Anthropogenic Food Subsidy to a Commensal Carnivore: The Value and Supply of Human Faeces in the Diet of Free-Ranging Dogs



Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
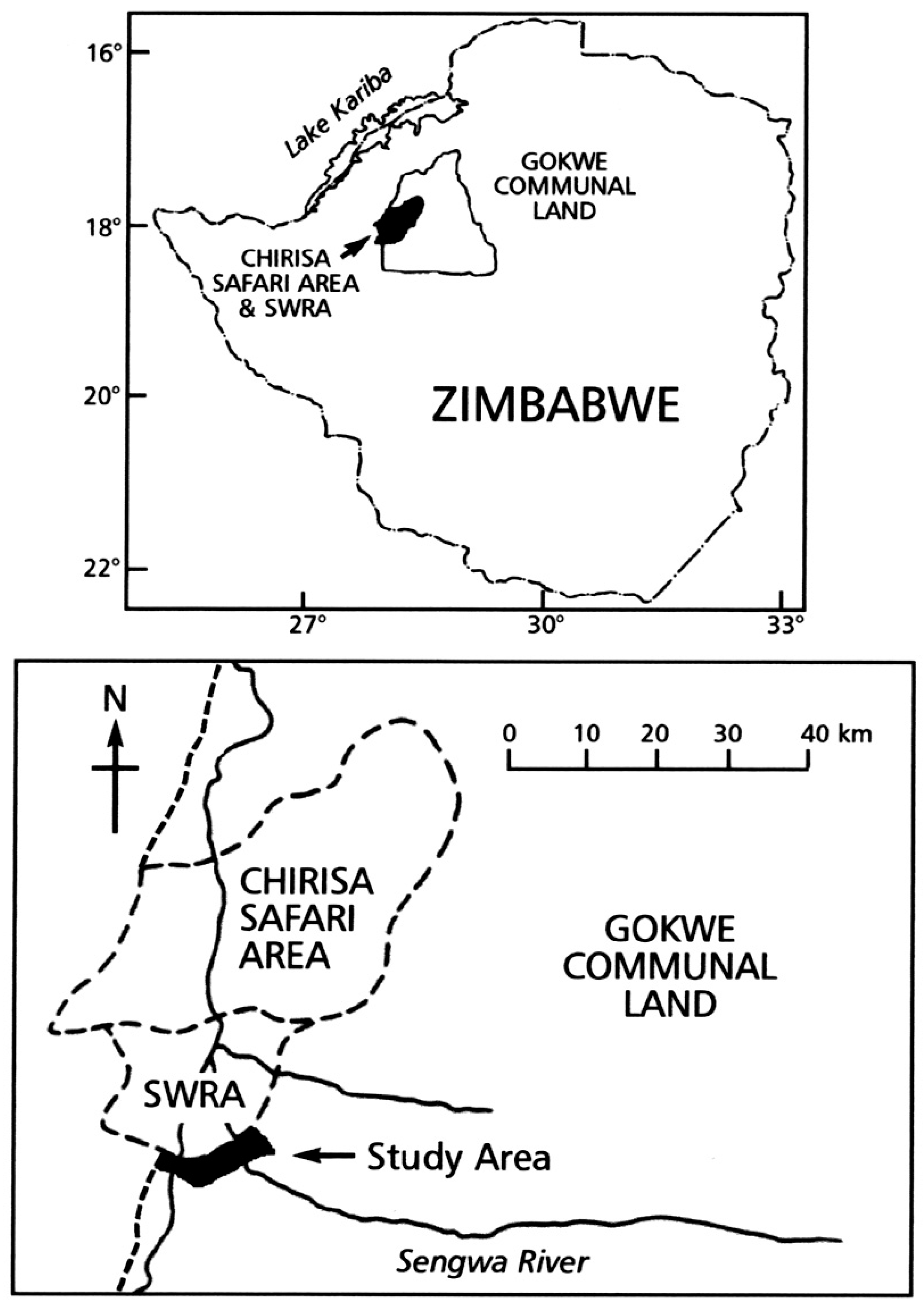
2.1. Study Area
2.2. Livelihoods and Waste Management
2.3. Dog Population and Ecology
2.4. Methods
2.4.1. Dietary Analysis
2.4.2. Nutritional Value of Food Items
2.4.3. Dog Condition
2.4.4. Sociology and Diet
2.4.5. Free and Prior Informed Consent
3. Results
3.1. Diet
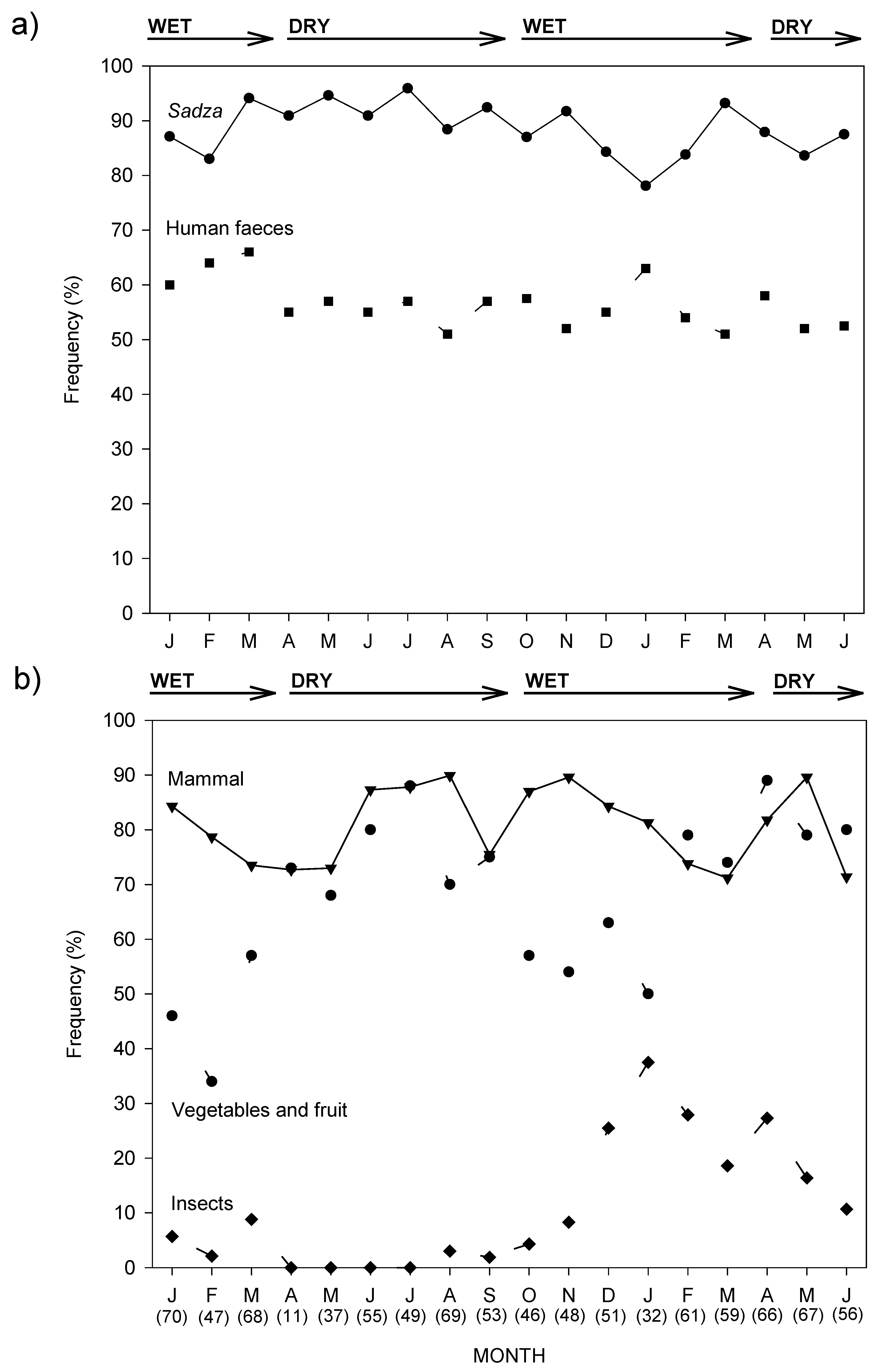
3.1.1. Scat Analysis
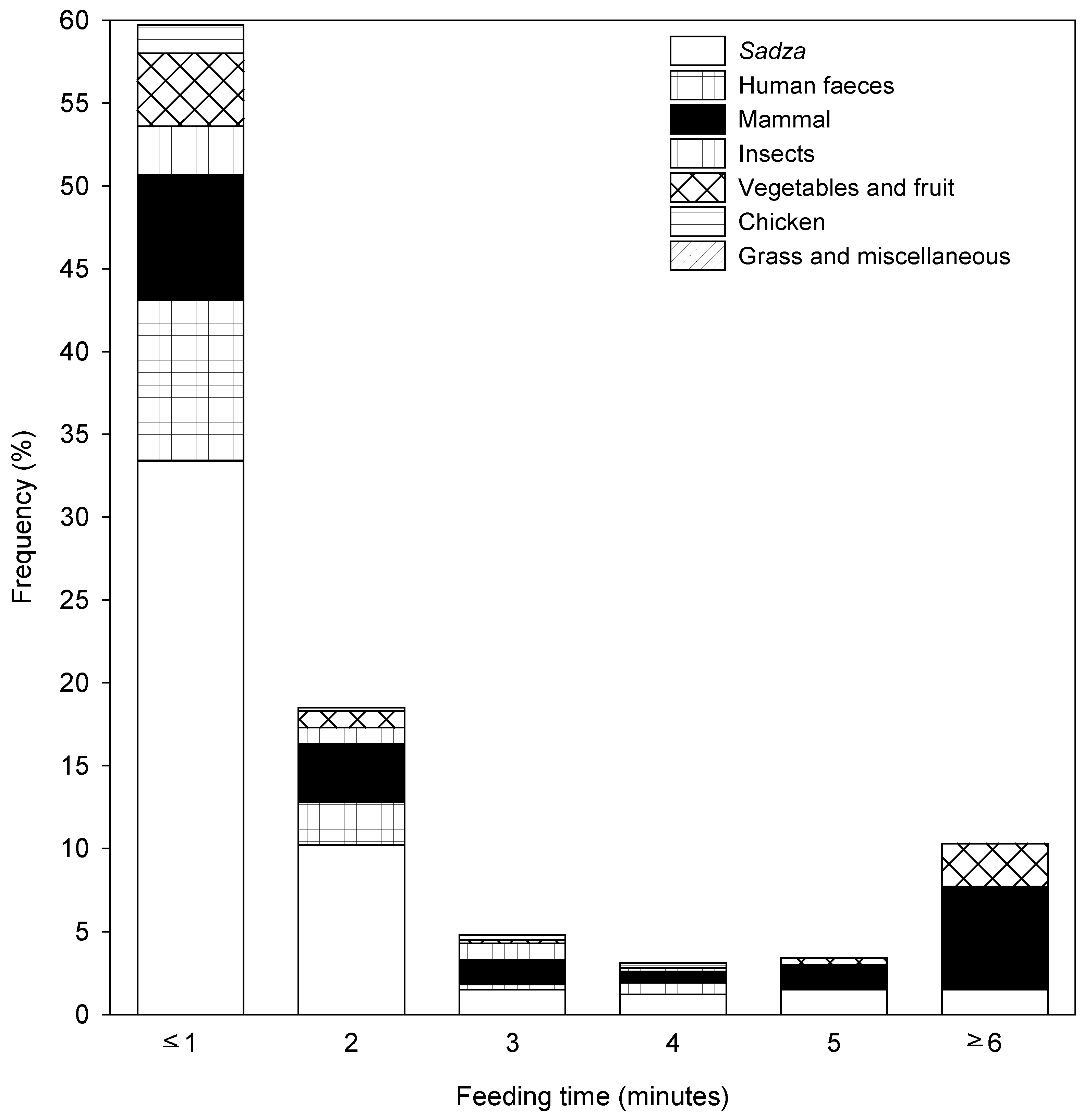
3.1.2. Observed Diet
3.2. Nutritional Value of Food Items
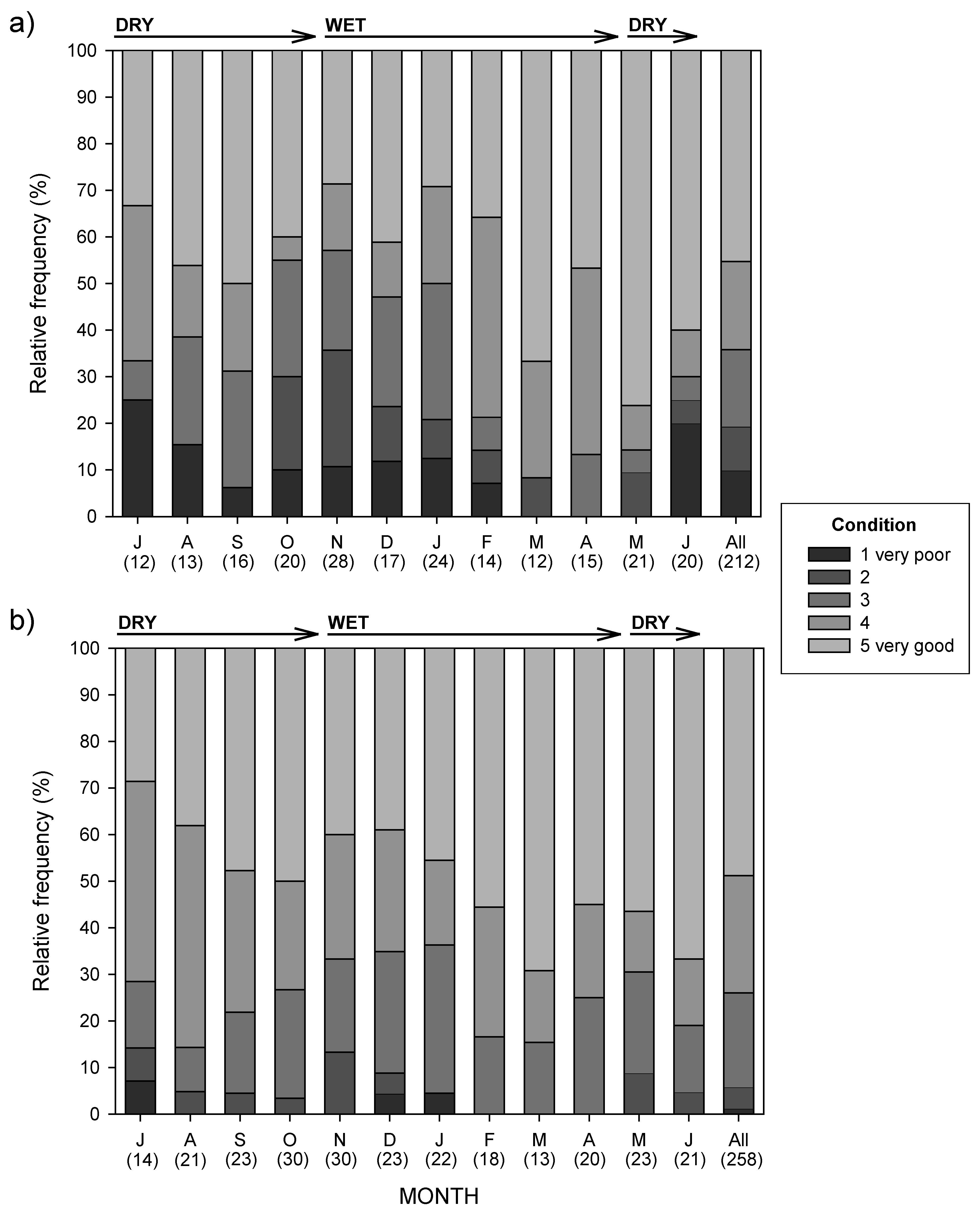
3.3. Dog Condition
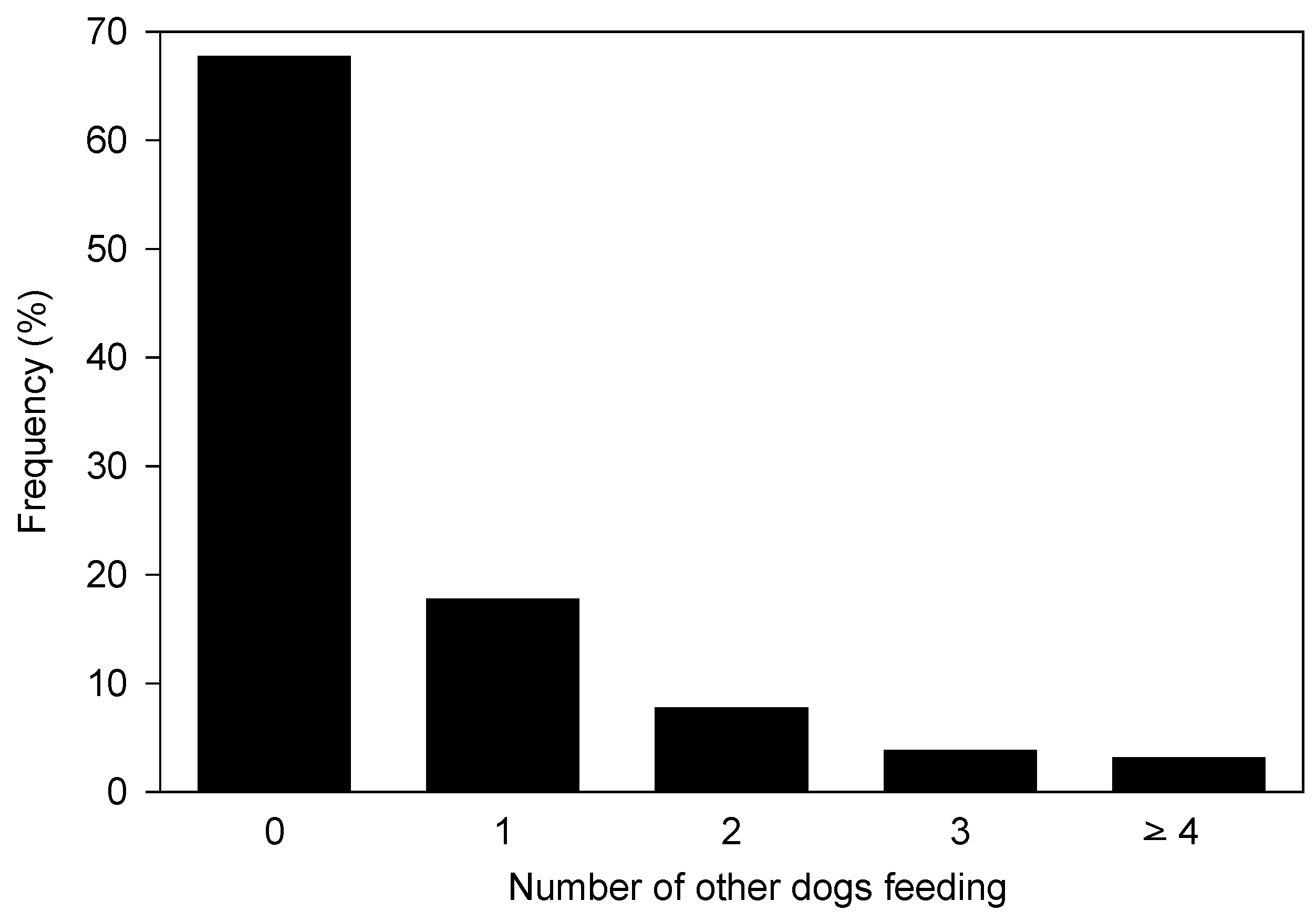
3.4. Sociology and Diet
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hughes, J.; Macdonald, D.W. A review of the interactions between free-roaming domestic dogs and wildlife. Biol. Conserv. 2013, 157, 341–351. [Google Scholar] [CrossRef]
- Butler, J.R.A.; Linnell, J.D.C.; Morrant, D.; Athreya, V.; Lescureux, N.; McKeown, A. Dog eat dog, cat eat dog: Social-ecological dimensions of dog predation by wild carnivores. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 117–143. [Google Scholar]
