Oxidative Stress and Inflammation in Sows with Excess Backfat: Up-Regulated Cytokine Expression and Elevated Oxidative Stress Biomarkers in Placenta


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Care and Use of Animals
2.2. Tissue Collection
2.3. Lipid Analyses
2.4. Free Fatty Acid (FAA) Assay
2.5. Reactive Oxygen Species (ROS) and Lipid Oxidation Assay
2.6. Placenta Histology and Vascular Density Determination
2.7. Real-Time PCR
2.8. Statistical Analysis
3. Results
3.1. Placental Lipids in Different Backfat Thickness Sows
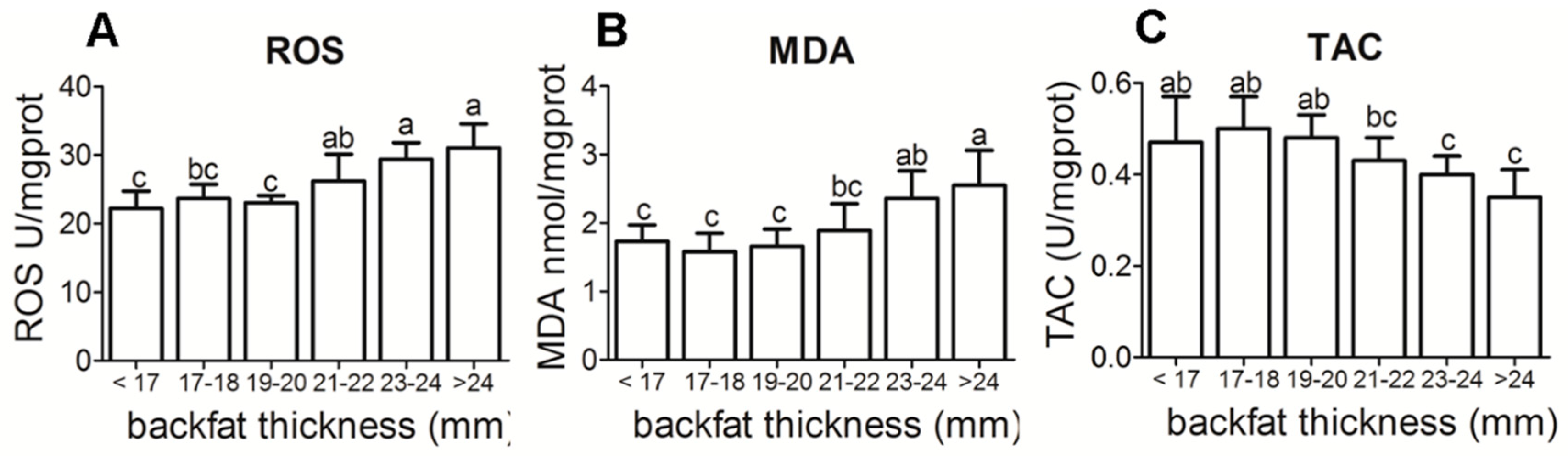
3.2. Placental Reactive Oxygen Species (ROS), Malondialdehyde (MDA) and Total Antioxidant Capacity (TAC) Levels in Different Backfat Thickness Sows
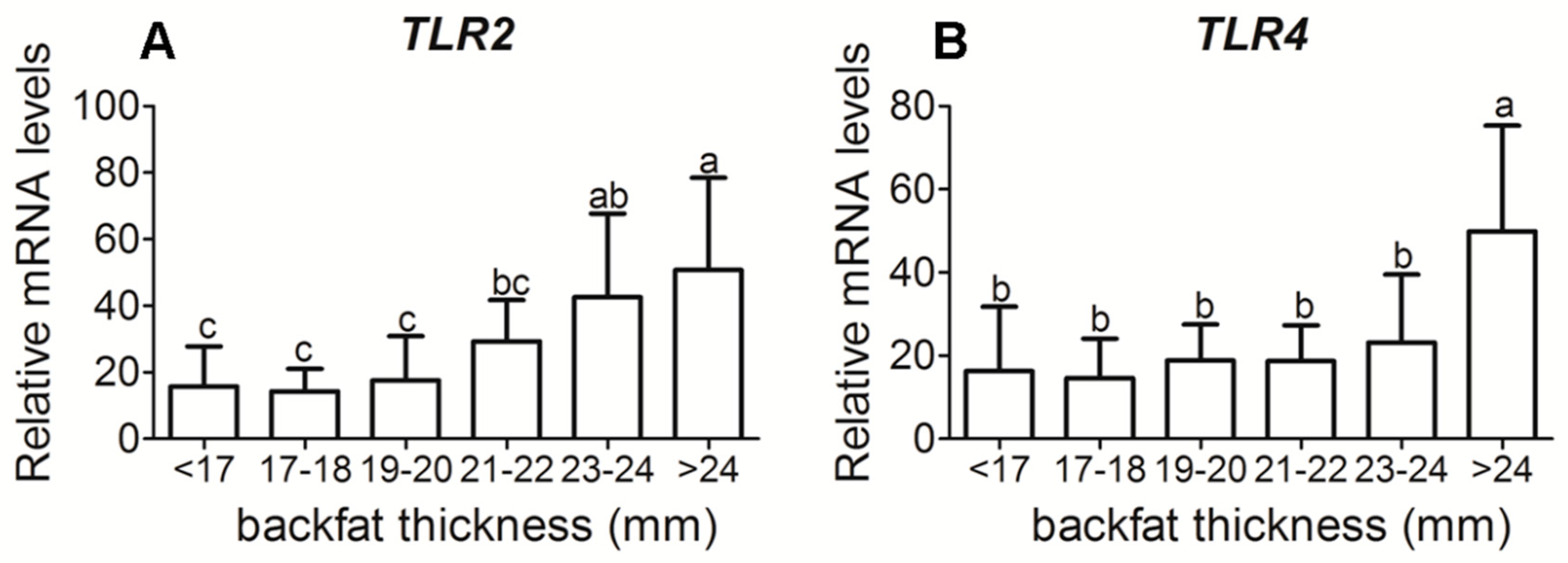
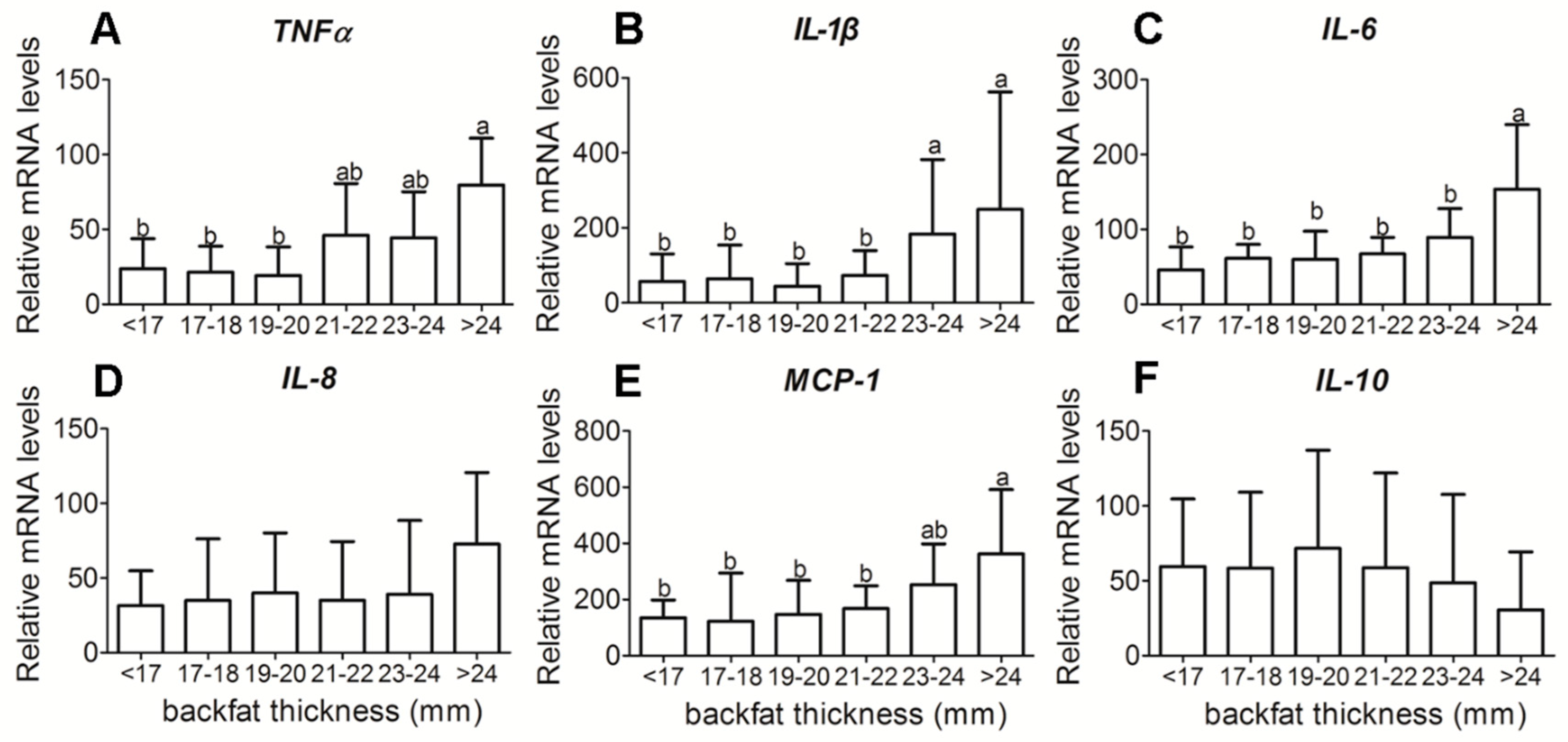
3.3. Placental Inflammatory-Related Cytokine Gene Expression in Different Backfat Thickness Sows
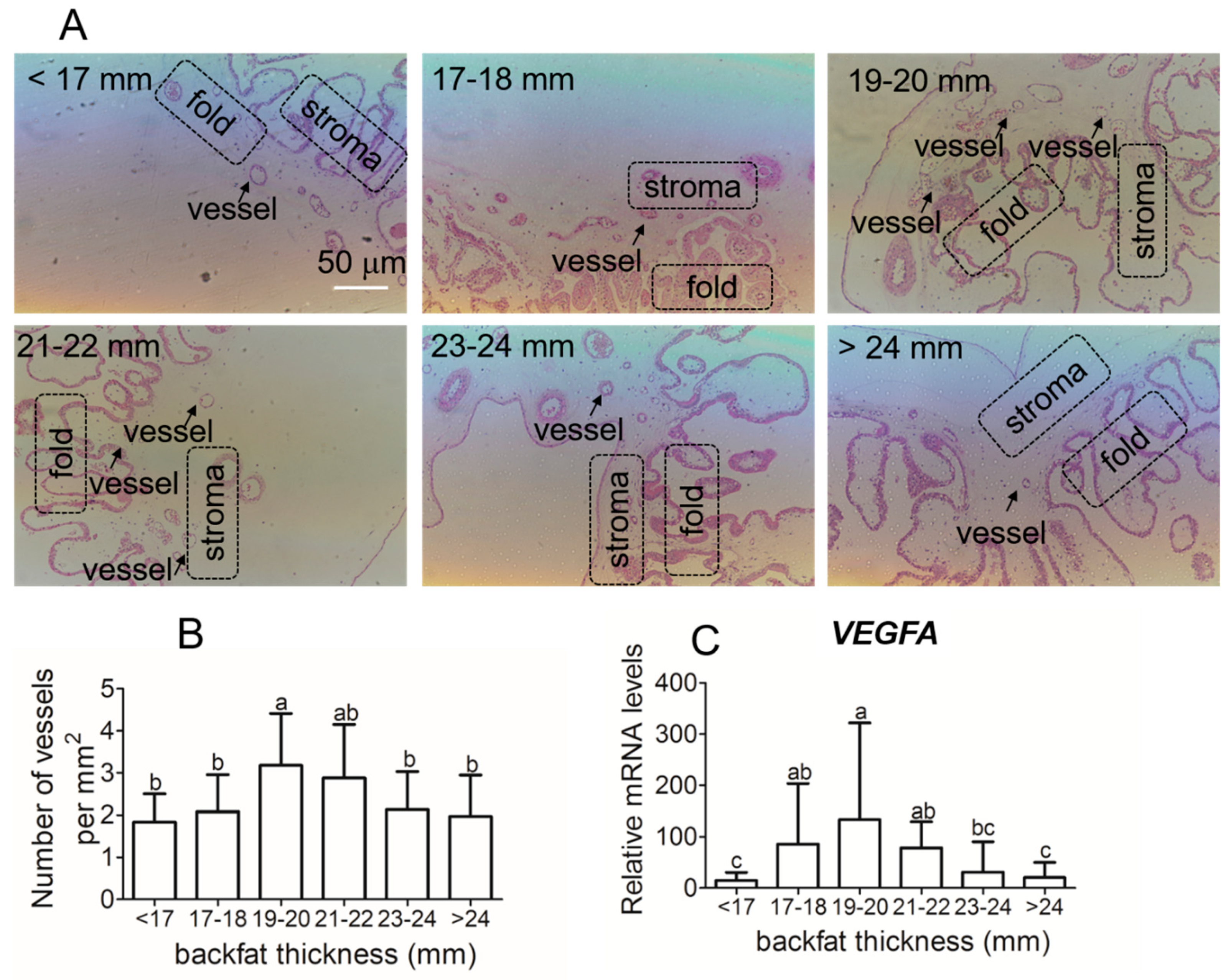
3.4. Placental Vessel Density and Angiogenic Regulators Gene Expression in Different Backfat Thickness Sows
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Bonis, M.; Torricelli, M.; Severi, F.M.; Luisi, S.; De Leo, V.; Petraglia, F. Neuroendocrine aspects of placenta and pregnancy. Gynecol. Endocrinol. 2012, 1, 22–26. [Google Scholar] [CrossRef]
- Howell, K.R.; Powell, T.L. Effects of maternal obesity on placental function and fetal development. Reproduction 2017, 153, 97–108. [Google Scholar] [CrossRef]
- Ma, Y.; Zhu, M.J.; Zhang, L.; Hein, S.M.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity and overnutrition alter fetal growth rate and cotyledonary vascularity and angiogenic factor expression in the ewe. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, 249–258. [Google Scholar] [CrossRef]
