Dog and Cat Interactions in a Remote Aboriginal Community


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
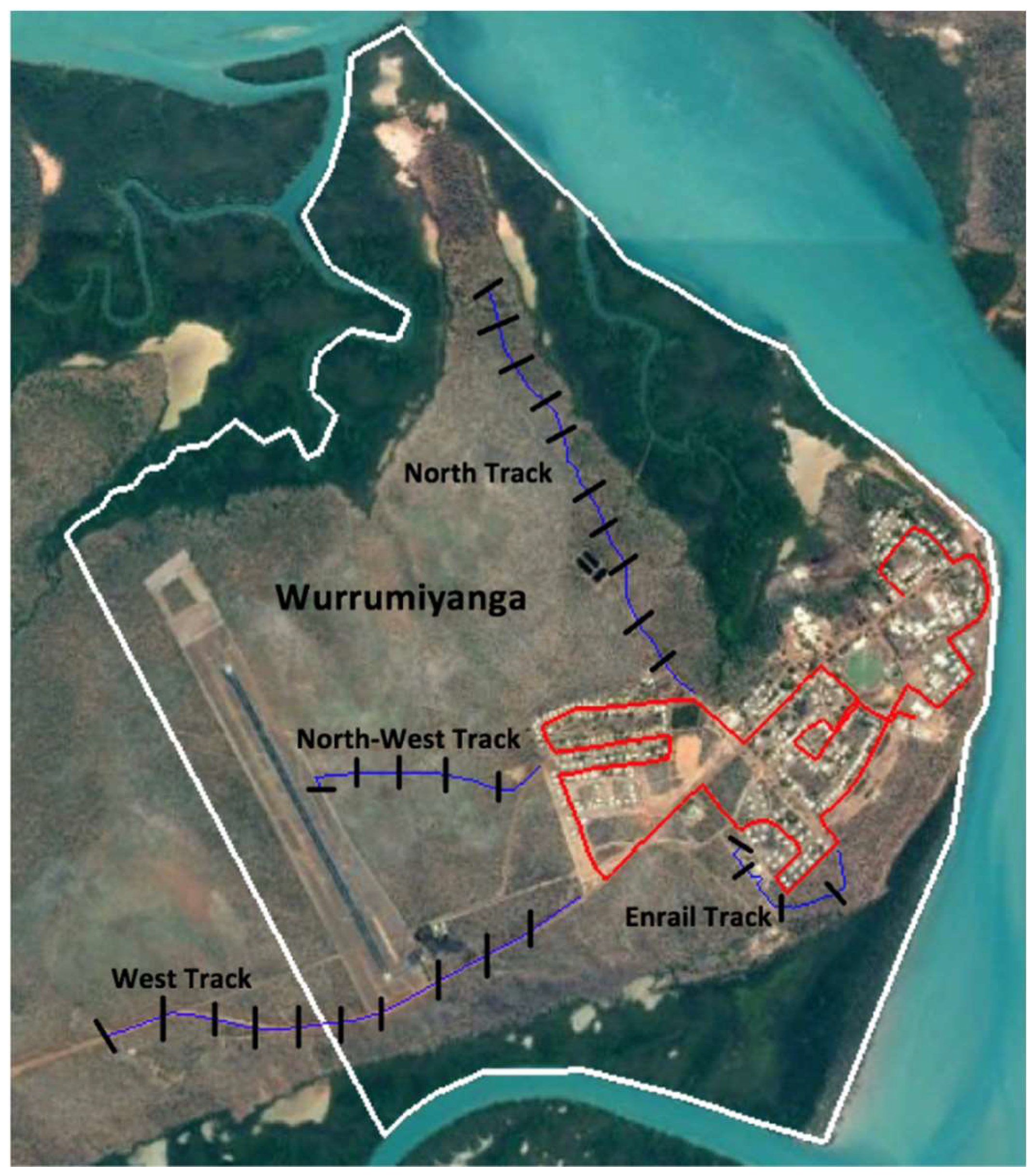
2.1. Study Site
2.2. Dog and Cat Census
2.3. Roaming Behaviours
2.3.1. Direct Observation—Transect Drives
2.3.2. Indirect Observation—Remote-Sensing Cameras
2.3.3. Tracking—GPS Loggers
2.4. Data and Statistical Analyses
3. Results
3.1. Dog and Cat Census
3.2. Roaming Behaviours
3.2.1. Direct Observation—Transect Drives
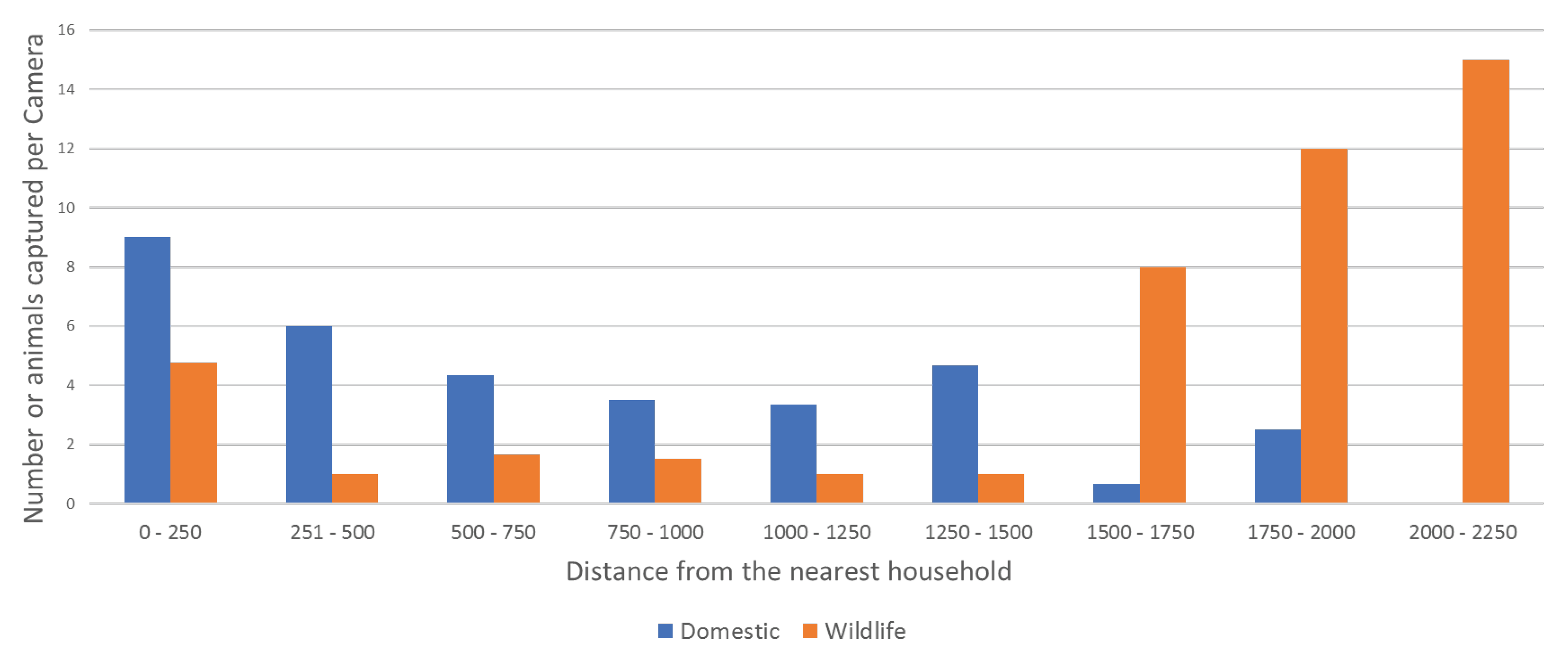
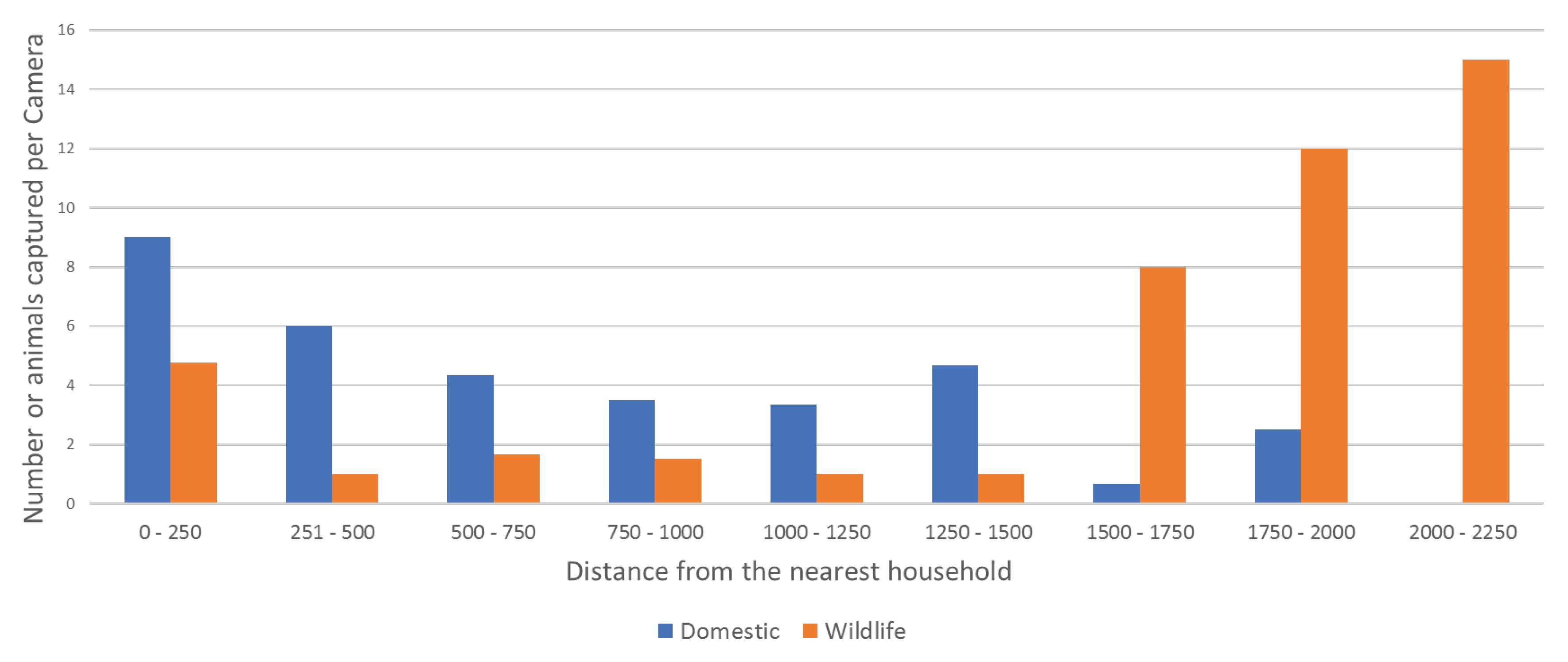
3.2.2. Indirect Observation—Remote-Sensing Cameras
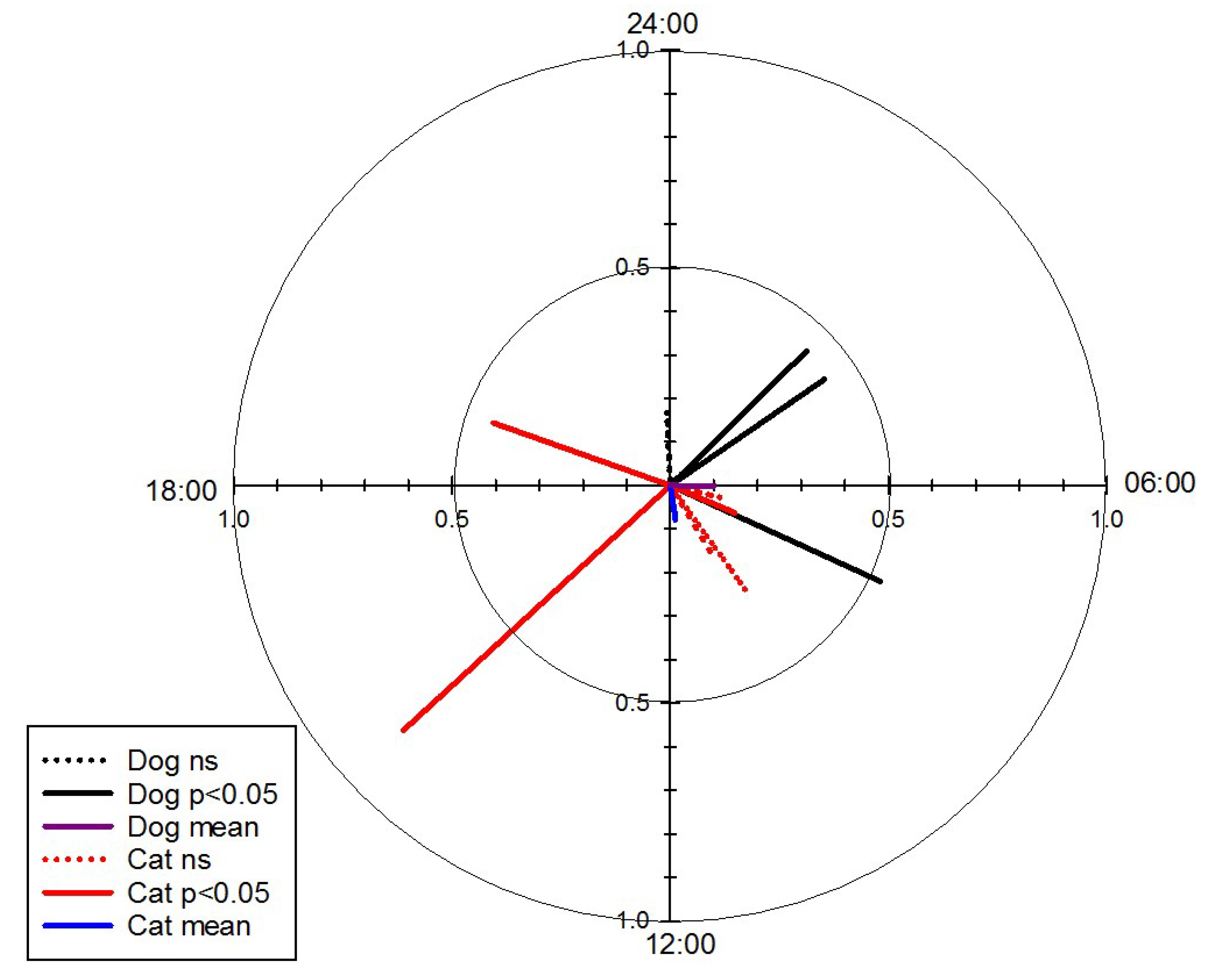
3.2.3. Tracking—GPS Loggers
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Track | Dist. (m) 1 | Camera | Night | Dog | Owner | Cat | Wallaby | Bandicoot | Possum | Curlew | Peewee | Cockatoo | Pigeon | Magpie | Unknown |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Enrail | 21 | 34 | 4 | 0 | 0 | 1 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Enrail | 46 | 29 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Enrail | 74 | 12 | 4 | 8 | 4 | 1 | 1 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 |
| Enrail | 149 | 28 | 4 | 4 | 10 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| North West | 247 | 14 | 2 | 5 | 2 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 |
| West | 419 | 4 | 6 | 8 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North West | 432 | 22 | 2 | 4 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North | 502 | 38 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| West | 543 | 35 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North West | 602 | 24 | 2 | 4 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North | 678 | 1 | 2 | 6 | 3 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| West | 776 | 11 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North West | 792 | 26 | 2 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North West | 911 | 9 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
| North | 914 | 19 | 5 | 11 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| West | 1005 | 18 | 6 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North | 1133 | 15 | 2 | 3 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| West | 1166 | 30 | 6 | 3 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| West | 1273 | 7 | 6 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North | 1358 | 31 | 2 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| West | 1411 | 40 | 6 | 6 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North | 1507 | 21 | 4 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| West | 1575 | 13 | 6 | 2 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| North | 1659 | 3 | 6 | 0 | 0 | 0 | 2 | 2 | 1 | 4 | 0 | 0 | 0 | 0 | 1 |
| North | 1804 | 25 | 5 | 0 | 0 | 0 | 7 | 0 | 2 | 14 | 0 | 0 | 0 | 0 | 5 |
| West | 1830 | 36 | 6 | 5 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| North | 2034 | 27 | 5 | 0 | 0 | 0 | 6 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 0 |
| West | 2116 | 16 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 |
| North | 2248 | 8 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| Device | ID | Sex 1 | Deployed 2 | Activity Range (Ha) | Distance/24 h (m) 3 | Distance/Day(m) 4 | Distance/Night(m) 5 | Vector Angle | Vector Length |
