
Orca Behavior and Subsequent Aggression Associated with Oceanarium Confinement



Abstract
:Simple Summary
Abstract
1. Introduction
2. Orca and Human Phylogeny, and an Introduction to the Observed Orcas
3. Anthropomorphism as a Tool for Cognitive Research
4. The Human-Animal Bond
5. Observations of Orca Behavior and the Petting Pool Environment
5.1. Orca Self-Control and Reaction to Human Fear
5.2. Orca Tests of Human Trust
5.3. Limited Use of Tactical Deception
5.4. Orca Pranks
5.5. Orca Greeting Rituals
5.6. Empathy
- Cognitive empathy has components of:
- ○
- Cognitive ToM which relates to taking the perspective of another.
- ○
- Affective ToM which relates to comprehending feelings in another.
- Affective empathy involves internalizing and mirroring the feelings of another.
5.7. Adaptation of Natural Behavior to Orca/Human Interactions
5.8. Cognitive Complexity
6. Pod Level Communications
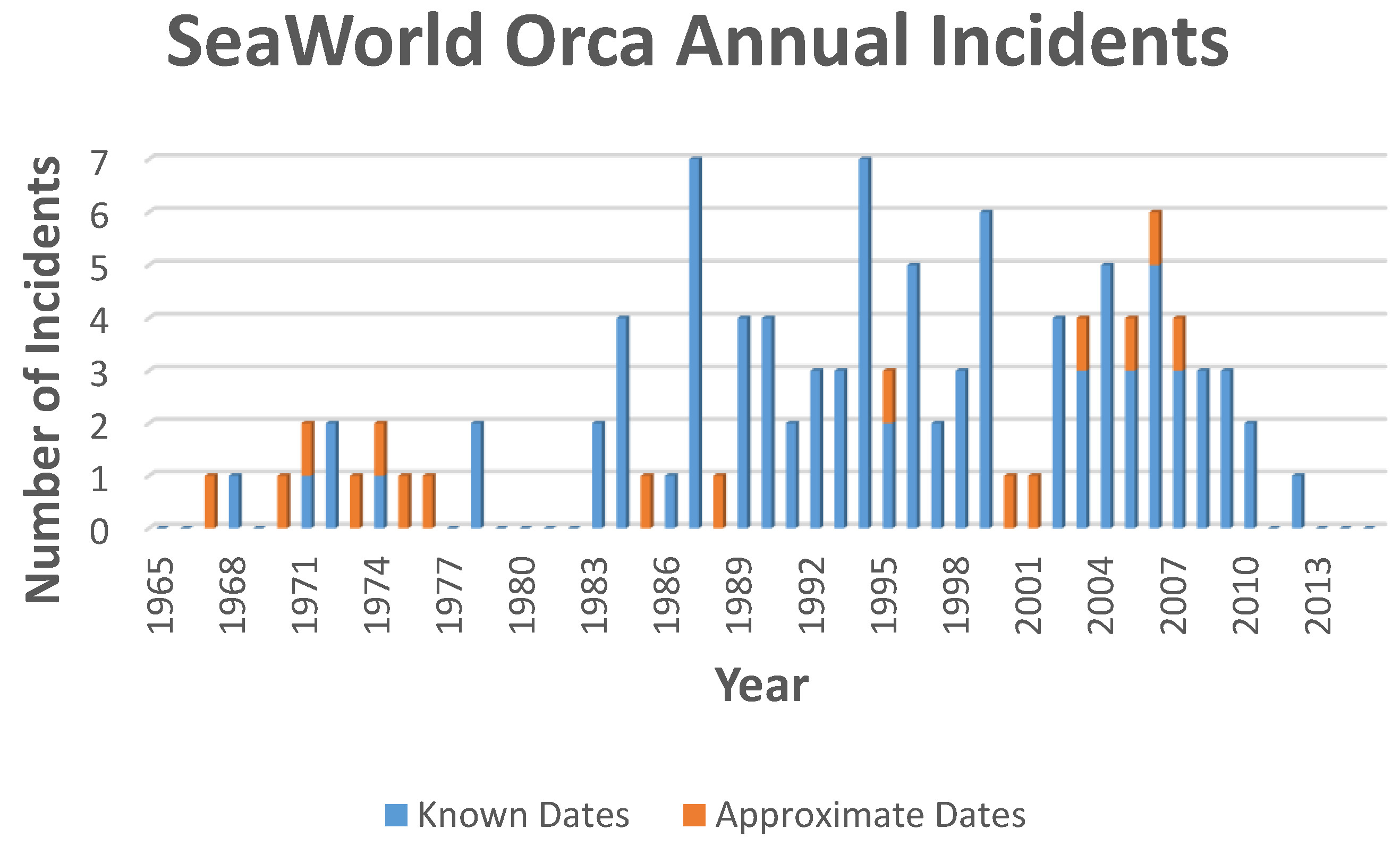
7. Subsequent Orca Aggression
- Pushing trainers about in the water
- Mouthing, sometimes to the extent of leaving permanent bite marks, and even breaking bones
- Lunging at staff who approached the water
- Pulling trainers into the water and holding them under
7.1. Post Petting Pool History
7.2. Changes To Deceptive Behavior
7.3. Possible Causation of the Behavioral Changes
8. Discussion
8.1. Common Theory of Mind Scenarios and Delphinids
8.2. Thoughts on Future Study of Orca Cognition
9. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DOAJ | Directory of open access journals |
| EQ | Encephalization Quotient |
| F1 | First generation descendant |
| F2 | Second generation descendant |
| MDPI | Multidisciplinary Digital Publishing Institute |
| RN | Royal Navy |
| ToM | Theory of Mind |
| UK | United Kingdom |
| USA | United States of America |
Appendix A

{kind=link}
{kind=link}
| Encephalization Quotient | Mammal |
|---|---|
| 7.4 to 7.8 | Human |
| 5.3 | Bottlenose Dolphin |
| 2.57 to 2.9 | Orca |
| 2.2 to 2.5 | Chimpanzee |
| 1.3 | Elephant |
| 0.9 | Horse |
References
- Gottfredson, L.S. Mainstream science on intelligence: An editorial with 52 signatories, history, and bibliography. Intelligence 1997, 24, 13–23. [Google Scholar] [CrossRef]
- Whiten, A.; van Schaik, C.P. The evolution of animal “cultures” and social intelligence. Philos. Trans. R. Soc. B 2007. [Google Scholar] [CrossRef] [PubMed]
- Brown, C. Fish intelligence, sentience and ethics. Anim. Cogn. 2015, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, J.K.; Hubrecht, R. Animal consciousness, cognition and welfare. Anim. Welf. 2001, 10, S5–S17. [Google Scholar]
