Parasite Load and Site-Specific Parasite Pressure as Determinants of Immune Indices in Two Sympatric Rodent Species

 , , , and
, , , and
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Rodent Trapping
2.3. Rodent Anesthetization, Sample Collection and Handling
2.4. Ectoparasite Screening
2.5. Gastrointestinal Parasite Screening
2.6. Screening for Microparasites
2.7. Immunological Assays
2.7.1. Haptoglobin
2.7.2. Hemolysis and Hemagglutination
2.7.3. Neutrophil to Lymphocyte Ratio
2.8. Data Analysis
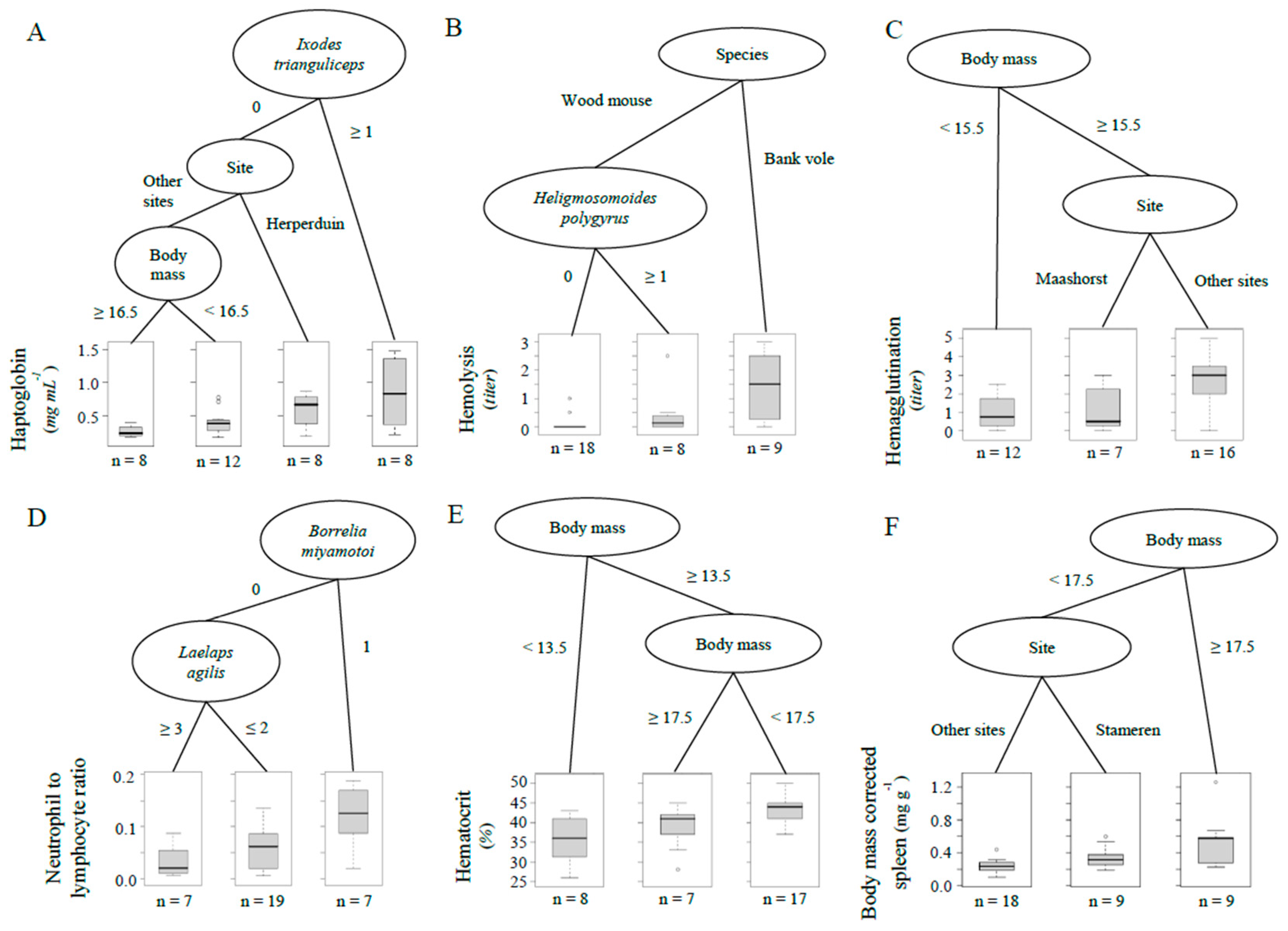
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Horrocks, N.P.C.; Matson, K.D.; Tieleman, B.I. Pathogen Pressure Puts Immune Defense into Perspective. Integr. Comp. Biol. 2011, 51, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Viney, M.; Riley, E.M. The Immunology of Wild Rodents: Current Status and Future Prospects. Front. Immunol. 2017, 8, 1481. [Google Scholar] [CrossRef] [PubMed]
- Råberg, L.; Graham, A.L.; Read, A.F. Decomposing health: Tolerance and resistance to parasites in animals. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Baucom, R.S.; de Roode, J.C. Ecological immunology and tolerance in plants and animals. Funct. Ecol. 2011, 25, 18–28. [Google Scholar] [CrossRef]
- Martin, L.B.; Hawley, D.M.; Ardia, D.R. An introduction to ecological immunology. Funct. Ecol. 2011, 25, 1–4. [Google Scholar] [CrossRef]
- Kurtenbach, K.; Dizij, A.; Seitz, H.M.; Margos, G.; Moter, S.E.; Kramer, M.D.; Wallich, R.; Schaible, U.E.; Simon, M.M. Differential immune responses to Borrelia burgdorferi in European wild rodent species influence spirochete transmission to Ixodes ricinus L. (Acari: Ixodidae). Infect. Immun. 1994, 62, 5344–5352. [Google Scholar]
- Matson, K.D.; Cohen, A.A.; Klasing, K.C.; Ricklefs, R.E.; Scheuerlein, A. No simple answers for ecological immunology: Relationships among immune indices at the individual level break down at the species level in waterfowl. Proc. R. Soc. B-Biol. Sci. 2006, 273, 815–822. [Google Scholar] [CrossRef]
- Previtali, M.A.; Ostfeld, R.S.; Keesing, F.; Jolles, A.E.; Hanselmann, R.; Martin, L.B. Relationship between pace of life and immune responses in wild rodents. Oikos 2012, 121, 1483–1492. [Google Scholar] [CrossRef]
- Matson, K.D.; Ricklefs, R.E.; Klasing, K.C. A hemolysis-hemagglutination assay for characterizing constitutive innate humoral immunity in wild and domestic birds. Dev. Comp. Immunol. 2005, 29, 275–286. [Google Scholar] [CrossRef]
- Matson, K.D.; Horrocks, N.P.C.; Versteegh, M.A.; Tieleman, B.I. Baseline haptoglobin concentrations are repeatable and predictive of certain aspects of a subsequent experimentally-induced inflammatory response. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 162, 7–15. [Google Scholar] [CrossRef]
- Moyer, B.R.; Drown, D.M.; Clayton, D.H. Low humidity reduces ectoparasite pressure: Implications for host life history evolution. Oikos 2002, 97, 223–228. [Google Scholar] [CrossRef]
