Use of Unpalatable Forages by Ruminants: The Influence of Experience with the Biophysical and Social Environment

{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. The Importance of Experience in Ruminant Animals
3. Positive Experience and Use of Low-Quality Feed
4. Positive Experience and Use of Toxin-Containing Feed
5. Experience and the Gut Microbiome
6. Use of Toxin-Containing Feed to Combat Disease
7. Negative Consequences of Being Exposed to an Unpalatable Feed
8. “Directions” of the Feeding Responses as a Function of Experience
9. Evolutionary Consequences of Incorporating Unpalatable Plants in the Diet
10. Plant Community Consequences of Incorporating Unpalatable Plants in the Diet
11. Behaviour-Based Management
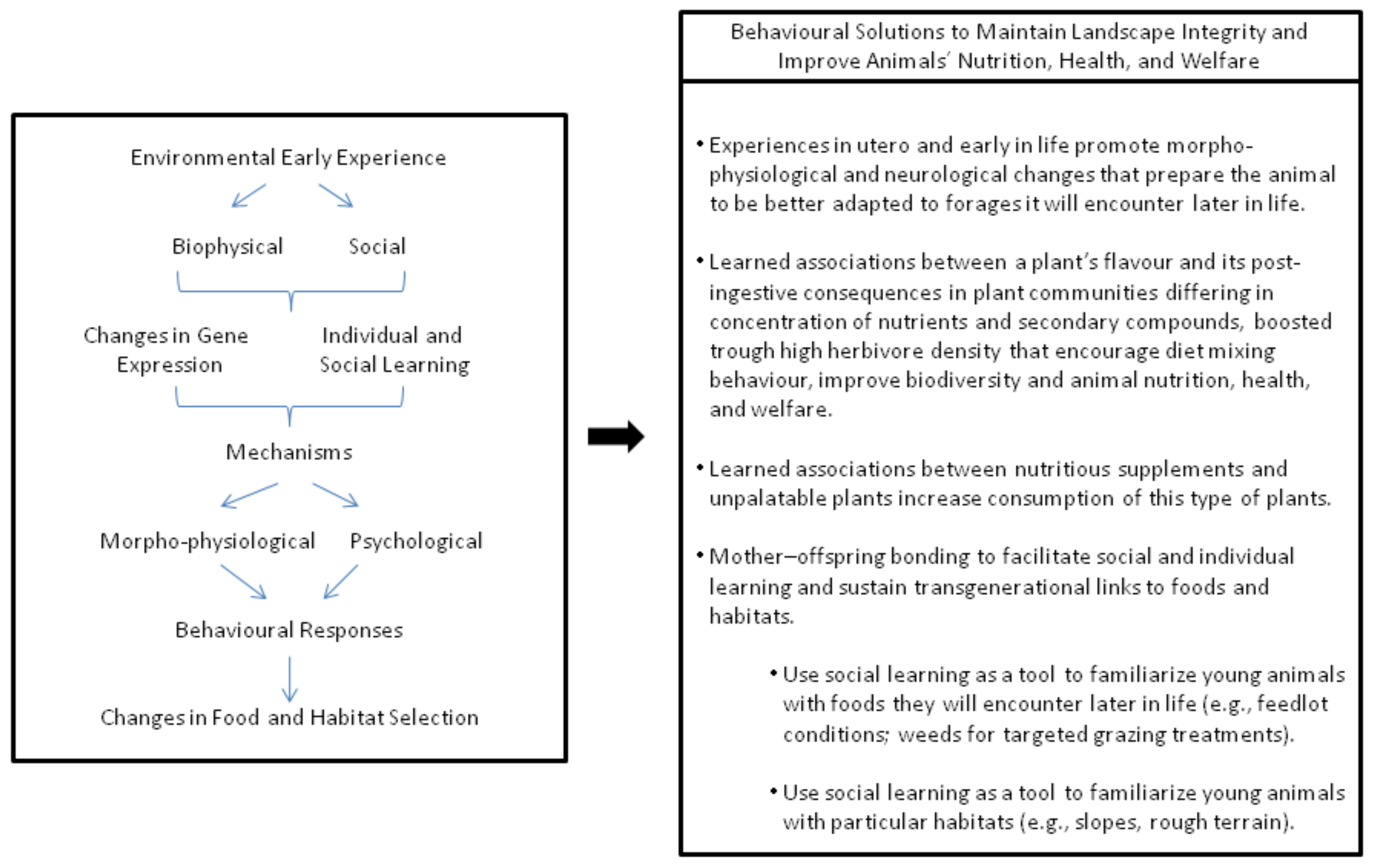
12. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Freeland, W.J.; Janzen, D.H. Strategies in herbivory by mammals: The role of plant secondary compounds. Am. Nat. 1974, 108, 269–289. [Google Scholar] [CrossRef]
- Hofmann, R.R. Anatomy of the gastrointestinal tract. In The Ruminant Animal; Church, D.C., Ed.; Prentice Hall: Englewood Cliffs, NJ, USA, 1988; pp. 14–43. [Google Scholar]
- Distel, R.A.; Provenza, F.D. Experience early in life affects voluntary intake of blackbrush by goats. J. Chem. Ecol. 1991, 17, 431–450. [Google Scholar] [CrossRef] [PubMed]
- Distel, R.A.; Villalba, J.J.; Laborde, H.E. Effects of early experience on voluntary intake of low-quality roughage by sheep. J. Anim. Sci. 1994, 72, 1191–1195. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Distel, R.A.; Villalba, J.J.; Laborde, H.E.; Burgos, M.A. Persistence of the effects of early experience on consumption of low-quality roughage by sheep. J. Anim. Sci. 1996, 74, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Freidin, E.; Catanese, F.; Didoné, N.; Distel, R.A. Mechanisms of intake induction of a low-nutritious food in sheep (Ovis aries). Behav. Processes 2011, 87, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Freidin, E.; Catanese, F.; Cuello, M.I.; Distel, R.A. Induction of low-nutritious food consumption by subsequent nutrient supplementation in sheep (Ovis aries). Animal 2012, 6, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Provenza, F.D.; Balph, D.F. Applicability of five diet-selection models to various foraging challenges ruminants encounter. In Behavioural Mechanisms of Food Selection; Hughes, R.N., Ed.; Springer: Berlin, Germany, 1990; pp. 423–459. [Google Scholar]
- Provenza, F.D.; Lynch, J.J.; Nolan, J.V. Food aversion conditioned in anesthetized sheep. Physiol. Behav. 1994, 55, 429–432. [Google Scholar] [CrossRef]
- Provenza, F.D. Postingestive feedback as an elementary determinant of food preference and intake in ruminants. J. Range Manag. 1995, 48, 2–17. [Google Scholar] [CrossRef]
- Provenza, F.D.; Villalba, J.J. Foraging in domestic herbivores: Linking the internal and external milieux. In Feeding in Domestic Vertebrates: From Structure to Behaviour; Bels, V.L., Ed.; CABI Publishers: Oxfordshire, UK, 2006; pp. 201–240. [Google Scholar]
- Bandura, A. Social Learning Theory; Prentice-Hall: Englewood Cliffs, NJ, USA, 1977. [Google Scholar]
