Rumen Fermentation Characteristics in Pre- and Post-Weaning Calves upon Feeding with Mulberry Leaf Flavonoids and Candida tropicalis Individually or in Combination as a Supplement



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Flavonoids and Active Dry Yeast
2.2. Animals, Diets, and Experimental Design
2.3. Measurements, Sample Collection, and Analysis
2.4. Statistical Analysis
3. Results
3.1. Intake, Growth Performance, and Fecal Score
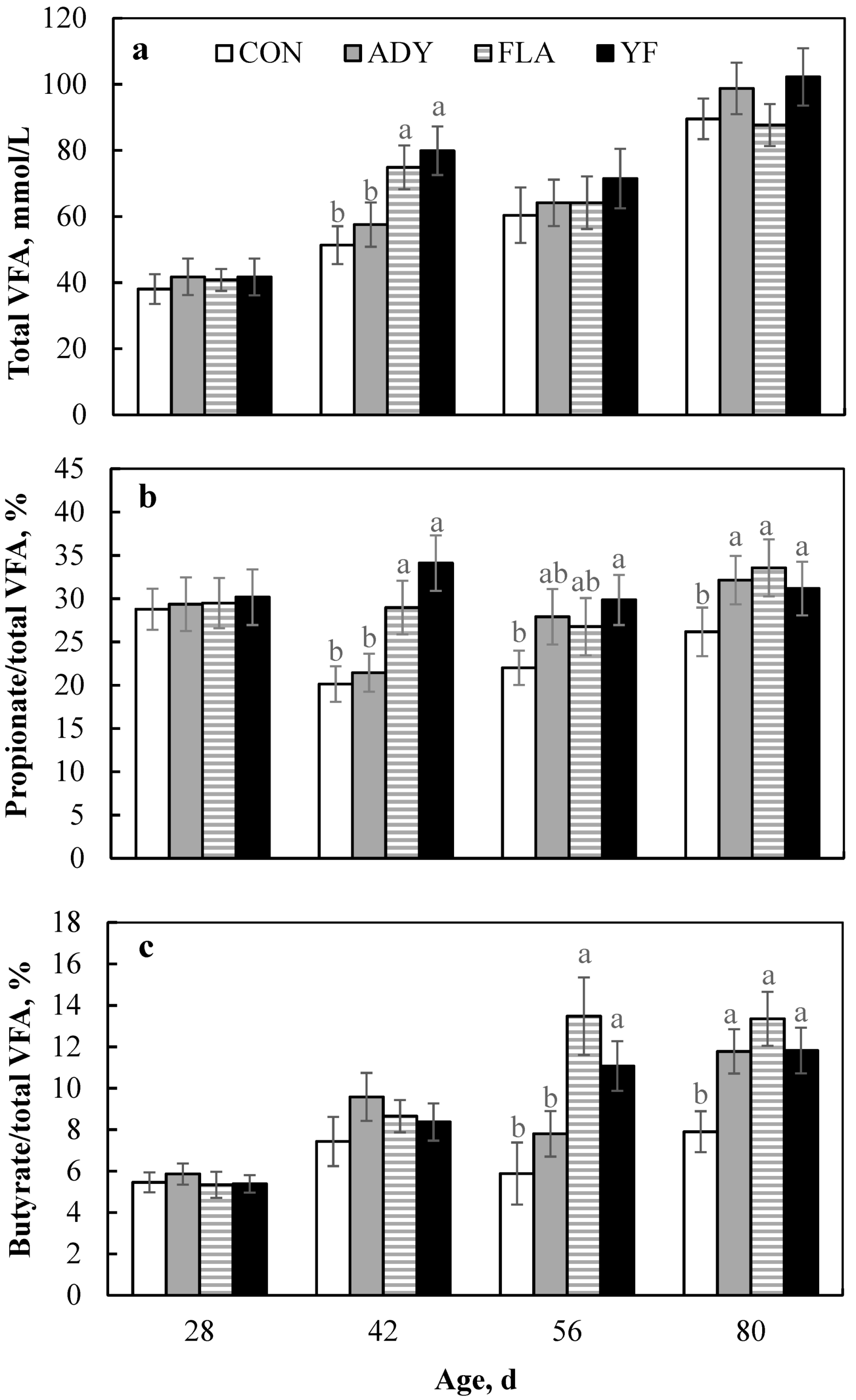
3.2. Rumen Fermentation Characteristics
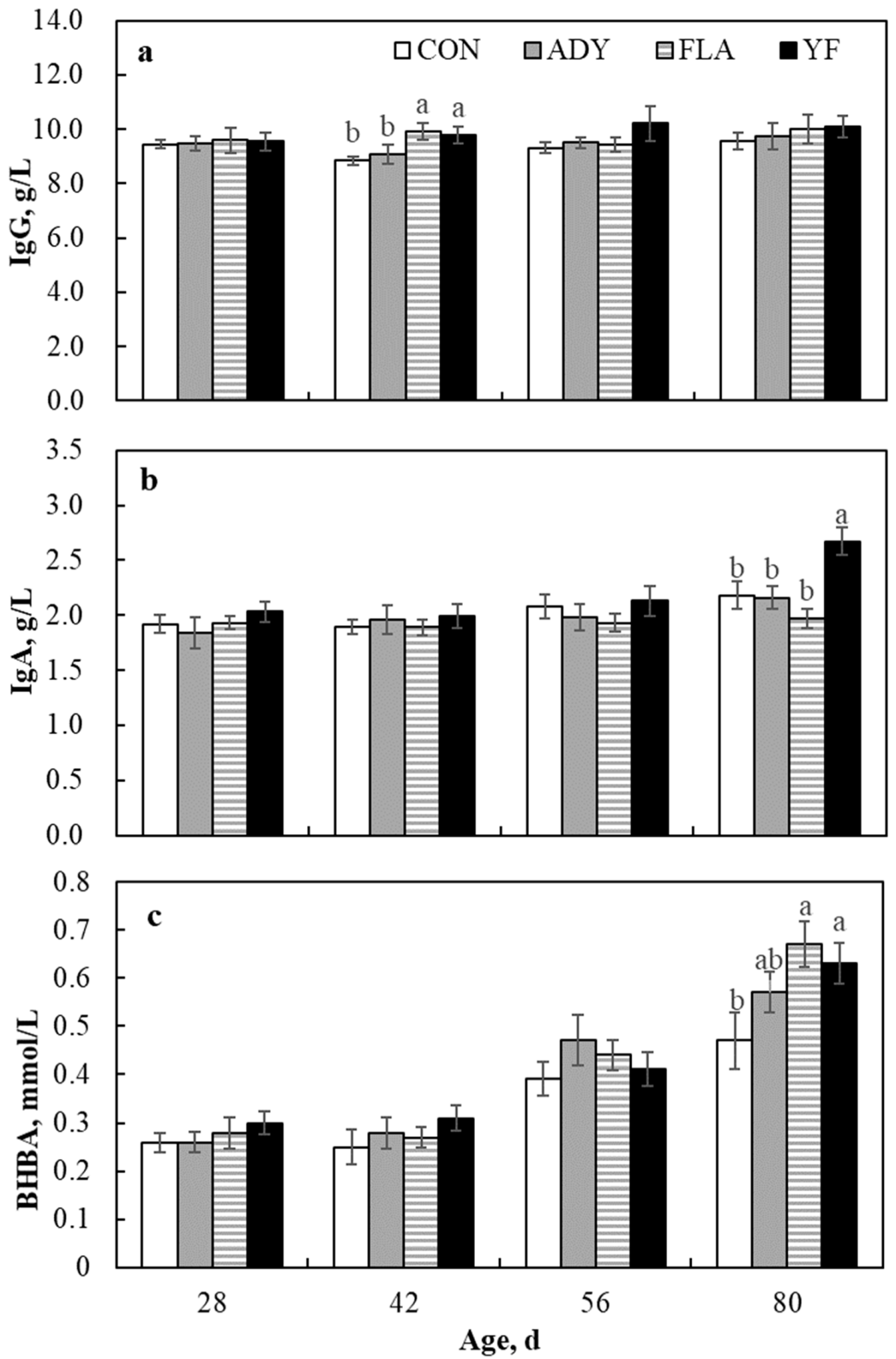
3.3. Blood Biomarkers
4. Discussion
4.1. Growth and Health Performance
4.2. Rumen Fermentation Parameters
4.3. Blood Parameters
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Magalhães, V.J.A.; Susca, F.; Lima, F.S.; Branco, A.F.; Yoon, I.; Santos, J.E.P. Effect of feeding yeast culture on performance, health, and immunocompetence of dairy calves. J. Dairy Sci. 2008, 91, 1497–1509. [Google Scholar] [CrossRef]
- Gudlaugsson, O.; Gillespie, S.; Lee, K.; Berg, J.V.; Hu, J.; Messer, S.; Herwaldt, L.; Pfaller, M.; Diekema, D. Attributable mortality of nosocomial candidiasis, revisited. Clin. Infect. Dis. 2003, 37, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Megri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. Fems. Microbiol. Rev. 2011, 36, 288–305. [Google Scholar] [CrossRef] [PubMed]
- Moran, G.; Coleman, D.; Sullivan, D. An Introduction to the Medically Important Candida Species, 2nd ed.; ASM Press: Washington, DC, USA, 2012; pp. 11–25. [Google Scholar]
- Hallen-Adams, H.E.; Kachman, S.D.; Kim, J.; Legge, R.M.; Martinez, I. Fungi inhabiting the healthy human gastrointestinal tract: A diverse and dynamic community. Fungal. Ecol. 2015, 15, 9–17. [Google Scholar] [CrossRef]
- Zhang, W.J.; Xu, Z.R.; Zhao, S.H.; Sun, J.Y.; Yang, X. Development of a microbial fermentation process for detoxification of gossypol in cottonseed meal. Anim. Feed Sci. Technol. 2007, 135, 176–186. [Google Scholar] [CrossRef]
- Marrero, Y.; Castillo, Y.; Burrola-Barraza, M.E.; Lobaina, T.; Rosa, C.A.; Ruiz, O.; Gonzalez-Rodriguez, E.; Basso, L.C. Morphological, biochemical and molecular identification of the yeast Levica 25: A Potential Ruminal Microbial Additive. Glob. Vet. 2011, 7, 60–85. [Google Scholar]