|---|---|---|---|---|---|---|---|---|---|
| G–4 | Cat 4 | FE | 6 | 2.47 | 9684.03 | 5324.36 | 4850.99 | 141.14 | 0.21 |
| G–7 | Cat 7 | FE | 5 | 0.15 | 541.19 | 144.46 | 541.19 | 187.96 | 0.22 * |
| G–8 | Cat 8 | FE | 6 | 0.53 | 1547.51 | 274.44 | 1024.63 | 269.25 | 0.32 * |
| G–9 | Cat 9 | MN | 5 | 3.61 | 1740.87 | 0 | 1335.44 | 235.28 | 0.81 |
| B–5 | Cat 12 | FS | 5 | 1.31 | 2862.07 | 1847.43 | 1271.96 | 112.35 | 0.19 * |
| K–1 | Cat 15 | FE | 4 | 0.10 | 635.32 | 753.42 | 249.08 | 144.22 | 0.31 |
| K–5 | Dog 2 | FS | 5 | 1.23 | 4479.17 | 4479.17 | 444.93 | 124.30 | 0.57 |
| K–11 | Dog 3 | ME | 6 | 8.41 | 8425.57 | 5285.96 | 4602.13 | 0.19 | 0.22 |
| Dog 3 # | 32,272.64 | ||||||||
| B–14 | Dog 5 | ME | 3 | 9.35 | 2455.74 | 2205.27 | 1646.5 | 75.34 | 0.14 # |
| B–20 | Dog 6 | FS | 4 | 29.61 | 14,568 | 9146.9 | 7394.83 | 44.71 | 0.50 |
References
- Burleigh, A.; McMahon, S.; Kiely, S. Owned dog and cat populations in remote Indigenous communities in the Northern Territory: A retrospective study. Aust. Vet. J. 2015, 93, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Gaskin, S.; Bentham, R.; Cromar, N.; Fallowfield, H. The zoonotic potential of dogs in Aboriginal communities in central Australia. Environ. Health 2007, 7, 36. [Google Scholar]
- Meloni, B.P.; Thompson, R.; Hopkins, R.M.; Reynoldson, J.A.; Gracey, M. The prevalence of Giardia and other intestinal parasites in children, dogs and cats from Aboriginal communities in the Kimberley. Med. J. Aust. 1993, 158, 157–159. [Google Scholar] [PubMed]