- Nolte, D.L.; Provenza, F.D. Foods preferences in lambs after exposure to flavours in milk. Appl. Anim. Behav. Sci. 1992, 32, 381–389. [Google Scholar] [CrossRef]
- Nolte, D.L.; Provenza, F.D.; Callan, R.; Panter, K.E. Garlic in the ovine fetal environment. Physiol. Behav. 1992, 52, 1091–1093. [Google Scholar] [CrossRef]
- Galef, B.G., Jr.; Laland, K.N. Social learning in animals: Empirical studies and theoretical models. Bioscience 2005, 55, 489–499. [Google Scholar] [CrossRef]
- Saint-Dizier, H.; Levy, F.; Ferreira, G. Influence of the mother in the development of flavored-food preference in lambs. Dev. Psychobiol. 2007, 49, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Mirza, S.N.; Provenza, F.D. Preference of the mother affects selection and avoidance of foods by lambs differing in age. Appl. Anim. Behav. Sci. 1990, 28, 255–263. [Google Scholar] [CrossRef]
- Glasser, T.A.; Ungar, E.D.; Landau, S.Y.; Perevolotsky, A.; Muklada, H.; Walker, J.W. Breed and maternal effects on the intake of tannin-rich browse by juvenile domestic goats (Capra hircus). Appl. Anim. Behav. Sci. 2009, 119, 71–77. [Google Scholar] [CrossRef]
- Heyes, C.M.; Galef, B.G., Jr. Social Learning and Imitation: The Roots of Culture; Academic Press: New York, NY, USA, 1996. [Google Scholar]
- Baraza, E.; Villalba, J.J.; Provenza, F.D. Nutritional context influences preferences of lambs for foods with plant secondary metabolites. Appl. Anim. Behav. Sci. 2005, 92, 293–305. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Shaw, R. Initial conditions and temporal delays influence preference for foods high in tannins and for foraging locations with and without foods high in tannins by sheep. Appl. Anim. Behav. Sci. 2006, 97, 190–205. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Catanese, F.; Distel, R.A. Understanding and manipulating diet choice in grazing animals. Anim. Prod. Sci. 2015, 55, 261–271. [Google Scholar] [CrossRef]
- Catanese, F.; Fernández, P.; Villalba, J.J.; Distel, R.A. The physiological consequences of ingesting a toxic plant (Diplotaxis tenuifolia) influence subsequent foraging decisions by sheep (Ovis aries). Physiol. Behav. 2016, 167, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Coppersmith, R.; Leon, M. Enhanced neural response to familiar olfactory cues. Science 1984, 225, 849–852. [Google Scholar] [CrossRef]
- LeDoux, J. Synaptic Self: How Our Brains Become Who We Are; Viking Penguin: New York, NY, USA, 2002. [Google Scholar]
- Doidge, N. The Brain that Changes Itself: Stories of Personal Triumph from the Frontiers of Brain Science; Penguin Books: New York, NY, USA, 2007. [Google Scholar]
- Schlichting, C.D.; Pigliucci, M. Phenotypic Evolution: A Reaction Norm Perspective; Sinauer Publications: Sunderland, MA, USA, 1998. [Google Scholar]
- Dufty, A.M., Jr.; Clobert, J.; Moller, A.P. Hormones, developmental plasticity and adaptation. Trends Ecol. Evol. 2002, 17, 190–196. [Google Scholar] [CrossRef]
- Provenza, F.D. What does it mean to be locally adapted and who cares anyway? J. Anim. Sci. 2008, 86, E271–E284. [Google Scholar] [CrossRef] [PubMed]
- DiPietro, J.A.; Ghera, M.M.; Costigan, K.; Hawkins, M. Measuring the ups and downs of pregnancy stress. J. Psychosom. Obstet. Gyn. 2004, 25, 189–201. [Google Scholar] [CrossRef]