- Marrero, Y.; Castillo, Y.; Ruiz, O.; Burrola, E.; Angulo, C. Feeding of yeast (Candida spp.) improves in vitro ruminal fermentation of fibrous substrates. J. Integr. Agric. 2015, 14, 514–519. [Google Scholar] [CrossRef]
- Timmerman, H.M.; Koning, C.J.M.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef]
- Bayatkouhsar, J.; Tahmasebi, A.M.; Naserian, A.A.; Mokarram, R.R.; Valizadeh, R. Effects of supplementation of lactic acid bacteria on growth performance, blood metabolites and fecal coliform and lactobacilli of young dairy calves. Anim. Feed Sci. Technol. 2013, 186, 1–11. [Google Scholar] [CrossRef]
- Rochfort, S.; Parker, A.J.; Dunshea, F.R. Plant bioactives for ruminant health and productivity. Phytochemistry 2008, 69, 299–322. [Google Scholar] [CrossRef]
- Stoldt, A.K.; Derno, M.; Nurnberg, G.; Weitzel, J.M.; Otten, W.; Starke, A.; Wolffram, S.; Metges, C.C. Effects of a 6-wk intraduodenal supplementation of quercetin reduces indicators of liver damage in periparturient dairy cows. J. Dairy Sci. 2015, 98, 4509–4520. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.D.; Chen, X.L.; Tu, Y.; Wang, B.; Lou, C.; Ma, T.; Diao, Q.Y. Effects of mulberry leaf flavonoid and resveratrol on methane emission and nutrient digestion in sheep. Anim. Nutr. 2015, 1, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.M.; Wein, S.; Blank, R.; Metges, C.C.; Wolffram, S. Bioavailability of the flavonol quercetin in cows after intraruminal application of quercetin aglycone and rutin. J. Dairy Sci. 2012, 95, 5047–5055. [Google Scholar] [CrossRef] [PubMed]
- Stoldt, A.K.; Derno, M.; Das, G.; Weitzel, J.M.; Wolffram, S.; Metges, C.C. Effects of rutin and buckwheat seeds on energy metabolism and methane production in dairy cows. J. Dairy Sci. 2016, 99, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Besle, J.M.; Viala, D.; Martin, B.; Pradel, P.; Meunier, B.; Berdague, J.L.; Fraisse, D.; Lamaison, J.L.; Coulon, J.B. Ultraviolet-absorbing compounds in milk are related to forage polyphenols. J. Dairy Sci. 2010, 93, 2846–2856. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Miyazawa, M.; Kamel, A.; Abe, K.; Kojima, T. Ameliorative effects of mulberry (Morus alba L.) leaves on hyperlipidemia in rats fed a high-fat diet: Induction of fatty acid oxidation, inhibition of lipogenesis, and suppression of oxidative stress. Biosci. Biotechnol. Biochem. 2010, 74, 2385–2395. [Google Scholar] [CrossRef]