- WHO. World Health Organization Expert Consultation on Rabies; World Health Organization Technical Report Series; WHO: Geneva, Switzerland, 2005; Volume 931, pp. 1–88. [Google Scholar]
- Smout, F.A.; Skerratt, L.F.; Butler, J.R.A.; Johnson, C.N.; Congdon, B.C.; Thompson, R.C.A. The hookworm Ancylostoma ceylanicum: An emerging public health risk in Australian tropical rainforests and Indigenous communities. One Health 2017, 3, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Gompper, M.E. The dog-human-wildlife interface: Assessing the scope of the problem. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 9–54. [Google Scholar]
- Butler, J.R.A.; du Toit, J.T. Diet of free-ranging domestic dogs (Canis familiaris) in rural Zimbabwe: Implications for wild scavengers on the periphery of wildlife reserves. Anim. Conserv. 2004, 5, 29–37. [Google Scholar] [CrossRef]
- Vanak, A.T.; Dickman, C.R.; Silva-Rodriguez, E.A.; Butler, J.R.A.; Ritchie, E.G. Top-dogs and under-dogs: Competition between dogs and sympatric carnivores. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 69–93. [Google Scholar]
- Knobel, D.L.; Butler, J.R.A.; Lembo, T.; Critchlow, R.; Gompper, M.E. Dogs, disease, and wildlife. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 144–169. [Google Scholar]