- Paul, A.M. Origins: How the Nine Months Before Birth Shape the Rest of Our Lives; Free Press: New York, NY, USA, 2010. [Google Scholar]
- Barker, D.J.; Osmond, C. Infant mortality, childhood nutrition, and ischaemic heart disease in England and Wales. Lancet 1986, 327, 1077–1081. [Google Scholar] [CrossRef]
- Barker, D.J.; Osmond, C.; Simmonds, S.J.; Wield, G.A. The relation of small head circumference and thinness at birth to death from cardiovascular disease in adult life. Br. Med. J. 1993, 306, 422–426. [Google Scholar] [CrossRef]
- Fish, E.W.; Shahrokh, D.; Bagot, R.; Caldji, C.; Bredy, T.; Szyf, M.; Meaney, M.J. Epigenetic programming of stress responses through variations in maternal care. Ann. N. Y. Acad. Sci. 2004, 1036, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Meaney, M.J.; Szyf, M. Maternal care as a model for experience-dependent chromatin plasticity? Trends Neurosci. 2005, 28, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Anacker, C.; O’Donnell, K.J.; Meaney, M.J. Early life adversity and the epigenetic programming of hypothalamic-pituitary-adrenal function. Dialogues Clin. Neurosci. 2014, 16, 321–333. [Google Scholar] [PubMed]
- Turecki, G.; Meaney, M.J. Effects of the social environment and stress on glucocorticoid receptor gene methylation: A systematic review. Biol. Psychiatry 2016, 79, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Funston, R.N.; Summers, A.F. Epigenetics: Setting up lifetime production of beef cows by managing nutrition. Annu. Rev. Anim. Biosci. 2013, 1, 339–363. [Google Scholar] [CrossRef] [PubMed]
- Provenza, F.D. Tracking variable environments: There is more than one kind of memory. J. Chem. Ecol. 1995, 21, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Green, G.C.; Elwin, R.L.; Mottershead, B.E.; Lynch, J.J. Long-term effects of early experience to supplementary feeding in sheep. Proc. Aust. Soc. Anim. Prod. 1984, 15, 373–375. [Google Scholar]
- Biquand, S.; Biquand-Guyot, V. The influence of peers, lineage and environment on food selection of the criollo goat. Appl. Anim. Behav. Sci. 1992, 43, 231–245. [Google Scholar] [CrossRef]
- Wiedmeier, R.D.; Provenza, F.D.; Burritt, E.A. Exposure to ammoniated wheat straw as suckling calves improves performance of mature beef cows wintered on ammoniated wheat straw. J. Anim. Sci. 2002, 80, 2340–2348. [Google Scholar] [CrossRef] [PubMed]
- Provenza, F.D. Twenty-five years of paradox in plant-herbivore interactions and “sustainable” grazing management. Rangelands 2003, 25, 4–15. [Google Scholar] [CrossRef]
- Davis, J.M.; Stamps, J.A. The effect of natal experience on habitat preferences. Trends Ecol. Evol. 2004, 19, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Laiolo, P.; Tella, J.L. Erosion of animal cultures in fragmented landscapes. Front. Ecol. Environ. 2007, 5, 68–72. [Google Scholar] [CrossRef]
- Chadwick, M.A.; Vercoe, P.E.; Williams, I.H.; Revell, D.K. Programming sheep production on saltbush: Adaptations of offspring from ewes that consumed high amounts of salt during pregnancy and early lactation. Anim. Prod. Sci. 2009, 49, 311–317. [Google Scholar] [CrossRef]
