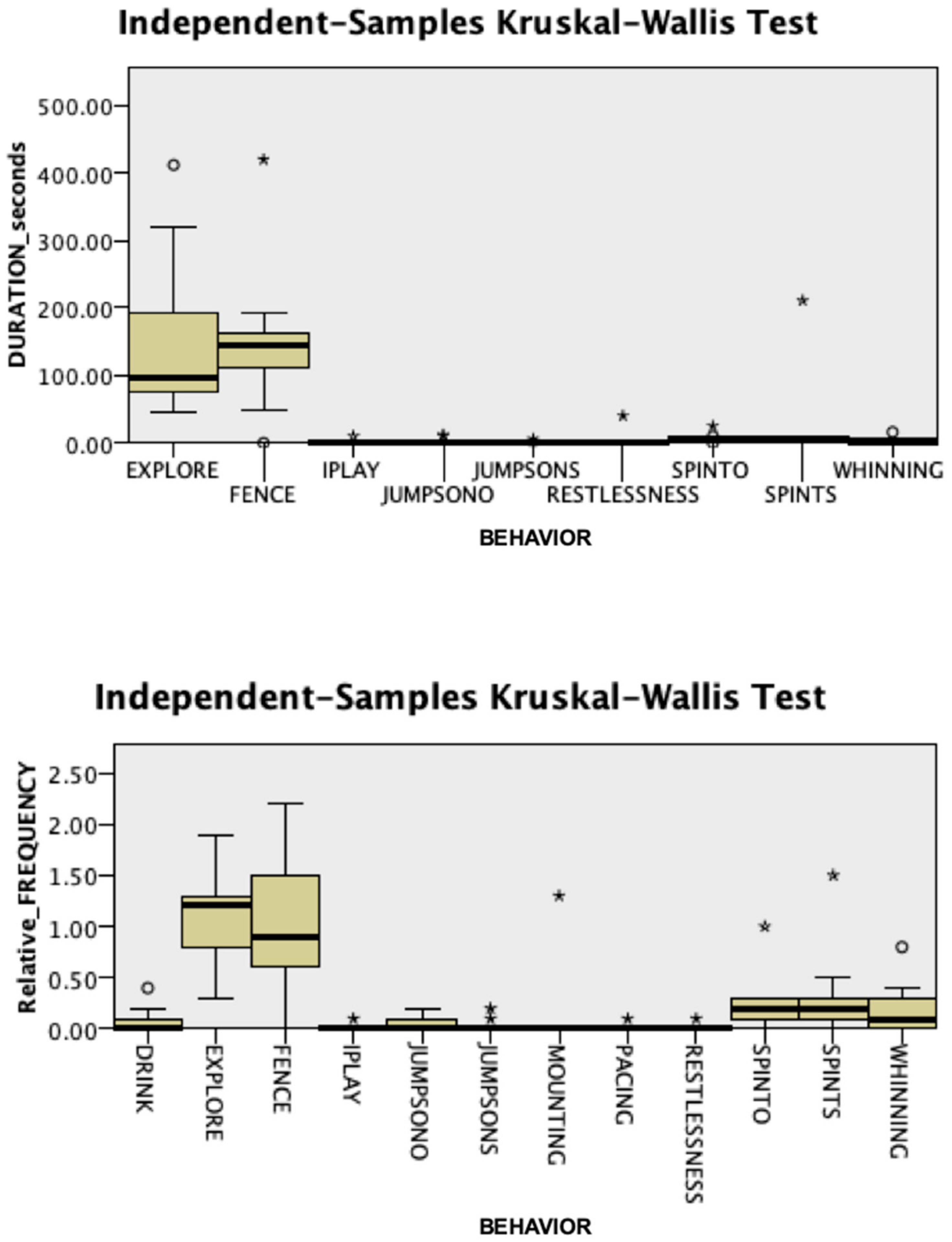
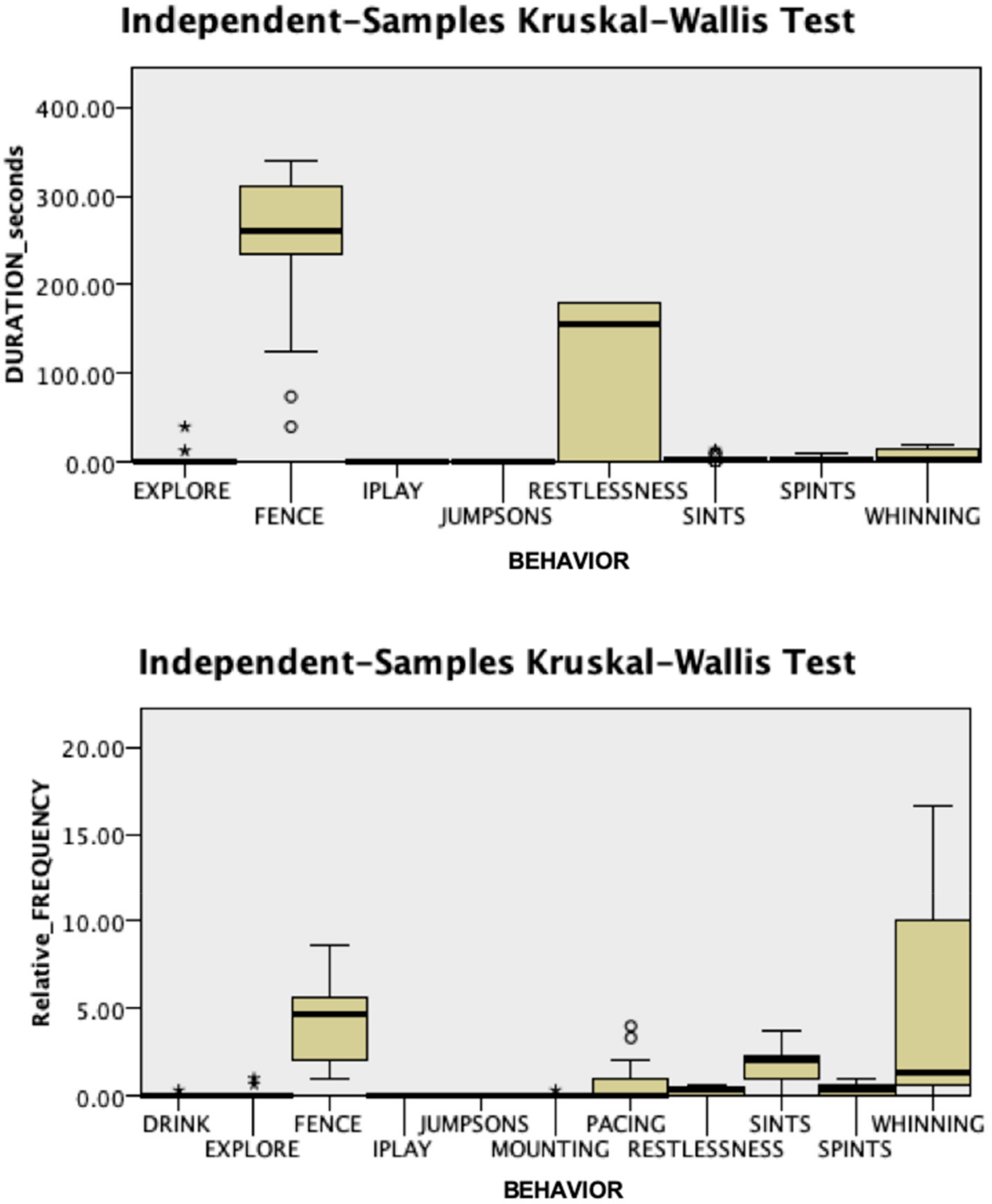
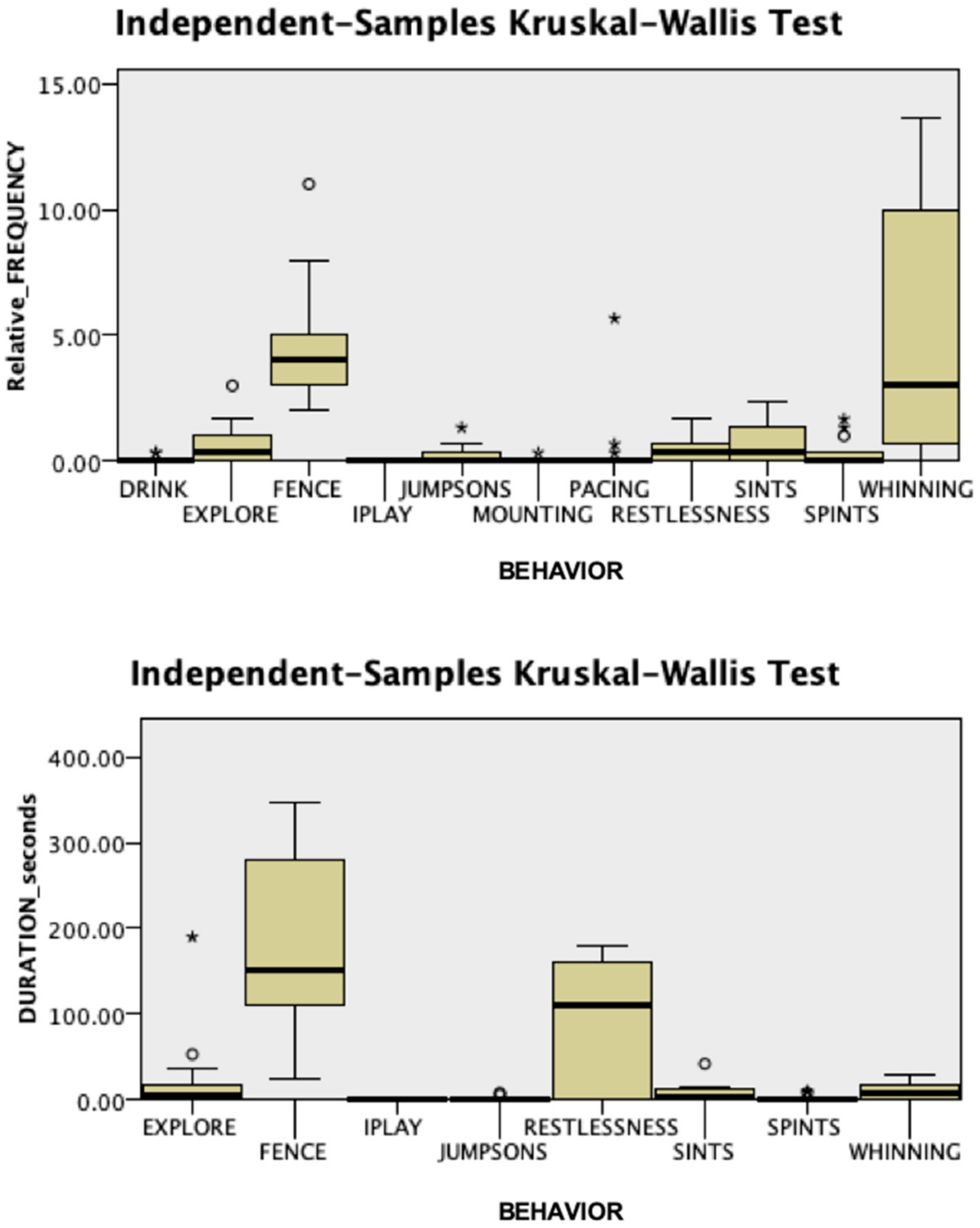
3.1. Behavioral Responses
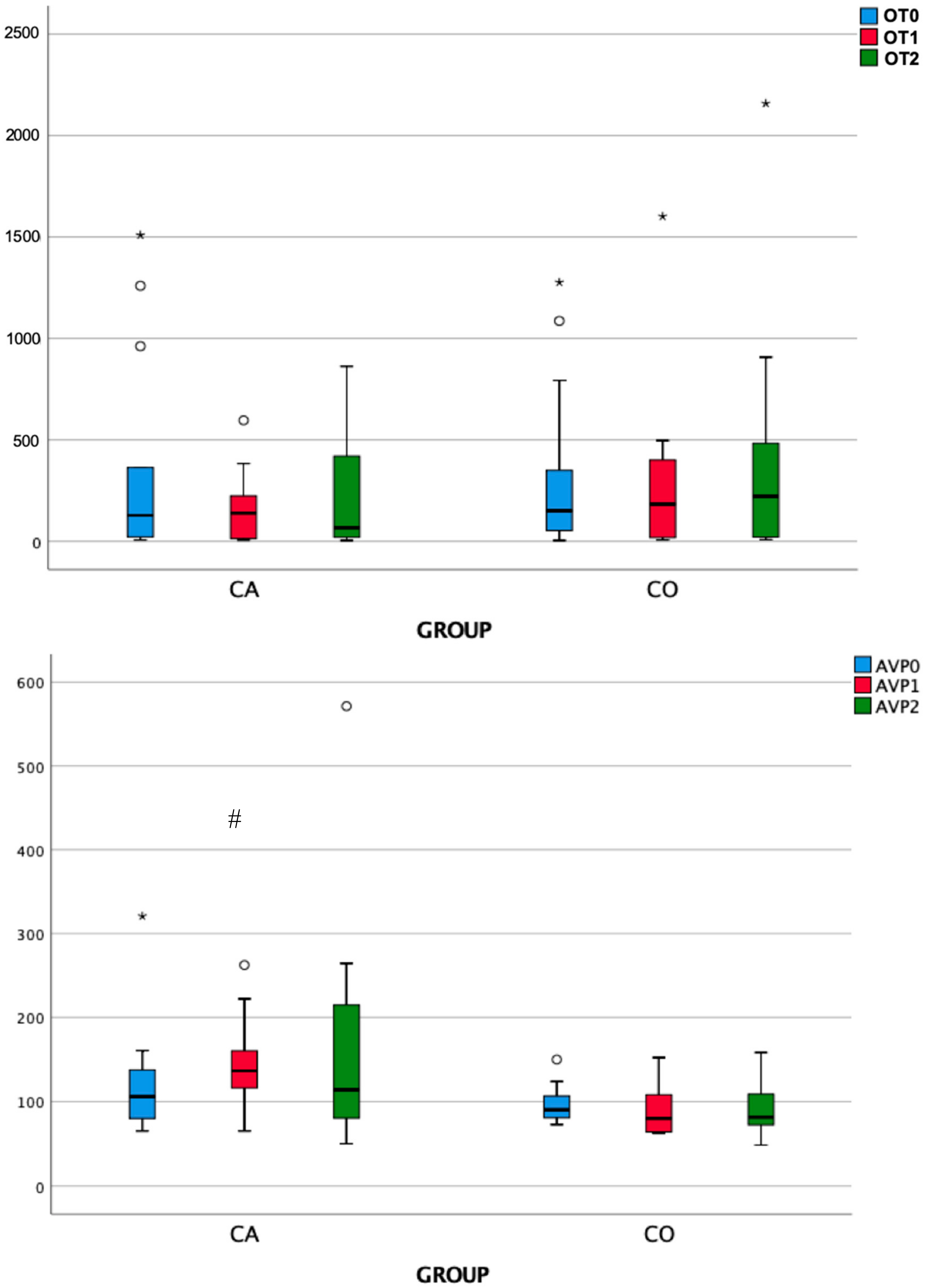
3.2. Endocrine Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carter, C.S. The Role of Oxytocin and Vasopressin in Attachment. Psychodyn. Psych. 2017, 45, 499–517. [Google Scholar] [CrossRef] [PubMed]
- Mariti, C.; Ricci, E.; Carlone, B.; Moore, J.L.; Sighieri, C.; Gazzano, A. Dog attachment to man: A comparison between pet and working dogs. J. Vet. Behav. 2013, 8, 135–145. [Google Scholar] [CrossRef]
- Rehn, T.; Handlin, L.; Uvnäs–Moberg, K.; Keeling, L.J. Dogs’ endocrine and behavioural responses at reunion are affected by how the human initiates contact. Physiol. Behav. 2014, 124, 45–53. [Google Scholar] [CrossRef]
- Parthasarathy, V.; Crowell–Davis, S.L. Relationship between attachment to owners and separation anxiety in pet dogs (Canis lupus familiaris). J. Vet. Behav. 2006, 1, 109–120. [Google Scholar] [CrossRef]
- Konok, V.; Dóka, A.; Miklósi, Á. The behavior of the domestic dog (Canis familiaris) during separation from and reunion with the owner: A questionnaire and an experimental study. Appl. Anim. Behav. Sci. 2011, 135, 300–308. [Google Scholar] [CrossRef]