- Berger, L.M.; Blank, R.; Zorn, F.; Wein, S.; Metges, C.C.; Wolffram, S. Ruminal degradation of quercetin and its influence on fermentation in ruminants. J. Dairy Sci. 2015, 98, 5688–5698. [Google Scholar] [CrossRef]
- Chung, Y.H.; Walker, N.D.; McGinn, S.M.; Beauchemin, K.A. Differing effects of 2 active dried yeast (Saccharomyces cerevisiae) strains on ruminal acidosis and methane production in nonlactating dairy cows. J. Dairy Sci. 2011, 94, 2431–2439. [Google Scholar] [CrossRef]
- Jiang, Y.; Ogunade, I.M.; Arriola, K.G.; Qi, M.; Vyas, D.; Staples, C.R.; Adesogan, A.T. Effects of the dose and viability of Saccharomyces cerevisiae. 2. Ruminal fermentation, performance of lactating dairy cows, and correlations between ruminal bacteria abundance and performance measures. J. Dairy Sci. 2017, 100, 8102–8118. [Google Scholar] [CrossRef]
- Heinrichs, A.J.; Jones, C.M.; Heinrichs, B.S. Effects of mannan oligosaccharide or antibiotics in neonatal diets on health and growth of dairy calves. J. Dairy Sci. 2003, 86, 4064–4069. [Google Scholar] [CrossRef]
- Dong, L.F.; Zhang, W.B.; Zhang, N.F.; Tu, Y.; Diao, Q.Y. Feeding different dietary protein to energy ratios to Holstein heifers: Effects on growth performance, blood metabolites and rumen fermentation parameters. J. Anim. Physiol. Anim. Nutr. 2016, 101, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.C.; Yang, H.J. Ruminal digestibility and fermentation characteristics in vitro of fenugreek and alfalfa hay combination with or without the inoculation of Neocallimastix sp. YAK11. Anim. Feed Sci. Technol. 2011, 169, 53–60. [Google Scholar] [CrossRef]
- Balcells, J.; Aris, A.; Serrano, A.; Seradj, A.R.; Crespo, J.; Devant, M. Effects of an extract of plant flavonoids (Bioflavex) on rumen fermentation and performance in heifers fed high-concentrate diets. J. Anim. Sci. 2012, 90, 4975–4984. [Google Scholar] [CrossRef] [PubMed]