- Saben, J.; Lindsey, F.; Zhong, Y.; Thakali, K.; Badger, T.M.; Andres, A.; Gomez-Acevedo, H.; Shankar, K. Maternal obesity is associated with a lipotoxic placental environment. Placenta 2014, 35, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.A.; Riley, S.C.; Reynolds, R.M.; Barr, S.; Evans, M.; Statham, A.; Hor, K.; Jabbour, H.N.; Norman, J.E.; Denison, F.C. Placental structure and inflammation in pregnancies associated with obesity. Placenta 2011, 32, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Oliva, K.; Barker, G.; Riley, C.; Bailey, M.J.; Permezel, M.; Rice, G.E.; Lappas, M. The effect of pre-existing maternal obesity on the placental proteome: Two-dimensional difference gel electrophoresis coupled with mass spectrometry. J. Mol. Endocrinol. 2012, 48, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.J.; Du, M.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity up-regulates inflammatory signaling pathways and enhances cytokine expression in the mid-gestation sheep placenta. Placenta 2010, 31, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.P.; Caton, J.S.; Redmer, D.A.; Grazul-Bilska, A.T.; Vonnahme, K.A.; Borowicz, P.P.; Luther, J.S.; Wallace, J.M.; Wu, G.; Spencer, T.E. Evidence for altered placental blood flow and vascularity in compromised pregnancies. J. Physiol. 2006, 572, 51–58. [Google Scholar] [CrossRef]