- Peñagaricano, F.; Souza, A.H.; Carvahlo, P.D.; Driver, A.M.; Gambra, R.; Kropp, J.; Hackbart, K.S.; Luchini, D.; Shaver, R.D.; Wiltbank, M.C.; et al. Effect of maternal methionine supplementation on the transcriptome of bovine preimplantation embryos. PLoS ONE 2013, 8, e72302. [Google Scholar] [CrossRef] [PubMed]
- Villalba, J.J.; Provenza, F.D.; Manteca, X. Links between ruminants’ food preference and their welfare. Animal 2010, 4, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Catanese, F.; Obelar, M.; Villalba, J.J.; Distel, R.A. The importance of diet choice on stress-related responses by lambs. Appl. Anim. Behav. Sci. 2013, 148, 37–45. [Google Scholar] [CrossRef]
- Provenza, F.D.; Pfister, J.A.; Cheney, C.D. Mechanisms of learning in diet selection with reference to phytotoxicosis in herbivores. J. Range Manag. 1992, 45, 36–45. [Google Scholar] [CrossRef]
- Provenza, F.D.; Balph, D.F. Development of dietary choice in livestock on rangelands and its implications for management. J. Anim. Sci. 1988, 66, 2356–2368. [Google Scholar] [CrossRef]
- Bossdorf, O.; Richards, C.L.; Pigliucci, M. Epigenetics for ecologists. Ecol. Lett. 2008, 11, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.D.; Provenza, F.D.; Pfister, J.A. Do plant secondary compounds induce epigenetic changes that confer resistance or susceptibility to toxicosis in animals? In Animal Farming and Environmental Interactions in the Mediterranean Region; Casasús, I., Rogošiç, J., Rosati, A., Štokoviç, I., Gabiña, D., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2012; pp. 33–44. [Google Scholar]
- Wiedmeier, R.W.; Villalba, J.J.; Summers, A.; Provenza, F.D. Eating a high fiber diet during pregnancy increases intake and digestibility of a high fiber diet by offspring in cattle. Anim. Feed Sci. Technol. 2012, 177, 144–151. [Google Scholar] [CrossRef]
- Nolte, D.L.; Provenza, F.D.; Balph, D.F. The establishment and persistence of food preferences in lambs exposed to selected foods. J. Anim. Sci. 1990, 68, 998–1002. [Google Scholar] [CrossRef] [PubMed]
- Montes-Sánchez, J.J.; Villalba, J.J. Effects of early experience and alternative feeds on medusahead (Taeniatherum caput-medusae ssp. asperum) intake by sheep. Appl. Anim. Behav. Sci. 2017, 188, 9–16. [Google Scholar] [CrossRef]
- Montes-Sánchez, J.J.; Van Miegroet, H.; Villalba, J.J. Effects energy supplementation and time on use of medusahead by grazing ewes and their lambs. Rangel. Ecol. Manag. 2017, 70, 380–387. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D. Preference for wheat straw by lambs conditioned with intraruminal infusions of starch. Br. J. Nutr. 1997, 77, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Villalba, J.J.; Provenza, F.D. Preference for flavoured foods by lambs conditioned with intraruminal administration of nitrogen. Br. J. Nutr. 1997, 78, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Catanese, F.; Distel, R.A.; Villalba, J.J. Expression of conditioned preference for low-quality food in sheep is modulated by foraging costs. Animal 2015, 9, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Catanese, F.; Distel, R.A.; Fernández, P.; Villalba, J.J. How the foraging decisions of a small ruminant are influenced