- Durmic, Z.; Moate, P.J.; Eckard, R.; Revell, D.K.; Williams, R.; Vercoe, P.E. In vitro screening of selected feed additives, plant essential oils and plant extracts for rumen methane mitigation. J. Sci. Food Agric. 2014, 94, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Elghandour, M.M.Y.; Chagoyán, J.C.V.; Salem, A.Z.M.; Kholif, A.E.; Castaneda, J.S.M.; Camacho, L.M.; Buendia, G. In vitro fermentative capacity of equine fecal inocula of nine fibrous forages in presence of different doses of Saccharomyces cerevisiae. J. Equine Vet. Sci. 2014, 34, 619–625. [Google Scholar] [CrossRef]
- Kumprechtová, D.; Illek, J.; Julien, C.; Homolka, P.; Jancik, F.; Auclair, E. Effect of live yeast (Saccharomyces cerevisiae.) supplementation on rumen fermentation and metabolic profile of dairy cows in early lactation. J. Anim. Physiol. Anim. Nutr. 2019, 103, 447–455. [Google Scholar]
- Geiger, A.J.; Ward, S.H.; Williams, C.C.; Rude, B.J.; Cabrera, C.J.; Kalestch, K.N.; Voelz, B.E. Short communication: Effects of increasing protein and energy in the milk replacer with or without direct-fed microbial supplementation on growth and performance of preweaned Holstein calves. J. Dairy Sci. 2014, 97, 7212–7219. [Google Scholar] [CrossRef]
- Yaghoubi, S.M.J.; Ghorbani, G.R.; Rahmani, H.R.; Nikkhah, A. Growth, weaning performance and blood indicators of humoral immunity in Holstein calves fed supplemental flavonoids. J. Anim. Physiol. Anim. Nutr. 2008, 92, 456–462. [Google Scholar] [CrossRef]
- Bainter, K. Vacuolation in the young. In Biology of the Intestine in Growing Animals; Zabielski, R., Gregory, P.C., Westrom, B., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 55–110. [Google Scholar]
- Blum, J.W. Nutritional physiology of neonatal calves. J. Anim. Physiol. Anim. Nutr. 2006, 90, 1–11. [Google Scholar] [CrossRef]
- Maciej, J.; Schäff, C.T.; Kanitz, E.; Tuchscherer, A.; Bruckmaier, R.M.; Wolffram, S.; Hammon, H.M. Bioavailability of the flavonol quercetin in neonatal calves after oral administration of quercetin aglycone or rutin. J. Dairy Sci. 2015, 98, 3906–3917. [Google Scholar] [CrossRef]
- Oskoueian, E.; Abdullah, N.; Oskoueian, A. Effects of flavonoids on rumen fermentation activity, methane production, and microbial population. BioMed Res. Int. 2013. under review. [Google Scholar] [CrossRef] [PubMed]