- Butler, J.R.A.; du Toit, J.T.; Bingham, J. Free-ranging domestic dogs Canis familiaris as predators and prey in rural Zimbabwe: Threats of competition and disease to large wild carnivores. Biol. Conserv. 2004, 115, 369–378. [Google Scholar] [CrossRef]
- Taylor, L.H.; Wallace, R.M.; Balaram, D.; Lindenmayer, J.M.; Eckery, D.C.; Mutonono-Watkiss, B.; Parravani, E.; Nel, L.H. The role of dog population management in rabies elimination—A review of current approaches and future opportunities. Front. Vet. Sci. 2017, 4, 109. [Google Scholar] [CrossRef] [PubMed]
- Knobel, D.L.; Cleaveland, S.; Coleman, P.G.; Fevre, E.M.; Meltzer, M.I.; Miranda, M.E.; Shaw, A.; Zinsstag, J.; Meslin, F.X. Re-evaluating the burden of rabies in Africa and Asia. Bull. World Health Organ. 2005, 83, 360–368. [Google Scholar] [PubMed]
- Ogada, D.L.; Keesing, F.; Virani, M.Z. Dropping dead: Causes and consequences of vulture population declines worldwide. Ann. N. Y. Acad. Sci. USA 2012, 1249, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Vanak, A.T.; Gompper, M.E. Dietary niche separation between sympatric free-ranging domestic dogs and Indian foxes in central India. J. Mammal. 2009, 90, 1058–1065. [Google Scholar] [CrossRef]