by past feeding experiences with low-quality food. Behav. Processes 2016, 126, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, M.A.; Williams, I.H.; Vercoe, P.E.; Revell, D.K. Feeding pregnant ewes a high-salt diet or saltbush suppresses their offspring’s postnatal renin activity. Animal 2009, 3, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.A.; Villalba, J.J.; Provenza, F.D. Influence of stock density and rate and temporal patterns of forage allocation on the diet mixing behavior of sheep grazing sagebrush steppe. Appl. Anim. Behav. Sci. 2006, 100, 207–218. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Banner, R.E. Influence of macronutrients and activated charcoal on intake of sagebrush by sheep and goats. J. Anim. Sci. 2002, 80, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.A.; Villalba, J.J.; Provenza, F.D. Influence of experience on browsing sagebrush by cattle and its impacts on plant community structure. Rangel. Ecol. Manag. 2014, 67, 78–87. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Mishra, A.S. Glucosinolates in animal nutrition: A review. Anim. Feed Sci. Technol. 2007, 132, 1–27. [Google Scholar] [CrossRef]
- Kohl, K.D.; Dearing, M.D. Experience matters: Prior exposure to plant toxins enhances diversity of gut microbes in herbivores. Ecol. Lett. 2012, 15, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Kohl, K.D.; Weiss, R.B.; Cox, J.; Dale, C.; Dearing, M.D. Gut microbes of mammalian herbivores facilitate intake of plant toxins. Ecol. Lett. 2014, 17, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Jasmonate and salicylate induce expression of herbivore cytochrome P450 genes. Nature 2002, 419, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Delgoda, R.; Westlake, A.C.G. Herbal interactions involving cytochrome P450 enzymes. Toxicol. Rev. 2004, 23, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Villalba, J.J.; Provenza, F.D.; Han, G.D. Experience influences diet mixing by herbivores: Implications for plant biochemical diversity. Oikos 2004, 107, 100–109. [Google Scholar] [CrossRef]
- Dziba, L.E.; Hall, J.O.; Provenza, F.D. Feeding behavior of the lambs in relation to kinetics of 1,8-cineole dosed intravenously or into the rumen. J. Chem. Ecol. 2006, 32, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Cortese, R.; Lu, L.; Yu, Y.; Ruden, D.; Claud, E.C. Epigenome-microbiome crosstalk: A potential new paradigm influencing neonatal susceptibility to disease. Epigenetics 2016, 11, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Wu, W.; Li, Y.; Li, L. Influences of the gut microbiota on DNA methylation and histone modification. Dig. Dis. Sci. 2017, 62, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Sugi, Y.; Nakano, K.; Tsuda, M.; Kurihara, K.; Hosono, A.; Kaminogawa, S. Epigenetic control of the host gene by commensal bacteria in large intestinal epithelial cells. J. Biol. Chem. 2011, 286, 35755–35762. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Prince, A.L.; Bader, D.; Hu, M.; Ganu, R.; Baquero, K.; Blundell, P.; Harris, R.A.; Frias, A.E.; Grove, K.L.; et al. High-fat maternal diet during pregnancy persistently alters the offspring microbiome in a primate model. Nat. Commun. 