- Palestrini, C.; Previde, E.P.; Spiezio, C.; Verga, M. Heart rate and behavioural responses of dogs in the Ainsworth’s Strange Situation: A pilot study. Appl. Anim. Behav. Sci. 2005, 94, 75–88. [Google Scholar] [CrossRef]
- Aloe, L.; Bracci–Laudiero, L.; Allevat, E.; Lambiase, A.; Micera, A.; Tirassa, P. Emotional stress induced by parachute jumping enhances blood nerve growth factor levels and the distribution of nerve growth factor receptors in lymphocytes (str/nxet/hormone). Proc. Natl. Acad. Sci. USA 1994, 91, 10440–10444. [Google Scholar] [CrossRef]
- Aloe, L.; Allevat, E.; Bohm, A.; Levi–Montalcini, R. Aggressive behavior induces release of nerve growth factor from mouse salivary gland into the bloodstream (submaxillary salivary gland/adrenal gland). Proc. Natl. Acad. Sci. USA 1986, 83, 6184–6187. [Google Scholar] [CrossRef]
- Campbell, T.; Lin, S.; DeVries, C.; Lambert, K. Coping strategies in male and female rats exposed to multiple stressors. Physiol. Behav. 2003, 78, 495–504. [Google Scholar] [CrossRef]
- Neumann, I.D.; Landgraf, R. Balance of brain oxytocin and vasopressin: Implications for anxiety, depression, and social behaviors. Trends Neurosci. 2012, 35, 649–659. [Google Scholar] [CrossRef]
