A Review of Biomechanical Gait Classification with Reference to Collected Trot, Passage and Piaffe in Dressage Horses


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Diagonally Coordinated Gaits of Dressage Horses
3. Spatiotemporal Variables
3.1. Footfall Sequence and Timing
3.2. Footfall Sequence and Timing
3.3. Limb Phase
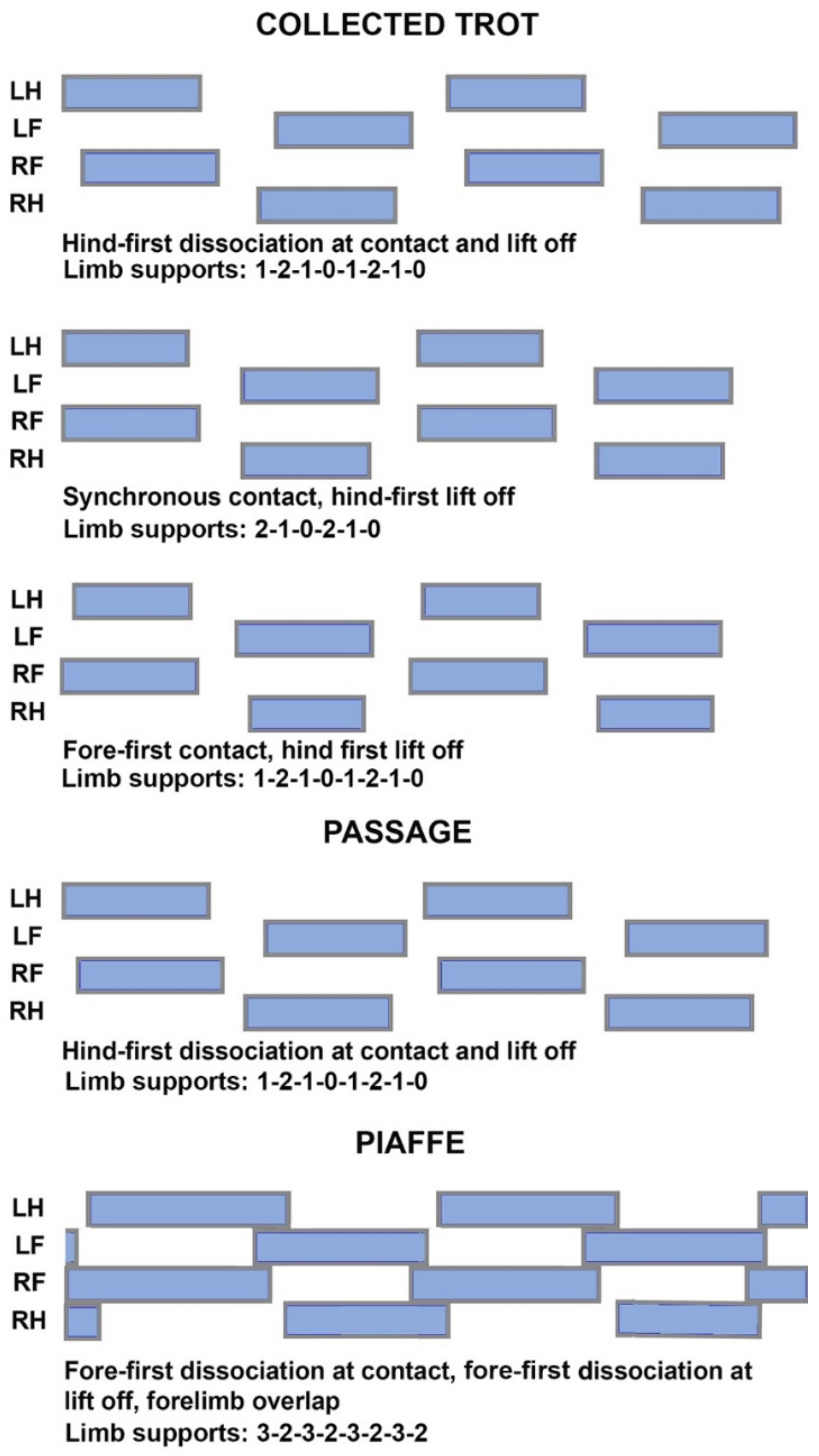
3.4. Limb Support Sequence
3.5. Aerial Phases
3.6. Kinematics of Collected Trot, Passage and Piaffe
4. Ground Reaction Forces
4.1. Vertical Ground Reaction Force
4.2. Longitudinal Ground Reaction Force
4.3. Ground Reaction Forces of Collected Trot, Passage and Piaffe
5. Gait Mechanics
5.1. Inverted Pendulum Mechanics
5.2. Spring-Mass Mechanics
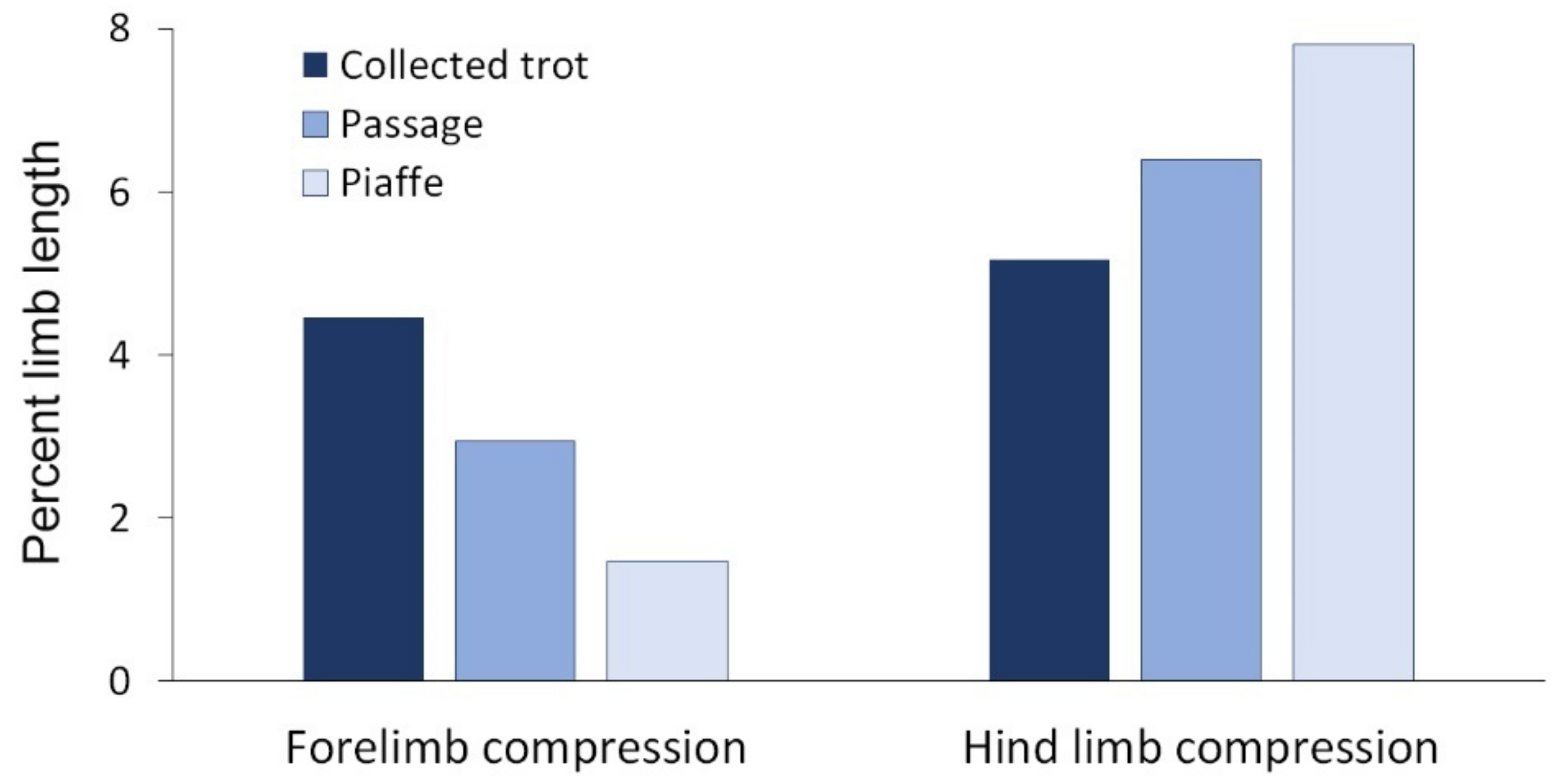
5.3. Center of Mass Mechanics in Collected Trot, Passage and Piaffe
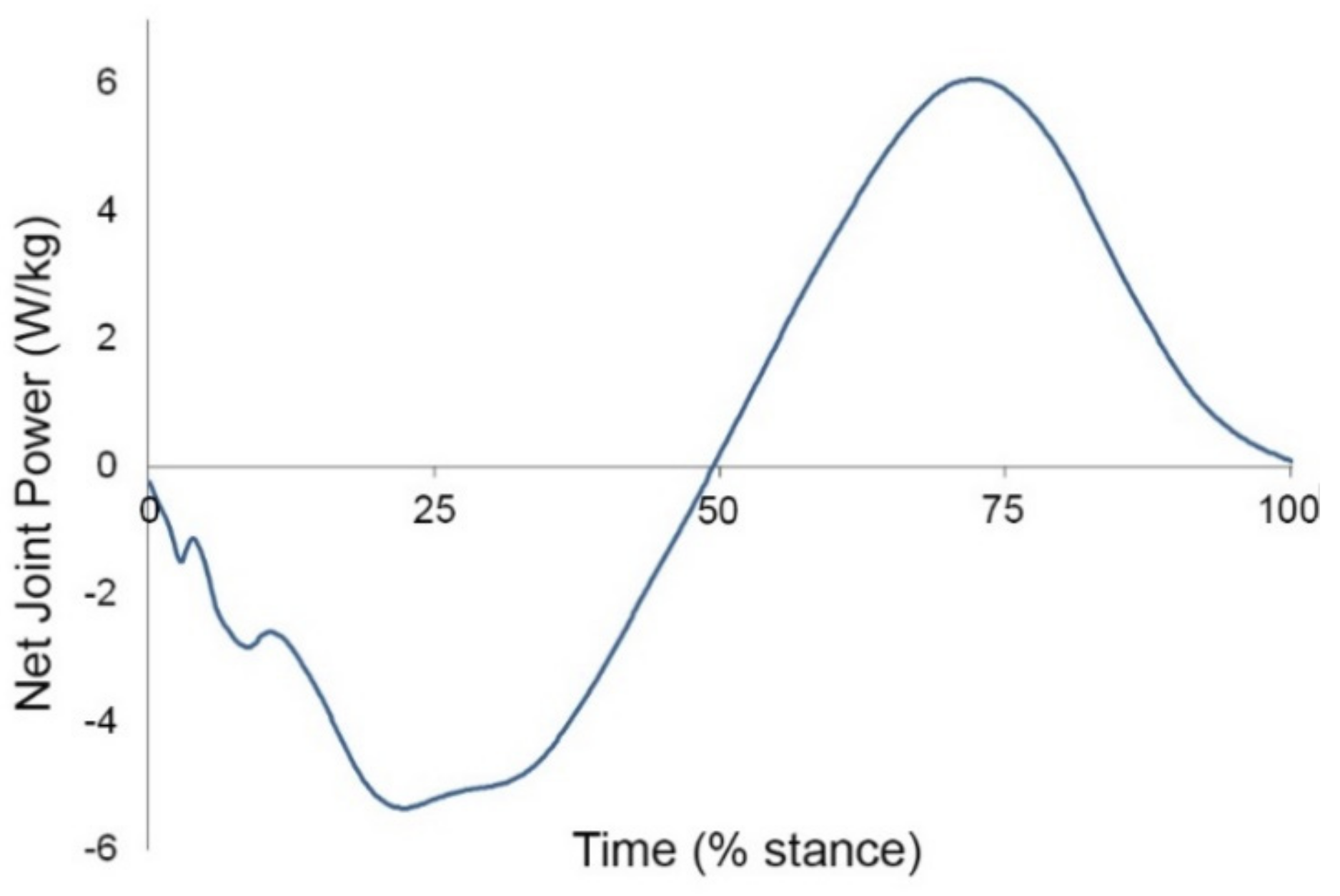
5.4. Equine Limb Mechanics
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hoyt, D.F.; Taylor, C.R. Gait and the energetics of locomotion in horses. Nature 1981, 292, 239–240. [Google Scholar] [CrossRef]
- Farley, C.; Taylor, C. A mechanical trigger for the trot-gallop transition in horses. Science 1991, 253, 306–308. [Google Scholar] [CrossRef] [PubMed]