- Atickem, A.; Bekele, A.; Williams, S.D. Competition between domestic dogs and Ethiopian wolf (Canis simensis) in the Bale Mountains National Park, Ethiopia. Afr. J. Ecol. 2010, 48, 401–407. [Google Scholar] [CrossRef]
- Boitani, L.; Ciucci, P.; Ortolani, A. Behaviour and social ecology of free-ranging dogs. In Behavioral Biology of Dogs; Jensen, P., Ed.; CABI Publishing: Wallingford, UK, 2007; pp. 147–165. [Google Scholar]
- Sen Majumder, S.; Bhadra, A.; Ghosh, A.; Mitra, S.; Bhattacharjee, D.; Chatterjee, J.; Nandi, A.K.; Bhadra, A. To be or not to be social: Foraging associations of free-ranging dogs in an urban ecosystem. Acta Ethol. 2014, 17, 1–8. [Google Scholar] [CrossRef]
- Marshall-Pescini, S.; Cafazzo, S.; Viranyi, Z.; Range, F. Integrating social ecology in explanations of wolf–dog behavioral differences. Curr. Opin. Behav. Sci. 2017, 16, 80–86. [Google Scholar] [CrossRef]
- Butler, J.R.A. The Ecology of Domestic Dogs Canis familiaris in the Communal Lands of Zimbabwe. Ph.D. Thesis, University of Zimbabwe, Harare, Zimbabwe, 1998. [Google Scholar]