- Uvnäs–Moberg, K. Oxytocin may mediate the benefits of positive social interaction and emotions. Psychoneuroendocrinology 1998, 23, 819–835. [Google Scholar] [CrossRef]
- Wotjak, C.; Kubota, M.; Liebsch, G.; Montkowski, A.; Holsboer, F.; Neumann, I.; Landgraf, R. Release of vasopressin within the rat paraventricular nucleus in response to emotional stress: A novel mechanism of regulating adrenocorticotropic hormone secretion? Soc. Neurosci. 1996, 16, 7725–7732. [Google Scholar] [CrossRef]
- MacLean, E.L.; Gesquiere, L.R.; Gee, N.R.; Levy, K.; Martin, W.L.; Carter, C.S. Effects of Affiliative Human–Animal Interaction on Dog Salivary and Plasma Oxytocin and Vasopressin. Front. Psychol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, M.; Mitsui, S.; En, S.; Ohtani, N.; Ohta, M.; Sakuma, Y.; Onaka, T.; Mogi, K.; Kikusui, T. Oxytocin–gaze positive loop and the coevolution of human–dog bonds. Science 2015, 348, 333–336. [Google Scholar] [CrossRef]
- Hydbring–Sandberg, E.; von Walter, L.; Höglund, K.; Svartberg, K.; Forkman, L.B.S. Physiological reactions to fear provocation in dogs. J. Endocrinol. 2004, 180, 439–448. [Google Scholar] [CrossRef]