- Seradj, A.R.; Abecia, L.; Crespo, J.; Villalba, D.; Fondevila, M.; Balcells, J. The effect of Bioflavex and its pure flavonoid components on in vitro fermentation parameters and methane production in rumen fluid from steers given high concentrate diets. Anim. Feed Sci. Technol. 2014, 197, 85–91. [Google Scholar] [CrossRef]
- Bayat, A.R.; Kairenius, P.; Stefański, T.; Leskinen, H.; Comtet-Marre, S.; Forano, E.; Chaucheyras-Durand, F.; Shingfield, K.J. Effect of camelina oil or live yeasts (Saccharomyces cerevisiae) on ruminal methane production, rumen fermentation, and milk fatty acid composition in lactating cows fed grass silage diets. J. Dairy Sci. 2014, 98, 3166–3181. [Google Scholar] [CrossRef] [PubMed]
- Aikman, P.C.; Henning, P.H.; Humphries, D.J.; Horn, C.H. Rumen pH and fermentation characteristics in dairy cows supplemented with Megasphaera elsdenii NCIMB 41125 in early lactation. J. Dairy Sci. 2010, 94, 2840–2849. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.H.; Zoetendal, E.; Mackie, R.I. Bacterial mechanisms to overcome inhibitory effects of dietary tannins. Microb. Ecol. 2005, 50, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Gorka, P.; Kowalski, Z.M.; Pietrzak, P.; Kotunia, A.; Kiljanczyk, R.; Flaga, J.; Holst, J.J.; Guilloteau, P.; Zabielski, R. Effect of sodium butyrate supplementation in milk replacer and starter diet on rumen development in calves. J. Physiol. Pharm. 2009, 60, 47–53. [Google Scholar]
- Hristov, A.N.; Varga, G.; Cassidy, T.; Long, M.; Heyler, K.; Karnati, S.K.R.; Corl, B.; Hovde, C.J.; Yoon, I. Effect of Saccharomyces cerevisiae fermentation product on ruminal fermentation and nutrient utilization in dairy cows. J. Dairy Sci. 2009, 93, 682–692. [Google Scholar] [CrossRef]
- Sun, P.; Wang, J.Q.; Zhang, H.T. Effects of Bacillus Subtilis Natto on performance and immune function of preweaning calves. J. Dairy Sci. 2010, 93, 5851–5855. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Ravetch, J.V. Antibody-mediated modulation of immune responses. Immunol. Rev. 2010, 236, 265–275. [Google Scholar] [CrossRef]
- Kaburagi, T.; Yamano, T.; Fukushima, Y.; Yoshino, H.; Mito, N.; Sato, K. Effect of Lactobacillus johnsonii La1 on immune function and serum albumin in aged and malnourished aged mice. Nutrition 2007, 23, 342–350. [Google Scholar] [CrossRef]