- Kim, J.S.; Yang, X.; Pangeni, D.; Baidoo, S.K. Relationship between backfat thickness of sows during late gestation and reproductive efficiency at different parities. Acta Agric. Scand. A Anim. Sci. 2015, 65, 1–8. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Xu, T.; Cai, A.L.; Wu, Y.H.; Wei, H.K.; Jiang, S.W.; Peng, J. Excessive backfat of sows at 109 d of gestation induces lipotoxic placental environment and is associated with declining reproductive performance. J. Anim. Sci. 2018, 96, 250–257. [Google Scholar] [CrossRef]
- Sulabo, R.C.; Jacela, J.Y.; Tokach, M.D.; Dritz, S.S.; Goodband, R.D.; DeRouchey, J.M.; Nelssen, J.L. Effects of lactation feed intake and creep feeding on sow and piglet performance. J. Anim. Sci. 2010, 88, 3145–3153. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.E.; Biensen, N.J.; Ford, S.P. Novel insight into the control of litter size in pigs, using placental efficiency as a selection tool. J. Anim. Sci. 1999, 77, 1654–1658. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Lu, J.; Deng, Z.; Xu, T.; Yang, Y.; Wei, H.; Jiang, S.; Peng, J. Maternal obesity aggravates the abnormality of porcine placenta by increasing N 6-methyladenosine. Int. J. Obes. 2018, 42, 1812. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Kwon, D.Y.; Rhee, J.S. A simple and rapid colorimetric method for determination of free fatty acids for lipase assay. J. Am. Oil Chem. Soc. 1986, 63, 89–92. [Google Scholar] [CrossRef]
- Zou, Y.; Wei, H.K.; Xiang, Q.H.; Wang, J.; Zhou, Y.F.; Peng, J. Protective effect of quercetin on pig intestinal integrity after transport stress is associated with regulation oxidative status and inflammation. J. Vet. Med. Sci. 2016, 78, 1487–1494. [Google Scholar] [CrossRef]