- MacLean, E.L.; Gesquiere, L.R.; Gruen, M.E.; Sherman, B.L.; Martin, W.L.; Carter, C.S. Endogenous Oxytocin, Vasopressin, and Aggression in Domestic Dogs. Front. Psychol. 2017, 8, 1613. [Google Scholar] [CrossRef]
- Overall, K.L. Manual of Clinical Behavioral Medicine for Dogs and Cats; Elsevier Saunders: Philadelphia, PA, USA, 2013; ISBN 9780323240659. [Google Scholar]
- Guthrie, A. Dogs behaving badly–Canine separation disorder research. Vet. Pr. 1999, 31, 12–13. [Google Scholar]
- Landsberg, G.; Hunthausen, W.; Ackerman, L. Fear and Phobias, Separation Anxiety. In Handbook of Behavior Problems of the Dog and Cat, 2nd ed.; Landsberg, G.M., Hunthausen, W.L., Ackerman, L., Eds.; Saunders Company: Philadelphia, PA, USA, 2003; pp. 258–267. [Google Scholar]
- Blackwell, E.; Casey, R.; Bradshaw, J. Controlled trial of behavioural therapy for separation–related disorders in dogs. Vet. Rec. 2006, 158, 551–554. [Google Scholar] [CrossRef]
- Overall, K.L.; Dunham, A.E.; Juarbe–Diaz, S.V. Phenotypic determination of noise reactivity in 3 breeds of working dogs: A cautionary tale of age, breed, behavioral assessment, and genetics. J. Vet. Behav. 2016, 16, 113–125. [Google Scholar] [CrossRef]