- Butler, J.R.A.; Bingham, J. Demography and dog-human relationships of the dog population in Zimbabwean communal lands. Vet. Rec. 2000, 147, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.R.A. The economic costs of wildlife predation on livestock in Gokwe communal land, Zimbabwe. Afr. J. Ecol. 2000, 38, 23–30. [Google Scholar] [CrossRef]
- Brooks, R. Survey of the dog population of Zimbabwe and its level of rabies vaccination. Vet. Rec. 1990, 127, 592–596. [Google Scholar] [PubMed]
- Bradley, M.; (National Institute of Health Research, Harare, Zimbabwe). Personal communication, 1995.
- Henschel, J.R.; Skinner, J.D. The diet of spotted hyaenas Crocuta crocuta in Kruger National Park. Afr. J. Ecol. 1990, 28, 69–82. [Google Scholar] [CrossRef]
- Sillero-Zubiri, C.; Gottelli, D. Diet and feeding behavior of Ethiopian wolves (Canis simensis). J. Mammal. 1995, 76, 531–541. [Google Scholar] [CrossRef]
- Corbett, L.K. Assessing the diet of dingoes from faeces: A comparison of 3 methods. J. Wildl. Manag. 1989, 53, 343–346. [Google Scholar] [CrossRef]
- Riney, T. A field technique for assessing physical condition of some ungulates. J. Wildl. Manag. 1960, 24, 92–94. [Google Scholar] [CrossRef]
- Chitsiku, I.C. Nutritive value of foods of Zimbabwe. Zambezia 1989, 16, 67–97. [Google Scholar]