- Granatosky, M.C.; Bryce, C.M.; Hanna, J.; Fitzsimons, A.; Laird, M.F.; Stilson, K.; Wall, C.E.; Ross, C.F. Inter-stride variability triggers gait transitions in mammals and birds. Proc. R. Soc. 2018, 285, 20181766. [Google Scholar] [CrossRef] [PubMed]
- Kristjansson, T.; Bjornsdottir, S.; Sigurdsson, A.; Andersson, L.S.; Lindgren, G.; Helyar, S.J.; Klonowski, A.M.; Arnason, T. The effect of the ‘Gait keeper’ mutation in the DMRT3 gene on gaiting ability in Icelandic horses. J. Anim. Breed Genet. 2014, 131, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Clayton, H.M. Comparison of the stride kinematics of the collected, medium, and extended walks in horses. Am. J. Vet. Res. 1995, 56, 849–852. [Google Scholar] [PubMed]
- Clayton, H.M. Comparison of the stride kinematics of the collected, working, medium and extended trot in horses. Equine Vet. J. 1994, 26, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Drevemo, S.; Dalin, G.; Fredricson, I.; Hjerten, G. Equine locomotion: I. The analysis of linear and temporal stride characteristics of trotting Standardbreds. Equine. Vet. J. 1980, 12, 60–65. [Google Scholar] [CrossRef]