- Thompson, R.; Meloni, B.; Hopkins, R.; Deplazes, P.; Reynoldson, J. Observations on the endo-and ectoparasites affecting dogs and cats in Aboriginal communities in the north-west of Western Australia. Aust. Vet. J. 1993, 70, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, L.; Corlette, S. Dog health program in Numbulwar, a remote Aboriginal community in east Arnhem Land. Aust. Vet. J. 2006, 84, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Smout, F.A.; Skerratt, L.F.; Butler, J.R.A.; Johnson, C.N.; Congdon, B.C.; Thompson, R.A. The hookworm Ancylostoma ceylanicum: An emerging public health risk in Australian tropical rainforests and Indigenous communities. One Health 2017, 3, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Kruuk, H.; Snell, H. Prey selection by feral dogs from a population of marine iguanas (Amblyrhynchus cristatus). J. Appl. Ecol. 1981, 197–204. [Google Scholar] [CrossRef]
- Taborsky, M. Kiwis and dog predation: Observations in Waitangi State Forest. Notornis 1988, 35, 197–202. [Google Scholar]
- Borroto-Páez, R. Invasive mammals in Cuba: An overview. Biol. Invasions 2009, 11, 2279. [Google Scholar] [CrossRef]
- Banks, P.B.; Bryant, J.V. Four-legged friend or foe? Dog walking displaces native birds from natural areas. Biol. Lett. 2007, 3, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Glover, H.K.; Weston, M.A.; Maguire, G.S.; Miller, K.K.; Christie, B.A. Towards ecologically meaningful and socially acceptable buffers: Response distances of shorebirds in Victoria, Australia, to human disturbance. Landsc. Urban Plan. 2011, 103, 326–334. [Google Scholar] [CrossRef]
- Weston, M.A.; Stankowich, T. Dogs as agents of disturbance. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press: Oxford, UK, 2013; pp. 94–113. [Google Scholar]