- Beran, M.J.; Parrish, A.E.; Perdue, B.M.; Washburn, D.A. Comparative cognition: Past, present, and future. Int. J. Comp. Psychol. 2014, 27, 3–30. [Google Scholar] [PubMed]
- Urquiza-Haas, E.G.; Kotrschal, K. The mind behind anthropomorphic thinking: Attribution of mental states to other species. Anim. Behav. 2015, 109, 167–176. [Google Scholar] [CrossRef]
- Mann, J.; Patterson, E.M. Tool use by aquatic animals. Philos. Trans. R. Soc. B 2013, 368. [Google Scholar] [CrossRef] [PubMed]
- Marino, L. Convergence of complex cognitive abilities in cetaceans and primates. Brain Behav. Evol. 2002, 59, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Morin, P.A.; Archer, F.I.; Foote, A.D.; Vilstrup, J.; Allen, E.E.; Wade, P.; Durban, J.; Parsons, K.; Pitman, R.; Li, L.; et al. Complete mitochondrial genome phylogeographic analysis of killer whales (Orcinus orca) indicates multiple species. Genome Res. 2010, 20, 908–916. [Google Scholar] [CrossRef] [PubMed]
- McDougall, I.; Brown, F.H.; Fleagle, J.G. Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature 2005, 433, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Emery, N.J.; Clayton, N.S.; Frith, C.D. Social intelligence: From brain to culture. Phil. Trans. R. Soc. B 2007, 362, 485–488. [Google Scholar] [CrossRef]
- Humphrey, N.K. The social function of intellect. In Growing Points in Ethology; Bateson, P.P.G., Hinde, R.A., Eds.; Cambridge University Press: Cambridge, UK, 1976; pp. 303–317. [Google Scholar]
- Huxley, L. Scott’s Last Expedition in 2 Volumes, Volume 1 Being the Journals of R.F. Scott, R.N, C.V.O; Dodd: Mead, NY, USA, 1913. [Google Scholar]
- Ford, J.K.B.; Ellis, G.M.; Balcomb, K.C. Killer Whales: The Natural History and Genealogy of Orcinus Orca in British Columbia and Washington, 2nd ed.; University of Washington Press: Seattle, WA, USA, 2000. [Google Scholar]
- Hargrove, J.; Chua-Eoan, H. Beneath the Surface; Palgrave Macmillan: Basingstoke, UK, 2015. [Google Scholar]
- Silk, J.B. Using the “F”-word in primatology. Behaviour 2001, 139, 421–446. [Google Scholar] [CrossRef]
- Beetz, A.; Uvnäs-Moberg, K.; Julius, H.; Kotrschal, K. Psychosocial and psychophysiological effects of human-animal interactions: The possible role of oxytocin. Front. Psychol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Nagawawa, M.; Mogi, K.; Kikusui, T. Attachment between humans and dogs. Jpn. Psychol. Res. 2009, 51, 209–221. [Google Scholar] [CrossRef]
- Bayne, K. Development of the human-research animal bond and its impact on animal well-being. ILAR J. 2002, 43, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. The Descent of Man, and Selection in Relation to Sex; John Murray: London, UK, 1871. [Google Scholar]