- Rynkiewicz, E.C.; Hawlena, H.; Durden, L.A.; Hastriter, M.W.; Demas, G.E.; Clay, K. Associations between innate immune function and ectoparasites in wild rodent hosts. Parasitol. Res. 2013, 112, 1763–1770. [Google Scholar] [CrossRef]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging Infectious Diseases of Wildlife - Threats to Biodiversity and Human Health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Hofmeester, T.R.; Jansen, P.A.; Wijnen, H.J.; Coipan, E.C.; Fonville, M.; Prins, H.H.T.; Sprong, H.; van Wieren, S.E. Cascading effects of predator activity on tick-borne disease risk. Proc. R. Soc. B 2017, 284. [Google Scholar] [CrossRef] [PubMed]
- Dizij, A.; Kurtenbach, K. Clethrionomys glareolus, but not Apodemus flavicollis, acquires resistance to Ixodes ricinus L., the main European vector of Borrelia burgdorferi. Parasite Immunol. 1995, 17, 177–183. [Google Scholar] [CrossRef]
- Telfer, S.; Lambin, X.; Birtles, R.; Beldomenico, P.; Burthe, S.; Paterson, S.; Begon, M. Species interactions in a parasite community drive infection risk in a wildlife population. Science 2010, 330, 243–246. [Google Scholar] [CrossRef]
- Campbell, M.T.; Dobson, F.S. Growth and Size in Meadow Voles (Microtus pennsylvanicus). Am. Midl. Nat. 1992, 128, 180–190. [Google Scholar] [CrossRef]
- Powell, R.A.; Proulx, G. Trapping and marking terrestrial mammals for research: Integrating ethics, performance criteria, techniques, and common sense. ILAR J. 2003, 44, 259–276. [Google Scholar] [CrossRef]
- Hillyard, P.D. Ticks of North-West Europe; Backhuys Publishers: London, UK, 1996. [Google Scholar]
- Whitaker, A.P. Handbooks for the Identification of British Insects: Fleas (Siphonaptera), 2nd ed.; Royal Entomological Society: London, UK, 2007. [Google Scholar]
- Franssen, F.; Swart, A.; van Knapen, F.; van der Giessen, J. Helminth parasites in black rats (Rattus rattus) and brown rats (Rattus norvegicus) from different environments in the Netherlands. Infect. Ecol. Epidemiol. 2016, 6, 31413. [Google Scholar] [CrossRef]
- Heylen, D.; Matthysen, E.; Fonville, M.; Sprong, H. Songbirds as general transmitters but selective amplifiers of Borrelia burgdorferi sensu lato genotypes in Ixodes rinicus ticks. Environ. Microbiol. 2014, 16, 2859–2868. [Google Scholar] [CrossRef] [PubMed]
- Hovius, J.W.R.; De Wever, B.; Sohne, M.; Brouwer, M.C.; Coumou, J.; Wagemakers, A.; Oei, A.; Knol, H.; Narasimhan, S.; Hodiamont, C.J.; et al. A case of meningoencephalitis by the relapsing fever spirochaete Borrelia miyamotoi in Europe. Lancet 2013, 382, 658. [Google Scholar] [CrossRef]
- Courtney, J.W.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.F. Multiplex real-time PCR for detection of Anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef] [PubMed]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Földvári, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasit. Vectors 2014, 7, 365. [Google Scholar] [CrossRef]