- Tami, G.A.G. Description of the behaviour of domestic dog (Canis familiaris) by experienced and inexperienced people. Appl. Anim. Behav. 2009, 120, 159–169. [Google Scholar] [CrossRef]
- Mariti, C.; Gazzano, A.; Moore, J.; Baragli, P. Perception of dogs’ stress by their owners. J. Vet. Behav. Clin. Appl. Res. 2012, 7, 213–219. [Google Scholar] [CrossRef]
- Karagiannis, C.I.; Burman, O.H.; Mills, D.S. Dogs with separation–related problems show a “less pessimistic” cognitive bias during treatment with fluoxetine (ReconcileTM) and a behaviour modification plan. BMC Vet. Res. 2015, 11, 80. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.; McPherson, J.; Casey, R.; Larter, I. Aetiology of separation–related behaviour in domestic dogs. Vet. Rec. 2002, 151, 43–46. [Google Scholar] [CrossRef]
- Soares, G.; Pereira, J.; Paixão, R. Exploratory study of separation anxiety syndrome in apartment dogs. Cienc. Rural. 2010, 40, 548–553. [Google Scholar] [CrossRef]
- Statista Number of dogs in the United States from 2000 to 2017 (in millions). Available online: https://www.statista.com/statistics/198100/dogs–in–the–united–states–since–2000/ (accessed on 15 July 2019).