- Quigley, J.R.; Caldwell, L.A.; Sinks, G.D.; Heitmann, R.N. Changes in blood glucose, nonesterified fatty acids, and ketones in response to weaning and feed intake in young calves. J. Dairy Sci. 1991, 74, 250–257. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Item | Starter | Milk Replacer |
|---|---|---|
| Dry matter, % as fed | 85.36 | 95.36 |
| Organic matter | 92.21 | 94.85 |
| Crude protein | 19.08 | 24.27 |
| Gross energy, MJ/kg of DM | 15.45 | 19.86 |
| Neutral detergent fiber | 18.59 | 4.02 |
| Acid detergent fiber | 10.65 | 2.11 |
| Ether extract | 2.21 | 12.85 |
| Calcium | 1.09 | 1.07 |
| Phosphorus | 0.47 | 0.48 |
| Item 1 | Treatment 2 | SEM 3 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | ADY | FLA | YF | Treatment | Time | Treatment × Time | ||
| DM intake, kg/d | ||||||||
| Milk replacer | ||||||||
| Pre-weaning | 0.76 | 0.76 | 0.78 | 0.77 | 0.02 | 0.618 | 0.445 | 0.592 |
| Starter | ||||||||
| Pre-weaning | 0.25 | 0.27 | 0.27 | 0.30 | 0.02 | 0.221 | 0.194 | 0.217 |
| Post-weaning | 1.40 | 1.49 | 1.51 | 1.65 | 0.03 | 0.130 | 0.691 | 0.541 |
| Overall | 0.96 | 0.99 | 1.02 | 1.11 | 0.02 | 0.205 | 0.427 | 0.313 |
| Total DMI, kg/d | ||||||||
| Pre-weaning | 1.00 | 1.03 | 1.05 | 1.08 | 0.02 | 0.124 | 0.242 | 0.121 |
| Overall | 1.12 | 1.13 | 1.16 | 1.22 | 0.02 | 0.212 | 0.117 | 0.422 |
| Average daily gain, kg/d | ||||||||
| Pre-weaning | 0.48 | 0.51 | 0.48 | 0.50 | 0.03 | 0.134 | 0.281 | 0.226 |
| Post-weaning | 0.69 b | 0.70 b | 0.77 a | 0.79 a | 0.02 | 0.032 | 0.028 | 0.032 |
| Overall | 0.58 b | 0.60 ab | 0.63 a | 0.64 a | 0.03 | 0.011 | 0.028 | 0.013 |
| Feed efficiency, kg/kg | ||||||||
| Pre-weaning | 0.51 | 0.53 | 0.56 | 0.53 | 0.01 | 0.286 | 0.226 | 0.100 |
| Post-weaning | 0.47 b | 0.55 a | 0.55 a | 0.50 b | 0.02 | 0.031 | 0.034 | 0.025 |
| Overall | 0.50 b | 0.54 a | 0.55 a | 0.51 b | 0.01 | 0.012 | 0.032 | 0.017 |
| Fecal score | ||||||||
| Pre-weaning | 1.53 a | 1.45 ab | 1.42 b | 1.42 b | 0.03 | 0.039 | 0.008 | 0.125 |
| Post-weaning | 1.26 | 1.19 | 1.14 | 1.16 | 0.02 | 0.325 | 0.032 | 0.742 |
| Overall | 1.39 | 1.30 | 1.29 | 1.30 | 0.02 | 0.073 | 0.025 | 0.513 |
| Item 1 | Treatment 2 | SEM 3 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | ADY | FLA | YF | Treatment | Time | Treatment × Time | ||
| pH | ||||||||
| Pre-weaning | 6.2 | 6.5 | 6.3 | 6.3 | 0.8 | 0.213 | 0.565 | 0.182 |