- Drevemo, S.; Dalin, G.; Fredricson, I.; Björne, K. Equine locomotion: 3. The reproducibility of gait in Standardbred trotters. Equine Vet. J. 1980, 12, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.D.; Neal, R.J.; Howard, A.; Groendyk, S. The gait of pacers: 1 Kinematics of the racing stride. Equine Vet. J. 1988, 20, 341–346. [Google Scholar] [CrossRef]
- Nicodemus, M.C.; Clayton, H.M. Temporal variables of four-beat, stepping gaits of gaited horses. Appl Anim. Behav. Sci. 2003, 80, 133–142. [Google Scholar] [CrossRef]
- Clayton, H.M. Comparison of the collected, working, medium, and extended canters. Equine Vet. J. Suppl. 1994, 17, 16–19. [Google Scholar] [CrossRef]
- Deuel, N.R.; Lawrence, L.M. Gallop velocity and limb contact variables of Quarter Horses. J. Equine Vet. Sci. 1986, 6, 143–147. [Google Scholar] [CrossRef]
- Cavagna, G.A.; Heglund, N.C.; Taylor, C.R. Mechanical work in terrestrial locomotion: Two basic mechanisms for minimizing energy expenditure. Am. J. Physiol. 1977, 233, 243–261. [Google Scholar] [CrossRef] [PubMed]
- Biewener, A.A. Patterns of mechanical energy change in tetrapod gait: Pendula, springs and work. J. Exp. Zool 2006, 305A, 899–911. [Google Scholar] [CrossRef]
- Biknevicius, A.R.; Mullineaux, D.R.; Clayton, H.M. Locomotor mechanics of the tölt in Icelandic horses: Taking the walk for a run. Am. J. Vet. Res. 2006, 67, 1505–1510. [Google Scholar] [CrossRef]
- Hildebrand, M. Symmetrical gaits of horses. Science 1965, 150, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Biknevicius, A.; Mullineau, D.R.; Clayton, H.M. Ground reaction forces and limb function in tölting Icelandic horses. Equine Vet. J. 2004, 36, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Merkens, H.W.; Schamhardt, H.C.; Hartman, W.; Kersjes, A.W. Ground reaction force patterns of Dutch Warmblood horses at normal walk. Equine Vet. J. 1985, 18, 207–214. [Google Scholar] [CrossRef]
- Robilliard, J.J.; Pfau, T.; Wilson, A.M. Gait characterisation and classification in horses. J. Exp. Biol. 2007, 210, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Starke, S.D.; Weller, R.; Wilson, A.M.; Pfau, T. Walk–run classification of symmetrical gaits in the horse: A multidimensional approach. J. R. Soc. Interface 2009, 6, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, S.J.; Clayton, H.M. Collisional mechanics of the diagonal gaits of horses over a range of speeds. Peer J. 2019, 7, e7689. [Google Scholar] [CrossRef] [PubMed]
- Rules for Dressage. Fédération Equestre Internationale 2019, 25th ed. Available online: https://inside.fei.org/fei/disc/dressage/rules (accessed on 2 October 2019).