- Statista Number of dogs in the European Union in 2017, by country (in 1000s). Available online: https://www.statista.com/statistics/414956/dog–population–european–union–eu–by–country/ (accessed on 15 July 2019).
- Diesel, G.; Brodbelt, D.; Pfeiffer, D.U. Characteristics of Relinquished Dogs and Their Owners at 14 Rehoming Centers in the United Kingdom. J. Appl. Anim. Welf. Sci. 2010, 13, 15–30. [Google Scholar] [CrossRef]
- New, J.C., Jr.; Salman, M.D.; Scarlett, J.M.; Kass, P.H.; Vaughn, J.A.; Scherr, S.; Kelch, W.J. Moving: Characteristics of Dogs and Cats and Those Relinquishing Them to 12 U.S. Animal Shelters. J. Appl. Anim. Welf. Sci. 1999, 2, 83–96. [Google Scholar] [CrossRef]
- MacLean, E.; Gesquiere, L.; Gee, N.; Levy, K. Validation of salivary oxytocin and vasopressin as biomarkers in domestic dogs. J. Neurosci. 2017, 293, 67–76. [Google Scholar] [CrossRef]
- O’Farrell, V. Owner attitudes and dog behaviour problems. Appl. Anim. Behav. Sci. 1997, 52, 205–213. [Google Scholar] [CrossRef]
- Pastore, C.; Pirrone, F.; Balzarotti, F.; Faustini, M.; Pierantoni, L.; Albertini, M. Evaluation of physiological and behavioral stress–dependent parameters in agility dogs. J. Vet. Behav. Clin. Appl. Res. 2011, 6, 188–194. [Google Scholar] [CrossRef]
- Pirrone, F.; Ripamonti, A.; Garoni, E.C.; Stradiotti, S.; Albertini, M. Measuring social synchrony and stress in the handler–dog dyad during animal–assisted activities: A pilot study. J. Vet. Behav. Clin. Appl. Res. 2017, 21, 45–52. [Google Scholar] [CrossRef]
- Prato–Previde, E.; Spiezio, C.; Sabatini, F.; Custance, D.M. Is the dog–human relationship an attachment bond? An observational study using Ainsworth’s strange situation. Behaviour 2003, 140, 225–254. [Google Scholar] [CrossRef]
- Scaglia, E. Video analysis of adult dogs when left home alone. J. Vet. Behav. Clin. Appl. Res. 2013, 8, 412–417. [Google Scholar] [CrossRef]
- Topál, J.; Gácsi, M.; Miklósi, Á.; Virányi, Z.; Kubinyi, E.; Csányi, V. Attachment to humans: a comparative study on hand–reared wolves and differently socialized dog puppies. Anim. Behav. 2005, 70, 1367–1375. [Google Scholar] [CrossRef]
- Choi, S.W. Life is lognormal! What to do when your data does not follow a normal distribution. Anaesthesia 2016, 71, 1363–1366. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–120. [Google Scholar] [CrossRef]
- Flannigan, G.; Dodman, N.H. Risk factors and behaviors associated with separation anxiety in dogs. J. Am. Vet. Med. Assoc. 2001, 219, 460–466. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Houpt, K.A.; Scarlett, J.M. Evaluation of treatments for separation anxiety in dogs. J. Am. Vet. Med. Assoc. 2000, 217, 342–345. [Google Scholar] [CrossRef]
- Ainsworth, M.D.S. Individual Differences in Strange–Situational Behaviour of One–Year–Olds. In Determinants of Infant Behavior (Vol. 4); Foss, B.M., Ed.; Methuen: London, UK, 1969; pp. 111–136. [Google Scholar]
- Bard, K.A.; Nadler, R.D. The effect of peer separation in young chimpanzees (Pan troglodytes). Am. J. Primatol. 1983, 5, 25–37. [Google Scholar] [CrossRef]
- Kaufman, I.C.; Rosenblum, L.A. Effects of separation from mother on the emotional behavior of infant monkeys. Ann. N.Y. Acad. Sci. 1969, 159, 681–695. [Google Scholar] [CrossRef]
- Topál, J.; Miklósi, Á.; Csányi, V.; Dóka, A. Attachment Behavior in Dogs (Canis familiaris): A New Application of Ainsworth’s (1969) Strange Situation Test. J. Comp. Psychol. 1998, 112, 219–229. [Google Scholar] [CrossRef]