| Post-weaning | 6.2 | 6.4 | 6.4 | 6.3 | 0.7 | 0.397 | 0.020 | 0.191 |
| Overall | 6.1 | 6.4 | 6.3 | 6.3 | 0.7 | 0.265 | 0.154 | 0.290 |
| NH3-N, mmol/L | ||||||||
| Pre-weaning | 4.44 | 4.22 | 5.18 | 4.09 | 1.71 | 0.260 | 0.380 | 0.513 |
| Post-weaning | 7.52 | 7.36 | 7.40 | 7.33 | 1.05 | 0.340 | 0.280 | 0.316 |
| Overall | 5.49 | 5.42 | 5.47 | 5.18 | 1.92 | 0.213 | 0.342 | 0.253 |
| Total VFA, mmol/L | ||||||||
| Pre-weaning | 44.20 b | 49.99 b | 57.81 a | 60.90 a | 3.21 | 0.027 | 0.018 | 0.014 |
| Post-weaning | 75.96 | 81.45 | 71.27 | 83.85 | 5.27 | 0.253 | 0.561 | 0.316 |
| Overall | 59.19 b | 64.40 b | 66.83 ab | 74.12 a | 4.35 | 0.031 | 0.002 | 0.025 |
| Acetate/total VFA, % | ||||||||
| Pre-weaning | 57.60 | 61.40 | 61.17 | 56.67 | 3.35 | 0.917 | 0.041 | 0.635 |
| Post-weaning | 55.13 | 56.75 | 52.50 | 58.75 | 2.12 | 0.208 | 0.466 | 0.446 |
| Overall | 54.14 | 54.36 | 56.21 | 55.79 | 2.04 | 0.849 | 0.617 | 0.559 |
| Propionate/total VFA, % | ||||||||
| Pre-weaning | 24.20 b | 25.40 b | 29.50 ab | 32.83 a | 2.92 | 0.047 | 0.036 | 0.038 |
| Post-weaning | 24.09 b | 29.50 a | 30.13 a | 30.50 a | 1.64 | 0.045 | 0.747 | 0.838 |
| Overall | 24.13 b | 27.92 ab | 29.86 ab | 31.50 a | 1.84 | 0.018 | 0.026 | 0.017 |
| Butyrate/total VFA, % | ||||||||
| Pre-weaning | 6.40 | 8.60 | 6.83 | 7.00 | 1.16 | 0.455 | 0.108 | 0.740 |
| Post-weaning | 6.63 | 9.75 | 13.25 | 11.38 | 1.87 | 0.055 | 0.337 | 0.839 |
| Overall | 6.54 | 9.31 | 10.50 | 9.50 | 1.07 | 0.193 | 0.004 | 0.669 |
| Acetate/propionate ratio | ||||||||
| Pre-weaning | 2.37 a | 2.41 a | 2.05 ab | 1.77 b | 0.30 | 0.047 | 0.055 | 0.044 |
| Post-weaning | 2.29 a | 1.92 b | 1.74 b | 1.93 b | 0.19 | 0.031 | 0.618 | 0.779 |
| Overall | 2.24 a | 1.95 ab | 1.88 b | 1.77 b | 0.26 | 0.042 | 0.051 | 0.033 |
| Item 1 | Treatment 2 | SEM 3 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | ADY | FLA | YF | Treatment | Time | Treatment × Time | ||
| IgG, g/L | ||||||||
| Pre-weaning | 8.93 b | 9.12 b | 9.64 a | 9.68 a | 0.19 | 0.032 | 0.341 | 0.008 |
| Post-weaning | 9.45 | 9.62 | 9.74 | 10.15 | 0.27 | 0.120 | 0.258 | 0.364 |
| Overall | 9.09 b | 9.27 ab | 9.72 a | 9.78 a | 0.19 | 0.041 | 0.003 | 0.052 |
| IgM, g/L | ||||||||
| Pre-weaning | 0.84 | 0.83 | 0.83 | 0.86 | 0.01 | 0.438 | 0.204 | 0.511 |