- Butler, J.R.A.; Toit, J. Diet of free-ranging domestic dogs (Canis familiaris) in rural Zimbabwe: Implications for wild scavengers on the periphery of wildlife reserves. Anim. Conserv. 2002, 5, 29–37. [Google Scholar] [CrossRef]
- Vanak, A.T.; Dickman, C.R.; Silva-Rodriguez, E.A.; Butler, J.R.A.; Ritchie, E.G. Top-dogs and under-dogs: Competition between dogs and sympatric carnivores. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press: Oxford, UK, 2013; pp. 69–93. [Google Scholar]
- Knobel, D.L.; Butler, J.R.A.; Lembo, T.; Critchlow, R.; Gompper, M.E. Dogs, disease, and wildlife. In Free-Ranging Dogs and Wildlife Conservation; Oxford University Press: Oxford, UK, 2014; p. 144. [Google Scholar]
- Butler, J.R.A.; Du Toit, J.; Bingham, J. Free-ranging domestic dogs (Canis familiaris) as predators and prey in rural Zimbabwe: Threats of competition and disease to large wild carnivores. Biol. Conserv. 2004, 115, 369–378. [Google Scholar] [CrossRef]
- Dickman, C.R. Overview of the Impacts of Feral Cats on Australian Native Fauna; Australian Nature Conservation Agency Canberra: Canberra, Australia, 1996. [Google Scholar]
- Woinarski, J.; Milne, D.; Wanganeen, G. Changes in mammal populations in relatively intact landscapes of Kakadu National Park, Northern Territory, Australia. Aust. Ecol. 2001, 26, 360–370. [Google Scholar] [CrossRef]
- Medina, F.M.; Bonnaud, E.; Vidal, E.; Tershy, B.R.; Zavaleta, E.S.; Josh Donlan, C.; Keitt, B.S.; Corre, M.; Horwath, S.V.; Nogales, M. A global review of the impacts of invasive cats on island endangered vertebrates. Glob. Chang. Biol. 2011, 17, 3503–3510. [Google Scholar] [CrossRef]