- Jahfari, S.; Fonville, M.; Hengeveld, P.; Reusken, C.; Scholte, E.J.; Takken, W.; Heyman, P.; Medlock, J.M.; Heylen, D.; Kleve, J.; et al. Prevalence of Neoehrlichia mikurensis in ticks and rodents from North-west Europe. Parasit. Vectors 2012, 5, 74. [Google Scholar] [CrossRef]
- Stenos, J.; Graves, S.R.; Unsworth, N.B. A highly sensitive and specific real-time PCR assay for the detection of spotted fever and typhus group Rickettsiae. Am. J. Trop. Med. Hyg. 2005, 73, 1083–1085. [Google Scholar] [CrossRef]
- Ahmed, A.; Engelberts, M.F.M.; Boer, K.R.; Ahmed, N.; Hartskeerl, R.A. Development and validation of a real-time PCR for detection of pathogenic leptospira species in clinical materials. PLoS ONE 2009, 4, e7093. [Google Scholar] [CrossRef]
- Müller, A.; Reiter, M.; Schötta, A.M.; Stockinger, H.; Stanek, G. Detection of Bartonella spp. in Ixodes ricinus ticks and Bartonella seroprevalence in human populations. Ticks Tick-Borne Dis. 2016, 7, 763–767. [Google Scholar]
- Lindblom, P.; Wilhelmsson, P.; Fryland, L.; Sjöwall, J.; Haglund, M.; Matussek, A.; Ernerudh, J.; Vene, S.; Nyman, D.; Andreassen, A.; et al. Tick-borne encephalitis virus in ticks detached from humans and follow-up of serological and clinical response. Ticks Tick-Borne Dis. 2014, 5, 21–28. [Google Scholar] [CrossRef]
- Kramski, M.; Meisel, H.; Klempa, B.; Krüger, D.H.; Pauli, G.; Nitsche, A. Detection and typing of human pathogenic hantaviruses by real-time reverse transcription-PCR and pyrosequencing. Clin. Chem. 2007, 53, 1899–1905. [Google Scholar] [CrossRef]
- Maas, M.; de Vries, A.; van Roon, A.; Takumi, K.; van der Giessen, J.; Rockx, B. High Prevalence of Tula Hantavirus in Common Voles in The Netherlands. Vector Borne Zoonotic Dis. Larchmt. N 2017, 17, 200–205. [Google Scholar] [CrossRef] [PubMed]
- De Vries, A.; Vennema, H.; Bekker, D.L.; Maas, M.; Adema, J.; Opsteegh, M.; van der Giessen, J.W.B.; Reusken, C.B.E.M. Characterization of Puumala hantavirus in bank voles from two regions in the Netherlands where human cases occurred. J. Gen. Virol. 2016, 97, 1500–1510. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.F.; Regnery, R.; Jameson, P.; Greene, C.; Krause, D.C. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J. Clin. Microbiol. 1995, 33, 1797–1803. [Google Scholar] [PubMed]
- Tijsse-Klasen, E.; Fonville, M.; Gassner, F.; Nijhof, A.M.; Hovius, E.K.E.; Jongejan, F.; Takken, W.; Reimerink, J.R.; Overgaauw, P.A.M.; Sprong, H. Absence of zoonotic Bartonella species in questing ticks: First detection of Bartonella clarridgeiae and Rickettsia felis in cat fleas in the Netherlands. Parasit. Vectors 2011, 4, 61. [Google Scholar] [CrossRef]
- De’ath, G.; Fabricius, K.E. Classification and regression trees: A powerful yet simple technique for ecological data analysis. Ecology 2000, 81, 3178–3192. [Google Scholar] [CrossRef]