2014, 5, 3889. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.S.; Shriker, E.; Gold, B.; Durman, T.; Zarecki, R.; Ruppin, E.; Mizrahi, I. Diet-induced changes of redox potential underlie compositional shifts in the rumen archaeal community. Environ. Microbiol. 2016, 19, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and feeding pattern affect the diurnal dynamics of the gut microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Romano, M. Gut microbiota as a trigger of accelerated directional adaptive evolution: Acquisition of herbivory in the context of extracellular vesicles, microRNAs and inter-kingdom crosstalk. Front. Microbiol. 2017, 8, 721. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.R.; Athanasiadou, S.; Kyriazakis, I.; Gordon, I.J. Can animals use foraging behaviour to combat parasites? Proc. Nutr. Soc. 2003, 62, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Min, B.R.; Hart, S.P. Tannins for suppression of internal parasites. J. Anim. Sci. 2003, 81, E102–E109. [Google Scholar]
- Provenza, F.D.; Villalba, J.J. The role of natural plant products in modulating the immune system: An adaptable approach for combating disease in grazing animals. Small Rumin. Res. 2010, 89, 131–139. [Google Scholar] [CrossRef]
- Lozano, G.A. Parasitic stress and self-medication in wild animals. In Advances in the Study of Behavior; Moler, A.P., Milinski, M., Slater, P.J.B., Eds.; Elsevier Science: London, UK, 1998; pp. 291–317. [Google Scholar]
- Niezen, J.H.; Robertson, H.A.; Waghorn, G.C.; Charleston, W.A. Production, faecal egg counts and worm burdens of ewe lambs which grazed six contrasting forages. Vet. Parasitol. 1998, 80, 15–27. [Google Scholar] [CrossRef]
- Butter, N.L.; Dawson, J.M.; Wakelin, D.; Buttery, P.J. Effect of dietary tannin and protein concentration on nematode infection (Trichostrongylus colubriformis) in lambs. J. Agric. Sci. 2000, 134, 89–99. [Google Scholar] [CrossRef]
- Paolini, V.; Fouraste, I.; Hoste, H. In vitro effects of three woody plant and sainfoin extracts on 3rd-stage larvae and adult worms of three gastrointestinal nematodes. Parasitology 2004, 129, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Athanasiadou, S.; Kyriazakis, I.; Jackson, F.; Coop, R.L. Effects of short-term exposure to condensed tannins on adult Trichostrongylus colubriformis. Vet. Rec. 2000, 146, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Athanasiadou, S.; Kyriazakis, I.; Jackson, F.; Coop, R.L. Direct anthelmintic effects of condensed tannins towards different gastrointestinal species: In vitro and in vivo studies. Vet. Parasitol. 2001, 99, 205–219. [Google Scholar] [CrossRef]
- Athanasiadou, S.; Kyriazakis, I. Plant secondary metabolites: Antiparasitic effects and their role in ruminant production systems. Proc. Nutr. Soc. 2004, 63, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Coop, R.L.; Kyriazakis, I. Influence of host nutrition on the development and consequences of nematode parasitism in ruminants. Trends Parasitol. 2001, 17, 325–350. [Google Scholar] [CrossRef]