- Biensen, N.J.; Wilson, M.E.; Ford, S.P. The impact of either a Meishan or a Yorkshire uterus on Meishan or Yorkshire fetal and placental development to days 70, 90, and 110 of gestation. J. Anim. Sci. 1998, 76, 2169–2176. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Ren, J.; Song, T.X.; Peng, J.; Wei, H.K. Methionine Regulates mTORC1 via the T1R1/T1R3-PLCβ-Ca2+-ERK1/2 Signal Transduction Process in C2C12 Cells. Int. J. Mol. Sci. 2016, 17, 1684. [Google Scholar] [CrossRef]
- Herrera, E.; Ortega-Senovilla, H. Lipid metabolism during pregnancy and its implications for fetal growth. Curr. Pharm. Biotechnol. 2014, 15, 24–31. [Google Scholar] [CrossRef]
- Jensen, M.D. Role of body fat distribution and the metabolic complications of obesity. J. Clin. Endocrinol. Metab. 2008, 93, 57–63. [Google Scholar] [CrossRef]
- Jarvie, E.; Hauguel-de-Mouzon, S.; Nelson, S.M.; Sattar, N.; Catalano, P.M.; Freeman, D.J. Lipotoxicity in obese pregnancy and its potential role in adverse pregnancy outcome and obesity in the offspring. Clin. Sci 2010, 119, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, B.; Gogg, S.; Hedjazifar, S.; Jenndahl, L.; Hammarstedt, A.; Smith, U. Inflammation and impaired adipogenesis in hypertrophic obesity in man. Am. J. Physiol. Endocrinol. Metab. 2009, 297, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rovira, L.; Tarrade, A.; Astiz, S.; Mourier, E.; Perez-Solana, M.; de la Cruz, P.; Gomez-Fidalgo, E.; Sanchez-Sanchez, R.; Chavatte-Palmer, P.; Gonzalez-Bulnes, A. Sex and breed-dependent organ development and metabolic responses in foetuses from lean and obese/leptin resistant swine. PLoS ONE 2013, 8, e66728. [Google Scholar] [CrossRef] [PubMed]