- Holmström, M.; Fredricson, I.; Drevemo, S. Biokinematic analysis of the Swedish Warmblood riding horse trot. Equine Vet. J. 1993, 26, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Back, W.; Schamhardt, H.C.; Hartman, W.; Barneveld, A. Kinematic differences between the distal portions of the forelimbs and hind limbs of horses at the trot. Am. J. Vet. Res. 1995, 56, 1522–1528. [Google Scholar]
- Clayton, H.M. Classification of collected trot, passage and piaffe using stance phase temporal variables. Equine Vet. J. Suppl. 1997, 23, 54–57. [Google Scholar]
- Deuel, N.R.; Park, J.J. The gait patterns of Olympic dressage horses. Int. J. Sports Biomech. 1990, 6, 198–226. [Google Scholar] [CrossRef]
- Holmström, M.; Fredricson, I.; Drevemo, S. Biokinematic differences between riding horses judged as good and poor at the trot. Equine Vet. J. 1994, 26, 51–56. [Google Scholar] [CrossRef]
- Weishaupt, M.A.; Byström, A.; Von Peinen, K.; Wiestner, T.; Meyer, H.; Waldern, N.; Johnston, C.; Van Weeren, R.; Roepstorff, L. Kinetics and kinematics of the passage. Equine Vet. J. 2009, 41, 263–267. [Google Scholar] [CrossRef]
- Hobbs, S.J.; Bertram, J.; Clayton, H.M. An exploration of the influence of diagonal dissociation and moderate changes in speed on locomotor parameters in trotting horses. Peer J. 2016, 4, e2190. [Google Scholar] [CrossRef]
- Demes, B.; Larson, S.G.; Stern, J.T., Jr.; Jungers, W.L.; Biknevicius, A.R.; Schmidtt, D. The kinetics of primate quadrupedalism: “hindlimb drive” reconsidered. J. Hum. Evol. 1994, 26, 353–374. [Google Scholar] [CrossRef]
- Back, W.; Hartmann, W.M.; Schamhardt, H.C.; Bruin, G.; Barneveld, A. Kinematic response to a 70-day training period in trotting Dutch Warmbloods. Equine Vet. J. 1995, 18, 127–131. [Google Scholar] [CrossRef]
- Hildebrand, M. Analysis of tetrapod gaits: General considerations and symmetrical gaits. In Neural Control of Locomotion; Herman, R.M., Grillner, S., Stein, P.S.G., Stuart, D.G., Eds.; Plenum Press: New York, NY, USA, 1976; pp. 203–236. [Google Scholar]
- Hildebrand, M. Walking and running. In Functional Vertebrate Morphology; Hildebrand, M., Bramble, D.M., Liem, K.F., Wake, D.B., Eds.; Belknap: Harvard, Cambridge, London, UK, 1985; pp. 38–57. [Google Scholar]
- Nicodemus, N.C.; Booker, J.E. Two-dimensional kinematics of the jog and lope of the stock breed western pleasure horse. Equine Compar. Exerc. Physiol. 2007, 4, 59–70. [Google Scholar] [CrossRef]
- Holmström, M.; Fredricson, I.; Drevemo, S. Biokinematic effects of collection in the elite dressage trot. Equine Vet. J. 1995, 27, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Weishaupt, M.A.; Hogg, H.P.; Auer, J.A.; Wiestner, T. Velocity-dependent changes of time, force and spatial parameters in Warmblood horses walking and trotting on a treadmill. Equine Vet. J. 2010, 42, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Clayton, H.M.; Hobbs, S.J. An exploration of strategies used by dressage horses to control moments around the center of mass when performing passage. Peer J. 2017, 5, e3866. [Google Scholar] [CrossRef]
- Clayton, H.M.; Hobbs, S.J. Ground reaction forces: The sine qua non of legged locomotion. J. Equine Vet. Sci. 2019, 76, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.M. A model of bipedal locomotion on compliant legs. Phil. Trans. R. Soc. Lond 1992, 338, 189–198. [Google Scholar]
- Merkens, H.W.; Schamhardt, H.C.; Osch, G.J.V.M.; van den Bogert, A.J. Ground reaction force patterns of Dutch Warmblood horses at normal trot. Equine Vet. J. 1993, 25, 134–137. [Google Scholar] [CrossRef]
- Hobbs, S.J.; Richards, J.; Clayton, H.M. The effect of centre of mass location on sagittal plane moments around the centre of mass in trotting horses. J. Biomech. 2014, 47, 1278–1286. [Google Scholar] [CrossRef]
- Dutto, D.J.; Hoyt, D.F.; Cogge, E.A.; Wickler, S.J. Ground reaction forces in horses trotting up an incline and on the level over a range of speeds. J. Exp. Biol. 2004, 207, 3507–3514. [Google Scholar] [CrossRef]