- White–Traut, R.; Watanabe, K.; Pournajafi–Nazarloo, H.; Schwertz, D.; Bell, A.; Carter, C.S. Detection of salivary oxytocin levels in lactating women. Dev. Psychobiol. 2009, 51, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Tiira, K.; Lohi, H. Early Life Experiences and Exercise Associate with Canine Anxieties. PLoS ONE 2015, 10, e0141907. [Google Scholar] [CrossRef]
- Ludwig, M.; Leng, G. Dendritic peptide release and peptide–dependent behaviours. Nat. Rev. Neurosci. 2006, 7, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, E.L.; Caldwell, H.K. The vasopressin 1b receptor and the neural regulation of social behavior. Horm. Behav. 2012, 61, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Baribeau, D.A.; Anagnostou, E. Oxytocin and vasopressin: Linking pituitary neuropeptides and their receptors to social neurocircuits. Front. Neurosci. 2015, 9. [Google Scholar] [CrossRef]
- Pow, D.; Morris, J. Dendrites of hypothalamic magnocellular neurons release neurohypophysial peptides by exocytosis. Neuroscience 1989, 32, 435–439. [Google Scholar] [CrossRef]
- Knobloch, H.S.; Charlet, A.; Hoffmann, L.C.; Eliava, M.; Khrulev, S.; Cetin, A.H.; Osten, P.; Schwarz, M.K.; Seeburg, P.H.; Stoop, R.; et al. Evoked Axonal Oxytocin Release in the Central Amygdala Attenuates Fear Response. Neuron 2012, 73, 553–566. [Google Scholar] [CrossRef]
- Neumann, R. Involvement of the brain oxytocin system in stress coping: Interactions with the hypothalamo–pituitary–adrenal axis. Trends Neurosci. 2002, 35, 649–659. [Google Scholar] [CrossRef]
- Weisman, O.; Schneiderman, I.; Zagoory–Sharon, O.; Feldman, R. Salivary vasopressin increases following intranasal oxytocin administration. Peptides 2013, 40, 99–103. [Google Scholar] [CrossRef]
- Carson, D. Plasma vasopressin concentrations positively predict cerebrospinal fluid vasopressin concentrations in human neonates. Peptides 2014, 61, 12–16. [Google Scholar] [CrossRef]
- Griebel, G.; Beeské, S.; Stahl, S.M. The Vasopressin V 1b Receptor Antagonist SSR149415 in the Treatment of Major Depressive and Generalized Anxiety Disorders: Results From 4 Randomized, Double–Blind, Placebo–Controlled Studies. J. Clin. Psychiatry 2012, 73, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Iijima, M.; Yoshimizu, T.; Shimazaki, T.; Tokugawa, K.; Fukumoto, K.; Kurosu, S.; Kuwada, T.; Sekiguchi, Y.; Chaki, S. Antidepressant and anxiolytic profiles of newly synthesized arginine vasopressin V 1B receptor antagonists: TASP0233278 and TASP0390325. Br. J. Pharmacol. 2014, 171, 3511–3525. [Google Scholar] [CrossRef] [PubMed]
- Cannas, S.; Frank, D.; Minero, M.; Aspesi, A.; Benedetti, R.; Palestrini, C. Video analysis of dogs suffering from anxiety when left home alone and treated with clomipramine. J. Vet. Behav. Clin. Appl. Res. 2014, 9, 50–57. [Google Scholar] [CrossRef]
- Heinrichs, M.; von Dawans, B.; Domes, G. Oxytocin, vasopressin, and human social behavior. Front. Neuroendocr. 2009, 30, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hackett, P.D.; DeMarco, A.C.; Feng, C.; Stair, S.; Haroon, E.; Ditzen, B.; Pagnoni, G.; Rilling, J.K. Effects of oxytocin and vasopressin on the neural response to unreciprocated cooperation within brain regions involved in stress and anxiety in men and women. Brain Imaging Behav. 2016, 10, 581–593. [Google Scholar] [CrossRef]
- Thielke, L.E.; Udell, M.A.R. The role of oxytocin in relationships between dogs and humans and potential applications for the treatment of separation anxiety in dogs. Biol. Rev. 2017, 92, 378–388. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behaviors | Description | Measured Values (F/D) |
|---|---|---|