- Oppenheimer, E.C.; Oppenheimer, J.R. Certain behavioral features in the pariah dog (Canis familiaris) in West Bengal. Appl. Anim. Ethol. 1975, 2, 81–92. [Google Scholar] [CrossRef]
- Rose, C.; Parker, A.; Jefferson, B.; Cartmell, E. The characterization of feces and urine: A review of the literature to inform advanced treatment technology. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1827–1879. [Google Scholar] [CrossRef] [PubMed]
- Scantlebury, M.; Butterwisk, R.; Speakman, J.R. Energetics of lactation in domestic dog (Canis familiaris) breeds of two sizes. Comp. Biochem. Physiol. Part A 2000, 125, 197–210. [Google Scholar] [CrossRef]
- Ontko, J.; Phillips, P.H. Reproduction and lactation studies with bitches fed semi-purified diets. J. Nutr. 1958, 65, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.L.; Calzada, J.; Saldana, A.; Santamaria, A.M.; Pineda, V.; Gonzalez, K.; Chaves, L.F.; Garner, B.; Gottdenker, N. Domestic dog health worsens with socio-economic deprivation of their home communities. Acta Trop. 2014, 135, 67–74. [Google Scholar] [CrossRef] [PubMed]
- McRee, A.; Wilkes, R.P.; Dawson, J.; Parry, R.; Foggin, C.; Adams, H.; Odoi, A.; Kennedy, M.A. Serological detection of infection with canine distemper virus, canine parvovirus and canine adenovirus in communal dogs from Zimbabwe. J. S. Afr. Vet. Assoc. 2014, 85, 1110. [Google Scholar] [CrossRef] [PubMed]