- Hobbs, S.J.; Clayton, H.M. Sagittal plane ground reaction forces, center of pressure and center of mass in trotting horses. Vet. J. 2013, 198, e14–e19. [Google Scholar] [CrossRef]
- Herr, H.M.; McMahon, T.A. A galloping horse model. Int. J. Robotics Res. 2001, 20, 26–37. [Google Scholar] [CrossRef]
- Clayton, H.M.; Schamhardt, H.C.; Hobbs, S.J. Ground reaction forces of elite dressage horses in collected trot and passage. Vet. J. 2017, 221, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Buchner, H.H.F.; Obermüller, S.; Scheidl, M. Body centre of mass movement in the sound horse. Vet. J. 2000, 160, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Merkens, H.W.; Schamhardt, H.C. Distribution of ground reaction forces of the concurrently loaded limbs of the Dutch Warmblood horse at the normal walk. Equine Vet. J. 1988, 20, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Griffin, T.M.; Main, R.P.; Farley, C.T. Biomechanics of quadrupedal walking: How do four-legged animals achieve inverted pendulum-like mechanics? J. Exp. Biol. 2004, 207, 3545–3558. [Google Scholar] [CrossRef] [PubMed]
- Kuo, A.D. Energetics of actively powered locomotion using the simplest walking model. J. Biomech. Eng. 2002, 124, 113–120. [Google Scholar] [CrossRef]
- Alexander, R.M. Why animals gallop. Am. Zool. 1988, 28, 237–245. [Google Scholar] [CrossRef]
- Dimery, N.J.; Alexander, R.M.; Ker, R.F. Elastic extension of leg tendons in the locomotion of horses (Equus caballus). J. Zool. 1986, 210, 415–425. [Google Scholar] [CrossRef]
- Minetti, A.E.; Ardigò, L.P.; Reinach, E.; Saibene, F. The relationship between mechanical work and energy expenditure of locomotion in horses. J. Exp. Biol. 1999, 202, 2329–2338. [Google Scholar]
- Wilson, A.M.; Watson, J.C.; Litchwark, G.A. A catapult action for rapid limb protraction. Nature 2003, 421, 35. [Google Scholar] [CrossRef]
- Clayton, H.M.; Lanovaz, J.L.; Schamhardt, H.C.; Willemen, M.A.; Colborne, G.L. Net joint moments and powers in the equine forelimb in the stance phase of the trot. Equine Vet. J. 1998, 30, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Biewener, A.A. Muscle-tendon stresses and elastic energy storage during locomotion in the horse. Compar Biochem. Physiol. Part. B. 1998, 120, 73–87. [Google Scholar] [CrossRef]
- Clayton, H.M.; Sha, D.H.; Stick, J.A.; Mullineaux, D.R. Three dimensional kinematics of the equine carpus at trot. Equine Vet. J. 2004, 36, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Saunders, J.B.M.; Inman, V.T.; Eberhart, H.D. The major determinants in normal and pathological gait. J. Bone Joint Surg. 1953, 35A, 543–558. [Google Scholar] [CrossRef]
- Farley, D.T.; Glasheen, J.; McMahon, T.A. Running springs: Speed and animal size. J. Exp. Biol. 1993, 185, 71–86. [Google Scholar] [PubMed]
- McMahon, T.A.; Valiant, G.; Frederick, E.C. Groucho running. J. Appl. Physiol. 1987, 62, 2326–2337. [Google Scholar] [CrossRef] [PubMed]
- Biknevicius, A.M.; Reilly, S. Correlation of symmetrical gaits and whole body mechanics: Debunking myths in locomotor biodynamics. J. Exp. Biol. 2006, 305A, 923–934. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clayton, H.M.; Hobbs, S.J. A Review of Biomechanical Gait Classification with Reference to Collected Trot, Passage and Piaffe in Dressage Horses. Animals 2019, 9, 763. https://doi.org/10.3390/ani9100763
Clayton HM, Hobbs SJ. A Review of Biomechanical Gait Classification with Reference to Collected Trot, Passage and Piaffe in Dressage Horses. Animals. 2019; 9(10):763. https://doi.org/10.3390/ani9100763
Chicago/Turabian StyleClayton, Hilary M., and Sarah Jane Hobbs. 2019. "A Review of Biomechanical Gait Classification with Reference to Collected Trot, Passage and Piaffe in Dressage Horses" Animals 9, no. 10: 763. https://doi.org/10.3390/ani9100763
APA StyleClayton, H. M., & Hobbs, S. J. (2019). A Review of Biomechanical Gait Classification with Reference to Collected Trot, Passage and Piaffe in Dressage Horses. Animals, 9(10), 763. https://doi.org/10.3390/ani9100763