- Reed, J.D. Nutritional toxicology of tannins and related polyphenols in forage legumes. J. Anim. Sci. 1995, 73, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Foley, W.J.; Iason, G.R.; McArthur, C. Role of plant secondary metabolites in the nutritional ecology of mammalian herbivores: How far have we come in 25 years? In Nutritional Ecology of Herbivores; Fahey, G.C., Jung, H.J.G., Eds.; American Society of Animal Science: Champaign, IL, USA, 1999; pp. 103–209. [Google Scholar]
- Lisonbee, L.D.; Villalba, J.J.; Provenza, F.D.; Hall, J.O. Tannins and self-medication: Implications for sustainable parasite control in herbivores. Behav. Processes 2009, 82, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Villalba, J.J.; Provenza, F.D.; Hall, J.O.; Lisonbee, L.D. Selection of tannins by sheep in response to gastrointestinal nematode infection. J. Anim. Sci. 2010, 88, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Juhnke, J.; Miller, J.; Hall, J.O.; Provenza, F.D.; Villalba, J.J. Preference for condensed tannins by sheep in response to challenge infection with Haemonchus contortus. Vet. Parasitol. 2012, 188, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Sanga, U.; Provenza, F.D.; Villalba, J.J. Transmission of self-medicative behaviour from mother to offspring in sheep. Anim. Behav. 2011, 82, 219–227. [Google Scholar] [CrossRef]
- Catanese, F.; Distel, R.A.; Rodríguez Iglesias, R.M.; Villalba, J.J. Role of early experience in the development of preference for low-quality food in sheep. Animal 2010, 4, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Catanese, F.; Freidin, E.; Cuello, M.I.; Distel, R.A. Devaluation of low-quality food during early experience by sheep. Animal 2011, 5, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Provenza, F.D. Acquired aversions as the basis for varied diets of ruminants foraging on rangelands. J. Anim. Sci. 1996, 74, 2010–2020. [Google Scholar] [CrossRef] [PubMed]
- Provenza, F.D.; Burritt, E.A.; Clausen, T.P.; Bryant, J.P.; Reichardt, P.B.; Distel, R.A. Conditioned flavor aversion: A mechanism for goats to avoid condensed tannins in blackbrush. Am. Nat. 1990, 136, 810–828. [Google Scholar] [CrossRef]
- Provenza, F.D.; Lynch, J.J.; Burritt, E.A.; Scott, C.B. How goats learn to distinguish between novel foods that differ in postingestive consequences. J. Chem. Ecol. 1994, 20, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Provenza, F.D.; Malechek, J.C. Diet selection by domestic goats in relation to blackbrush twig chemistry. J. Appl. Ecol. 1984, 21, 831–841. [Google Scholar] [CrossRef]
- Provenza, F.D.; Malechek, J.C. A comparison of food selection and foraging behavior in juvenile and adult goats. Appl. Anim. Behav. Sci. 1986, 16, 49–61. [Google Scholar] [CrossRef]
- Johnson, I.T.; Gee, J.M.; Price, K.; Curl, C.; Fenwick, G.R. Influence of saponins on gut permeability and active nutrient transport in vitro. J. Nutr. 1986, 116, 2270–2277. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Rivera, L.R.; Cho, H.J.; Bravo, D.M.; Callaghan, B. The gut as a sensory organ. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Longmore, A.T. Does Experience with Sagebrush in Utero and Early in Life Influence the Use of Sagebrush by Sheep? Master’s Thesis, Utah State University, Logan, UT, USA, 2016. [Google Scholar]