- Krzywinski, M.; Altman, N. Points of Significance: Classification and regression trees. Nat. Methods 2017, 14, 757–758. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Therneau, T.; Atkinson, B.; Ripley, B. Rpart: Recursive Partitioning and Regression Trees; Mayo Foundation: Rochester, MN, USA, 2019; Available online: https://cran.r-project.org/web/packages/rpart/index.html (accessed on 2 October 2019).
- Mann, H.B.; Whitney, D.R. On a Test of Whether one of Two Random Variables is Stochastically Larger than the Other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Quaye, I.K. Haptoglobin, inflammation and disease. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 735–742. [Google Scholar] [CrossRef]
- Horrocks, N.P.C.; Hegemann, A.; Matson, K.D.; Hine, K.; Jaquier, S.; Shobrak, M.; Williams, J.B.; Tinbergen, J.M.; Tieleman, B.I. Immune Indexes of Larks from Desert and Temperate Regions Show Weak Associations with Life History but Stronger Links to Environmental Variation in Microbial Abundance. Physiol. Biochem. Zool. 2012, 85, 504–515. [Google Scholar] [CrossRef]
- Jain, S.; Gautam, V.; Naseem, S. Acute-phase proteins: As diagnostic tool. J. Pharm. Bioallied Sci. 2011, 3, 118–127. [Google Scholar] [CrossRef]
- Richter, D.; Matuschka, F.-R.; Spielman, A.; Mahadevan, L. How ticks get under your skin: Insertion mechanics of the feeding apparatus of Ixodes ricinus ticks. Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef] [PubMed]
- Sudhindra, P.; Wang, G.; Schriefer, M.E.; McKenna, D.; Zhuge, J.; Krause, P.J.; Marques, A.R.; Wormser, G.P. Insights into Borrelia miyamotoi infection from an untreated case demonstrating relapsing fever, monocytosis and a positive C6 Lyme serology. Diagn. Microbiol. Infect. Dis. 2016, 86, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Frank, C. Über die Bedeutung von Laelaps agilis C.L. Koch 1836 (Mesostigmata: Parasitiformae) für die Übertragung von Hepatozoon sylvatici Coles 1914 (Sporozoa: Haemogregarinidae). Z. Parasitenkd. 1977, 53, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Laakkonen, J.; Sukura, A.; Oksanen, A.; Henttonen, H.; Soveri, T. Haemogregarines of the genus Hepatozoon (Apicomplexa: Adeleina) in rodents from northern Europe. Folia Parasitol. 2001, 48, 263–267. [Google Scholar] [CrossRef]
- Smith, T.G. The Genus Hepatozoon (Apicomplexa: Adeleina). J. Parasitol. 1996, 82, 565–585. [Google Scholar] [CrossRef]
- Shutler, D.; Smith, T.G.; Robinson, S.R. Relationships between leukocytes and Hepatozoon spp. In green frogs, Rana clamitans. J. Wildl. Dis. 2009, 45, 67–72. [Google Scholar] [CrossRef]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Reynolds, L.A.; Filbey, K.J.; Maizels, R.M. Immunity to the model intestinal helminth parasite Heligmosomoides polygyrus. Semin. Immunopathol. 2012, 34, 829–846. [Google Scholar] [CrossRef]