| Post-weaning | 0.85 | 0.91 | 0.90 | 0.90 | 0.16 | 0.085 | 0.338 | 0.213 |
| Overall | 0.84 | 0.86 | 0.85 | 0.87 | 0.01 | 0.255 | 0.111 | 0.302 |
| IgA, g/L | ||||||||
| Pre-weaning | 1.96 | 1.93 | 1.92 | 2.05 | 0.02 | 0.551 | 0.399 | 0.612 |
| Post-weaning | 2.13 b | 2.07 b | 1.95 b | 2.40 a | 0.02 | 0.013 | 0.632 | 0.021 |
| Overall | 2.02 b | 1.98 b | 1.93 b | 2.20 a | 0.04 | 0.031 | 0.031 | 0.333 |
| BHBA, mmol/L | ||||||||
| Pre-weaning | 0.30 | 0.35 | 0.33 | 0.34 | 0.02 | 0.487 | 0.677 | 0.478 |
| Post-weaning | 0.43 b | 0.55 a | 0.56 a | 0.52 ab | 0.04 | 0.024 | 0.012 | 0.015 |
| Overall | 0.38 | 0.41 | 0.42 | 0.41 | 0.31 | 0.181 | <0.001 | 0.406 |
| EGF, pg/mL | ||||||||
| Pre-weaning | 28.83 ab | 26.47 b | 35.38 a | 28.04 ab | 1.49 | 0.047 | 0.193 | 0.180 |
| Post-weaning | 30.23 | 28.29 | 33.44 | 29.57 | 2.07 | 0.217 | 0.418 | 0.321 |
| Overall | 29.44 | 27.23 | 34.89 | 28.94 | 1.80 | 0.161 | 0.586 | 0.606 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, L.; Yang, C.; Dong, L.; Diao, Q.; Si, B.; Ma, J.; Tu, Y. Rumen Fermentation Characteristics in Pre- and Post-Weaning Calves upon Feeding with Mulberry Leaf Flavonoids and Candida tropicalis Individually or in Combination as a Supplement. Animals 2019, 9, 990. https://doi.org/10.3390/ani9110990
Kong L, Yang C, Dong L, Diao Q, Si B, Ma J, Tu Y. Rumen Fermentation Characteristics in Pre- and Post-Weaning Calves upon Feeding with Mulberry Leaf Flavonoids and Candida tropicalis Individually or in Combination as a Supplement. Animals. 2019; 9(11):990. https://doi.org/10.3390/ani9110990
Chicago/Turabian StyleKong, Luxin, Chuntao Yang, Lifeng Dong, Qiyu Diao, Bingwen Si, Junnan Ma, and Yan Tu. 2019. "Rumen Fermentation Characteristics in Pre- and Post-Weaning Calves upon Feeding with Mulberry Leaf Flavonoids and Candida tropicalis Individually or in Combination as a Supplement" Animals 9, no. 11: 990. https://doi.org/10.3390/ani9110990
APA StyleKong, L., Yang, C., Dong, L., Diao, Q., Si, B., Ma, J., & Tu, Y. (2019). Rumen Fermentation Characteristics in Pre- and Post-Weaning Calves upon Feeding with Mulberry Leaf Flavonoids and Candida tropicalis Individually or in Combination as a Supplement. Animals, 9(11), 990. https://doi.org/10.3390/ani9110990