- Coppinger, R.; Coppinger, L. Dogs: A New Understanding of Canine Origin, Behavior and Evolution; The University of Chicago Press: Chicago, IL, USA, 2001. [Google Scholar]
- Graves, I.L.; Oppenheimer, J.R. Human viruses in animals in West Bengal: An ecological analysis. Hum. Ecol. 1975, 3, 105–130. [Google Scholar] [CrossRef]
- Morrant, D.S.; Wurster, C.; Johnson, C.N.; Butler, J.R.A.; Congdon, B.C. Prey use by dingoes in a contested landscape: Ecosystem service provider or biodiversity threat? Ecol. Evol. 2017, 7, 8927–8935. [Google Scholar] [CrossRef] [PubMed]
- Morrant, D.S.; Johnson, C.N.; Butler, J.R.A.; Congdon, B.C. Biodiversity friend or foe: Ecology of a top predator, the dingo in contested landscapes of the Australian Wet Tropics. Austral Ecol. 2017, 42, 252–264. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Item | Scats (%F) | Observed (%RF) |
|---|---|---|
| Sadza 1 | 87.9 | 22.1 |
| Mammal meat, bones and skin 2 | 81.3 | 48.8 |
| Vegetables and fruit 3 | 69.8 | 2.3 |
| Human faeces | 56.2 | 20.5 |
| Grass | 24.9 | 0.1 |
| Insects 4 | 11.7 | 3.1 |
| Chicken 5 | 6.7 | 1.8 |
| Food Item | Crude Protein (%) | Gross Energy (MJ/kg) |