- Martin, L.B.; Burgan, S.C.; Adelman, J.S.; Gervasi, S.S. Host Competence: An Organismal Trait to Integrate Immunology and Epidemiology. Integr. Comp. Biol. 2016, 56, 1225–1237. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Parasite | Bank Vole (n = 10) | Wood Mouse (n = 26) | ||
|---|---|---|---|---|
| Microparasites | ||||
| Bartonella grahamii 1 | 0.40 | 0.04 | ||
| Bartonella taylorii 1 | 0.10 | 0.31 | ||
| Borrelia burgdorferi s.l. 1 | 0 | 0.04 | ||
| Borrelia miyamotoi 1 | 0.10 | 0.23 | ||
| Candidatus Neoehrlichia mikurensis 1 | 0.30 | 0.12 | ||
| Ectoparasites | ||||
| Ixodes ricinus (larvae) 2 | 3.20 (0–8) | [0.80] | 11.08 (0–80) | [0.92] |
| Ixodes ricinus (nymphs) 2 | 0 | [0] | 0.08 (0–2) | [0.04] |
| Ixodes trianguliceps (larvae) 2 | 0.70 (0–4) | [0.40] | 0.38 (0–4) | [0.15] |
| Echinonyssus isabellinus 2 | 0.10 (0–1) | [0.10] | 0 | [0] |
| Eulaelaps stabularis 2 | 0 | [0] | 1.23 (0–27) | [0.19] |
| Haemogamasus nidi 2 | 0 | [0] | 0.35 (0–9) | [0.04] |
| Laelaps agilis2 | 0 | [0] | 3.69 (0–27) | [0.58] |
| Ctenophtalmus agyrtes 2 | 0.10 (0–1) | [0.10] | 0.08 (0–1) | [0.08] |
| Megabothris turbidus 2 | 0 | [0] | 0.08 (0–2) | [0.04] |
| Polyplax serrata 2 | 0 | [0] | 0.85 (0–8) | [0.19] |
| Gastrointestinal parasites | ||||
| Aonchoteca murissylvatici 2 | 2.10 (0–14) | [0.30] | 5.65 (0–75) | [0.35] |
| Aspiculuris tianjinensis 2 | 15.40 (0–150) | [0.20] | 2.31 (0–29) | [0.19] |
| Heligmosomoides polygyrus 2 | 2.80 (0–18) | [0.40] | 0.38 (0–3) | [0.31] |
| Heterakis spumosa 2 | 0 | [0] | 0.04 (0–1) | [0.04] |
| Hymenolepis diminuta 2 | 0.10 (0–1) | [0.10] | 0 | [0] |
| Syphacia petruzewiczi 2 | 3.80 (0–24) | [0.20] | 0 | [0] |
| Syphacia stroma 2 | 0 | [0] | 12.46 (0–76) | [0.58] |
| Non-Trichinella coiled nematode 2 | 0.90 (0–7) | [0.20] | 0 | [0] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofmeester, T.R.; Bügel, E.J.; Hendrikx, B.; Maas, M.; Franssen, F.F.J.; Sprong, H.; Matson, K.D. Parasite Load and Site-Specific Parasite Pressure as Determinants of Immune Indices in Two Sympatric Rodent Species. Animals 2019, 9, 1015. https://doi.org/10.3390/ani9121015
Hofmeester TR, Bügel EJ, Hendrikx B, Maas M, Franssen FFJ, Sprong H, Matson KD. Parasite Load and Site-Specific Parasite Pressure as Determinants of Immune Indices in Two Sympatric Rodent Species. Animals. 2019; 9(12):1015. https://doi.org/10.3390/ani9121015
Chicago/Turabian StyleHofmeester, Tim R., Esther J. Bügel, Bob Hendrikx, Miriam Maas, Frits F. J. Franssen, Hein Sprong, and Kevin D. Matson. 2019. "Parasite Load and Site-Specific Parasite Pressure as Determinants of Immune Indices in Two Sympatric Rodent Species" Animals 9, no. 12: 1015. https://doi.org/10.3390/ani9121015
APA StyleHofmeester, T. R., Bügel, E. J., Hendrikx, B., Maas, M., Franssen, F. F. J., Sprong, H., & Matson, K. D. (2019). Parasite Load and Site-Specific Parasite Pressure as Determinants of Immune Indices in Two Sympatric Rodent Species. Animals, 9(12), 1015. https://doi.org/10.3390/ani9121015