- Adam-Vizi, V.; Chinopoulos, C. Bioenergetics and the formation of mitochondrial reactive oxygen species. Trends Pharmacol. Sci. 2006, 27, 639–645. [Google Scholar] [CrossRef]
- Bilodeau, J.F. Review: Maternal and placental antioxidant response to preeclampsia—Impact on vasoactive eicosanoids. Placenta 2014, 35, S32–S38. [Google Scholar] [CrossRef]
- Araujo, J.R.; Ramalho, C.; Correia-Branco, A.; Faria, A.; Ferraz, T.; Keating, E.; Martel, F. A parallel increase in placental oxidative stress and antioxidant defenses occurs in pre-gestational type 1 but not gestational diabetes. Placenta 2013, 34, 1095–1098. [Google Scholar] [CrossRef]
- Challier, J.C.; Basu, S.; Bintein, T.; Minium, J.; Hotmire, K.; Catalano, P.M.; Hauguel-de Mouzon, S. Obesity in pregnancy stimulates macrophage accumulation and inflammation in the placenta. Placenta 2008, 29, 274–281. [Google Scholar] [CrossRef]
- Shankar, K.; Zhong, Y.; Kang, P.; Lau, F.; Blackburn, M.L.; Chen, J.R.; Borengasser, S.J.; Ronis, M.J.; Badger, T.M. Maternal obesity promotes a proin flammatory signature in rat uterus and blastocyst. Endocrinology 2011, 152, 4158–4170. [Google Scholar] [CrossRef]
- Huang, L.; Liu, J.; Feng, L.; Chen, Y.; Zhang, J.; Wang, W. Maternal prepregnancy obesity is associated with higher risk of placental pathological lesions. Placenta 2014, 35, 563–569. [Google Scholar] [CrossRef]
- Steller, J.G.; Alberts, J.G.; Ronca, A.E. Oxidative Stress as Cause, Consequence, or Biomarker of Altered Female Reproduction and Development in the Space Environment. Int. J. Mol. Sci. 2018, 19, 3729. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Backfat Thickness (mm) of Sows at 109 Day of Gestation | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ≤16 | 17–18 | 19–20 | 21–22 | 23–24 | ≥25 | L 3 | Q 4 | ||