|---|---|---|
| Human faeces | 18.7 | 18.7 |
| Sadza | 8.3 | 18.5 |
| Mammal meat, bones and skin | 13.5–30.1 | 3.5–18.8 |
| Insects | 12.9–15.1 | 5.0–5.9 |
| Vegetables and fruit | 0.5–27.0 | 0.6–24.7 |
| Chicken | 10.0–26.7 | 8.2–14.6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butler, J.R.A.; Brown, W.Y.; Du Toit, J.T. Anthropogenic Food Subsidy to a Commensal Carnivore: The Value and Supply of Human Faeces in the Diet of Free-Ranging Dogs. Animals 2018, 8, 67. https://doi.org/10.3390/ani8050067
Butler JRA, Brown WY, Du Toit JT. Anthropogenic Food Subsidy to a Commensal Carnivore: The Value and Supply of Human Faeces in the Diet of Free-Ranging Dogs. Animals. 2018; 8(5):67. https://doi.org/10.3390/ani8050067
Chicago/Turabian StyleButler, James R. A., Wendy Y. Brown, and Johan T. Du Toit. 2018. "Anthropogenic Food Subsidy to a Commensal Carnivore: The Value and Supply of Human Faeces in the Diet of Free-Ranging Dogs" Animals 8, no. 5: 67. https://doi.org/10.3390/ani8050067
APA StyleButler, J. R. A., Brown, W. Y., & Du Toit, J. T. (2018). Anthropogenic Food Subsidy to a Commensal Carnivore: The Value and Supply of Human Faeces in the Diet of Free-Ranging Dogs. Animals, 8(5), 67. https://doi.org/10.3390/ani8050067