- Stephens, D.W.; Brown, J.S.; Ydenberg, R.C. Foraging: Behavior and Ecology; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Bailey, D.W.; Gross, J.E.; Laca, E.A.; Rittenhouse, L.R.; Coughenour, M.B.; Swift, D.M.; Sims, P.L. Mechanisms that result in large herbivore grazing distribution patterns. J. Range Manag. 1996, 49, 386–400. [Google Scholar] [CrossRef]
- Focardi, S.; Marcellini, P.; Montanaro, P. Do ungulates exhibit a food density threshold? A field study of optimal foraging and movement patterns. J. Anim. Ecol. 1996, 65, 606–620. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Briske, D.D. Developmental morphology and physiology of grasses. In Grazing Management: An Ecological Perspective; Heitschmidt, R.K., Stuth, J.W., Eds.; Timber Press: Portland, OR, USA, 1991; pp. 85–108. [Google Scholar]
- Archer, S.; Smeins, F.E. Ecosystem-level processes. In Grazing Management: An Ecological Perspective; Heitschmidt, R.K., Stuth, J.W., Eds.; Timber Press: Portland, OR, USA, 1991; pp. 109–139. [Google Scholar]
- Moretto, A.S.; Distel, R.A. Requirement of vegetation gaps for seedling establishment of two unpalatable grasses in a native grassland of central Argentina. Austral Ecol. 1998, 23, 419–423. [Google Scholar] [CrossRef]
- Moretto, A.S.; Distel, R.A. Effects of selective defoliation on the competitive interaction between palatable and unpalatable grasses native to a temperate semi-arid grassland of Argentina. J. Arid Environ. 1999, 42, 167–175. [Google Scholar] [CrossRef]
- De Villalobos, A.E.; Peláez, D.V.; Elia, O.R. Factors related to establishment of Prosopis caldenia Burk. seedlings in central rangelands of Argentina. Acta Oecol. 2005, 27, 99–106. [Google Scholar] [CrossRef]
- De Villalobos, A.E.; Peláez, D.V.; Elia, O.R. Growth of Prosopis caldenia Burk. seedlings in central semi-arid rangelands of Argentina. J. Arid Environ. 2005, 61, 345–356. [Google Scholar] [CrossRef]
- Provenza, F.D.; Villalba, J.J.; Dziba, L.E.; Atwood, S.B.; Banner, R.E. Linking herbivore experience, varied diets, and plant biochemical diversity. Small Rumin. Res. 2003, 49, 257–274. [Google Scholar] [CrossRef]
- Dziba, L.E.; Provenza, F.D.; Villalba, J.J.; Atwood, S.B. Supplemental energy and protein increase use of sagebrush by sheep. Small Rumin. Res. 2007, 69, 203–207. [Google Scholar] [CrossRef]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Distel, R.A.; Villalba, J.J. Use of Unpalatable Forages by Ruminants: The Influence of Experience with the Biophysical and Social Environment. Animals 2018, 8, 56. https://doi.org/10.3390/ani8040056
Distel RA, Villalba JJ. Use of Unpalatable Forages by Ruminants: The Influence of Experience with the Biophysical and Social Environment. Animals. 2018; 8(4):56. https://doi.org/10.3390/ani8040056
Chicago/Turabian StyleDistel, Roberto A., and Juan J. Villalba. 2018. "Use of Unpalatable Forages by Ruminants: The Influence of Experience with the Biophysical and Social Environment" Animals 8, no. 4: 56. https://doi.org/10.3390/ani8040056
APA StyleDistel, R. A., & Villalba, J. J. (2018). Use of Unpalatable Forages by Ruminants: The Influence of Experience with the Biophysical and Social Environment. Animals, 8(4), 56. https://doi.org/10.3390/ani8040056