| Number, n | 10 | 12 | 12 | 11 | 12 | 10 | |||
| Triglyceride, mmol/gprot | 0.17 | 0.16 | 0.17 | 0.18 | 0.22 | 0.25 | 0.01 | 0.02 | 0.15 |
| Cholesterol, mmol/gprot | 0.11 | 0.10 | 0.10 | 0.13 | 0.15 | 0.19 | 0.01 | 0.02 | 0.32 |
| LDL-C 1, μmol/gprot | 25.65 | 27.42 | 27.53 | 32.84 | 38.03 | 42.45 | 0.52 | 0.01 | 0.52 |
| FFA 2, ng/mgprot | 96.18 | 94.94 | 96.07 | 99.32 | 105.66 | 119.24 | 1.26 | 0.03 | 0.37 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Xu, T.; Wu, Y.; Wei, H.; Peng, J. Oxidative Stress and Inflammation in Sows with Excess Backfat: Up-Regulated Cytokine Expression and Elevated Oxidative Stress Biomarkers in Placenta. Animals 2019, 9, 796. https://doi.org/10.3390/ani9100796
Zhou Y, Xu T, Wu Y, Wei H, Peng J. Oxidative Stress and Inflammation in Sows with Excess Backfat: Up-Regulated Cytokine Expression and Elevated Oxidative Stress Biomarkers in Placenta. Animals. 2019; 9(10):796. https://doi.org/10.3390/ani9100796
Chicago/Turabian StyleZhou, Yuanfei, Tao Xu, Yinghui Wu, Hongkui Wei, and Jian Peng. 2019. "Oxidative Stress and Inflammation in Sows with Excess Backfat: Up-Regulated Cytokine Expression and Elevated Oxidative Stress Biomarkers in Placenta" Animals 9, no. 10: 796. https://doi.org/10.3390/ani9100796
APA StyleZhou, Y., Xu, T., Wu, Y., Wei, H., & Peng, J. (2019). Oxidative Stress and Inflammation in Sows with Excess Backfat: Up-Regulated Cytokine Expression and Elevated Oxidative Stress Biomarkers in Placenta. Animals, 9(10), 796. https://doi.org/10.3390/ani9100796