- Davies, H.F.; McCarthy, M.A.; Firth, R.S.; Woinarski, J.C.; Gillespie, G.R.; Andersen, A.N.; Geyle, H.M.; Nicholson, E.; Murphy, B.P. Top-down control of species distributions: Feral cats driving the regional extinction of a threatened rodent in northern Australia. Divers. Distrib. 2016, 3, 272–283. [Google Scholar] [CrossRef]
- Doherty, T.S.; Dickman, C.R.; Glen, A.S.; Newsome, T.M.; Nimmo, D.G.; Ritchie, E.G.; Vanak, A.T.; Wirsing, A.J. The global impacts of domestic dogs on threatened vertebrates. Biol. Conserv. 2017, 210, 56–59. [Google Scholar] [CrossRef]
- Brown, W.; Kennedy, B.; Allen, J.; Cutter, S.; De Santis, R. Collaboration improves dog management and welfare in Tiwi Islands. J. Vet. Behav. Clin. Appl. Res. 2014, 9, e12. [Google Scholar] [CrossRef]
- Firth, R.S.; Woinarski, J.C.; Brennan, K.G.; Hempel, C. Environmental relationships of the brush-tailed rabbit-rat, Conilurus penicillatus, and other small mammals on the Tiwi Islands, northern Australia. J. Biogeogr. 2006, 33, 1820–1837. [Google Scholar] [CrossRef]
- Australian Government Bureau of Meteorology. Climate Statistics for Australian Locations: Point Fawcett. Available online: http://www.bom.gov.au/climate/averages/tables/cw_200731.shtml (accessed on 14 November 2017).
- Australian Bureau of Statistics. 2016 Census Quickstats: Wurrumiyanga. 2017. Available online: http://www.censusdata.abs.gov.au/census_services/getproduct/census/2016/quickstat/SSC70295?opendocument (accessed on 15 November 2017).
- Australian Government Bureau of Meteorology. Point Fawcett (Bathurst Island) June 2017 Daily Weather Observations. 2017. Available online: http://www.bom.gov.au/climate/dwo/201706/html/IDCJDW8042.201706.shtml (accessed on 14 November 2017).