| Social behaviors | ||
| Jumping up | Both of the dog’s forelegs were out of contact with the ground, regardless of the position of the hind legs; the dog was in proximity to a person. The dog might also be entirely on a person’s lap | F, D |
| Spontaneous interactions | Staying close to and seeking attention and physical contact (nuzzling or pawing for attention, soliciting petting) from the owner or the stranger | F, D |
| Mounting | Sexual mounting of people or inanimate objects | F, D |
| Non–social behaviors | ||
| Explore | Activity directed towards physical aspects of the environment, including sniffing, visual inspection, and gentle licking | F, D |
| Individual play | Any behavior performed vigorously or at a galloping gait and directed towards an object when clearly not interacting with any human; these play behaviors included chewing, biting, shaking from side to side, scratching or batting with the paw, chasing rolling balls, and tossing objects using the mouth | F, D |
| Standing by the fence | Standing close to the fence (<1 m), regardless of whether the face was oriented towards the exit | F, D |
| Attention oriented towards the fence | Staring fixedly at the fence, either when close to it or from a distance | F, D |
| Behaviors oriented towards the fence | All activities, resulting in physical contact with the fence, including scratching the gate with the paws, jumping on the fence, and pulling on the fence with the forelegs or mouth. | F, D |
| Restlessness | A feeling of agitation expressed by continual motion; changing the state of locomotion; digging (scratching the floor with the forepaws in a way that is similar to when dogs are digging holes) | F |
| Drinking | Taking in fluids by lapping up water from the bowl with the tongue | F |
| Whining | High-pitched vocalization | F |
| Pacing | Increased motor activity, walking or running around without exploring the environment | F |
| Group | OT0 | OT1 | OT2 | Friedman test | |
| χ2 | P | ||||
| Case | 127.87 | 138.79 | 67.04 | 3.231 | 0.199 |
| Control | 149.99 | 183 | 221.60 | 0.462 | 0.794 |
| Mann–Whitney U test | 86 | 98 | 100 | ||
| Mann–Whitney U test p | 0.960 | 0.511 | 0.448 | ||
| Group | AVP0 | AVP1 | AVP2 | Friedman test | |
| χ2 | P | ||||
| Case | 105.97 | 136.61 | 114 | 2.923 | 0.232 |
| Control | 90.40 | 80.12 | 81.53 | 3.231 | 0.199 |
| Mann–Whitney U test | 71 | 29 | 55 | ||
| Mann–Whitney U test p | 0.511 | 0.003 | 0.139 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pirrone, F.; Pierantoni, L.; Bossetti, A.; Uccheddu, S.; Albertini, M. Salivary Vasopressin as a Potential Non–Invasive Biomarker of Anxiety in Dogs Diagnosed with Separation–Related Problems. Animals 2019, 9, 1033. https://doi.org/10.3390/ani9121033
Pirrone F, Pierantoni L, Bossetti A, Uccheddu S, Albertini M. Salivary Vasopressin as a Potential Non–Invasive Biomarker of Anxiety in Dogs Diagnosed with Separation–Related Problems. Animals. 2019; 9(12):1033. https://doi.org/10.3390/ani9121033
Chicago/Turabian StylePirrone, Federica, Ludovica Pierantoni, Andrea Bossetti, Stefania Uccheddu, and Mariangela Albertini. 2019. "Salivary Vasopressin as a Potential Non–Invasive Biomarker of Anxiety in Dogs Diagnosed with Separation–Related Problems" Animals 9, no. 12: 1033. https://doi.org/10.3390/ani9121033
APA StylePirrone, F., Pierantoni, L., Bossetti, A., Uccheddu, S., & Albertini, M. (2019). Salivary Vasopressin as a Potential Non–Invasive Biomarker of Anxiety in Dogs Diagnosed with Separation–Related Problems. Animals, 9(12), 1033. https://doi.org/10.3390/ani9121033