- Morris, G.; Conner, L.M. Assessment of accuracy, fix success rate, and use of estimated horizontal position error (EHPE) to filter inaccurate data collected by a common commercially available GPS logger. PLoS ONE 2017, 12, e0189020. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, J.; Körtner, G.; Ballard, G.; Fleming, P.J.; Brown, W.Y. Effects of sex and reproductive state on interactions between free-roaming domestic dogs. PLoS ONE 2014, 9, e116053. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Englewood Cliffs, NJ, USA, 1996. [Google Scholar]
- Körtner, G.; Holznagel, N.; Fleming, P.J.; Ballard, G. Home range and activity patterns measured with GPS collars in spotted-tailed quolls. Aust. J. Zool. 2016, 63, 424–431. [Google Scholar] [CrossRef]
- Crawley, M.J. The R Book, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Meek, P.; Ballard, G.; Claridge, A.; Kays, R.; Moseby, K.; O’brien, T.; O’Connell, A.; Sanderson, J.; Swann, D.; Tobler, M. Recommended guiding principles for reporting on camera trapping research. Biodivers. Conserv. 2014, 23, 2321–2343. [Google Scholar] [CrossRef]
- Meek, P.D.; Ballard, G.; Fleming, P.J. The pitfalls of wildlife camera trapping as a survey tool in Australia. Aust. Mammal. 2015, 37, 13–22. [Google Scholar] [CrossRef]
- Thomson, P.C. The behavioural ecology of dingoes in north-western Australia. II. Activity patterns, breeding season and pup rearing. Wildl. Res. 1992, 19, 519–529. [Google Scholar] [CrossRef]
- Meek, P.D. Home range of house cats Felis catus living within a National Park. Aust. Mammal. 2003, 25, 51–60. [Google Scholar]
- Horn, J.A.; Mateus-Pinilla, N.; Warner, R.E.; Heske, E.J. Home range, habitat use, and activity patterns of free-roaming domestic cats. J. Wildl. Manag. 2011, 75, 1177–1185. [Google Scholar] [CrossRef]
- Schmidt, P.M.; Lopez, R.R.; Collier, B.A. Survival, fecundity, and movements of free-roaming cats. J. Wildl. Manag. 2007, 71, 915–919. [Google Scholar] [CrossRef]
- Legge, S.; Murphy, B.; McGregor, H.; Woinarski, J.; Augusteyn, J.; Ballard, G.; Baseler, M.; Buckmaster, T.; Dickman, C.; Doherty, T. Enumerating a continental-scale threat: How many feral cats are in Australia? Biol. Conserv. 2017, 206, 293–303. [Google Scholar] [CrossRef]
- Burbidge, A.A.; McKenzie, N.; Brennan, K.; Woinarski, J.; Dickman, C.; Baynes, A.; Gordon, G.; Menkhorst, P.; Robinson, A. Conservation status and biogeography of Australia’s terrestrial mammals. Aust. J. Zool. 2009, 56, 411–422. [Google Scholar] [CrossRef]
- Woinarski, J.C.; Legge, S.; Fitzsimons, J.A.; Traill, B.J.; Burbidge, A.A.; Fisher, A.; Firth, R.S.; Gordon, I.J.; Griffiths, A.D.; Johnson, C.N. The disappearing mammal fauna of northern Australia: Context, cause, and response. Conserv. Lett. 2011, 4, 192–201. [Google Scholar] [CrossRef]

| Sex | Desexed | Entire | Unknown | Total |
|---|---|---|---|---|
| Dog Census | ||||
| February | ||||
| Female | 74 (71.2%) | 23 (22.1%) | 7 (6.7%) | 104 |
| Male | 36 (47.3%) | 29 (38.2%) | 11 (14.5%) | 76 |
| Unknown | 0 | 4 (36.4%) | 7 (63.6%) | 11 |
| Total | 110 (57.6%) | 56 (29.3%) | 25 (13.1%) | 191 |
| Cat Census | ||||
| February | ||||
| Female | 4 (22.2%) | 12 (66.7%) | 2 (11.1%) | 18 |
| Male | 3 (25%) | 8 (66.7%) | 1 (8.3%) | 12 |
| Unknown | 0 | 11 (100%) | 0 | 11 |
| Total | 7 (17.1%) | 31 (75.6%) | 3 (7.3%) | 41 |
| April | ||||
| Female | 14 (36.8%) | 21 (55.3%) | 3 (7.9%) | 38 |
| Male | 6 (28.6%) | 14 (66.7%) | 1 (4.8%) | 21 |
| Unknown | 0 | 12 (92.3%) | 1(7.7%) | 13 |
| Total | 20 (27.8%) | 47 (65.3%) | 5 (6.9%) | 72 |
| June | ||||
| Female | 29 (60.4%) | 19 (39.6%) | 0 | 48 |
| Male | 13 (43.3%) | 17 (56.7%) | 0 | 30 |
| Unknown | 0 | 2 (100%) | 0 | 2 |
| Total | 42 (52.5%) | 38 (47.5%) | 0 | 80 |
| August | ||||
| Female | 37 (72.5%) | 14 (27.5%) | 0 | 51 |
| Male | 19 (61.3%) | 12 (38.7%) | 0 | 31 |
| Unknown | 1 (100%) | 0 | 0 | 1 |
| Total | 57 (68.7%) | 26 (31.3%) | 0 | 83 |
| Parameter/Statistical Test | Mean ± SD | n | DF | F | p |
|---|---|---|---|---|---|
| Transect | |||||
| April | |||||
| Roaming | 0.5 ± 0.61 | 18 | |||
| Nonroaming | 4.66 ± 3.16 | 18 | |||
| ANOVA (behaviour v survey) | 1 | 30.1 | <0.001 | ||
| June | |||||
| Roaming | 0.5 ± 0.85 | 18 | |||
| Nonroaming | 2.9 ± 1.73 | 18 | |||
| ANOVA (behaviour v survey) | 1 | 28.82 | <0.001 | ||
| Activity range (AR) | |||||
| Dog | 12.15 ha ± 12.19 ha | 4 | |||
| Cat | 1.36 ha ± 1.42 ha | 6 | |||
| ANOVA (AR v Species) | 1 | 4.901 | 0.057 * | ||
| Distance Travelled per 24 h | |||||
| Dog | 1233.06 m ± 545.43 m | 20 (4) | |||
| Cat | 703.14 m ± 348.12 m | 29 (6) | |||
| M-EM # (Distance v Species) | 1, 8 | 5.11 | 0.054 * | ||
| Distance Travelled per day | |||||
| Dog | 949.68 m ± 342.82 m | 20 (4) | |||
| Cat | 455.42 m ± 238.11 m | 29 (6) | |||
| M-EM # (Distance v Species) | 1, 7 | 7.67 | 0.028 | ||
| Distance Travelled per night | |||||
| Dog | 790.5 m ± 255.86 m | 20 (4) | |||
| Cat | 585.1 m ± 161.24 m | 29 (6) | |||
| M-EM # (Distance v Species) | 1, 8 | 9.55 | 0.015 | ||
| Parameter | Vector Time ± SD | n | r | R | z | p |
|---|---|---|---|---|---|---|
| Transects (pooled) | ||||||
| Roaming | - | 18 | 0.25 | 4.58 | 1.17 | 0.32 * |
| Non-roaming | 12:31 a.m. ± 4 h 34 min | 130 | 0.28 | 36.72 | 10.37 | <0.001 |
| Camera | ||||||
| Dogs a | 8:06 a.m. and 8:06 p.m. ± 3 h 52 min | 50 | 0.62 | 31.46 | 19.80 | <0.001 |
| Cats b | 3 | |||||
| Wildlife | 11:50 p.m. ± 3 h 52 min | 0.48 | 49.14 | 23.67 | <0.001 | |
| Wildlife (flightless) | 12:06 a.m. ± 3 h 9 min | 0.65 | 59.21 | 38.96 | <0.001 | |
| GPS Loggers | ||||||
| Dogs | 6:02 a.m. ± 5 h 6 min | 4 | 0.10 | 68.71 | 6.99 | <0.001 |
| Cats | 11:24 a.m. ± 5 h 10 min | 6 | 0.08 | 58.29 | 4.80 | 0.008 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kennedy, B.; Brown, W.Y.; Vernes, K.; Körtner, G.; Butler, J.R.A. Dog and Cat Interactions in a Remote Aboriginal Community. Animals 2018, 8, 65. https://doi.org/10.3390/ani8050065
Kennedy B, Brown WY, Vernes K, Körtner G, Butler JRA. Dog and Cat Interactions in a Remote Aboriginal Community. Animals. 2018; 8(5):65. https://doi.org/10.3390/ani8050065
Chicago/Turabian StyleKennedy, Brooke, Wendy Y. Brown, Karl Vernes, Gerhard Körtner, and James R. A. Butler. 2018. "Dog and Cat Interactions in a Remote Aboriginal Community" Animals 8, no. 5: 65. https://doi.org/10.3390/ani8050065
APA StyleKennedy, B., Brown, W. Y., Vernes, K., Körtner, G., & Butler, J. R. A. (2018). Dog and Cat Interactions in a Remote Aboriginal Community. Animals, 8(5), 65. https://doi.org/10.3390/ani8050065