- Burghardt, G.M. Amending Tinbergen: A fifth aim for ethology. In Anthropomorphism, Anecdotes, and Animals; Mitchell, R.W., Thompson, N.S., Miles, H.L., Eds.; SUNY Press: Albany, NY, USA, 1997; pp. 254–276. [Google Scholar]
- Tamietto, M.; de Gelder, B. Neural bases of the non-conscious perception of emotional signals. Nat. Rev. Neurosci. 2010, 11, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.V.; Proops, L.; Grounds, K.; Wathan, J.; McComb, K. Functionally relevant responses to human facial expressions of emotion in the domestic horse (Equus caballus). Biol. Lett. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Byrne, R.W.; Whiten, A. Computation and mindreading in primate tactical deception. In Natural Theories of Mind: Evolution, Development and Simulation of Everyday Mindreading; Whiten, A., Ed.; Basil Blackwell: Cambridge, UK, 1991; pp. 127–141. [Google Scholar]
- Mitchell, R.W.; Thompson, N.S. (Eds.) Deception, Perspectives on Human and Nonhuman Deceit; State University of New York Press: Albany, NY, USA, 1986.
- Fitzgerald, C.J.; Gerhart, J.I. Primates. In Encyclopedia of Deception; Levine, T.R., Ed.; SAGE Publications: Thousand Oaks, CA, USA, 2014; pp. 775–779. [Google Scholar]
- Riesch, R.; Deeke, V.B. Whistle communication in mammal-eating killer whales (Orcinus orca): Further evidence for acoustic divergence between ecotypes. Behav. Ecol. Sociobiol. 2010, 65, 1377–1387. [Google Scholar]
- Kirkpatrick, C. Tactical deception and the great apes: Insight into the question of theory of mind. Totem Univ. West. Ont. J. Anthropol. 2011, 15, 31–37. [Google Scholar]
- Schlinger, H.D., Jr. Theory of mind: An overview and behavioral perspective. Psychnol. Rec. 2009, 59, 435–448. [Google Scholar]
- Dvash, J.; Shamay-Tsoory, S.G. Theory of mind and empathy as multidimensional constructs—Neurological foundations. Top. Lang. Disord. 2014, 34, 282–295. [Google Scholar] [CrossRef]
- Ford, J.K.B.; Ellis, G.M. Selective foraging by fish-eating killer whales Orcinus orca in British Columbia. Mar. Ecol. Prog. Ser. 2006, 316, 185–199. [Google Scholar] [CrossRef]
- Kirby, D. Death at Seaworld; St. Martin’s Griffin: New York, NY, USA, 2012. [Google Scholar]
- Coscarella, M.A.; Bellazzi, G.; Gaffet, M.L.; Berzano, M.; Degrati, M. Short note: Technique used by killer whales (Orcinus orca) when hunting for dolphins in Patagonia, Argentina. Aquat. Mamm. 2015, 41, 192–197. [Google Scholar] [CrossRef]
- Bekoff, M.; Anderson, R.; Waayers, R.; Hockins, R.; Larrabee, L.; Garrett, H. Do Orcas Go Crazy Because of Petting Pools and False Hopes? Available online: https://www.psychologytoday.com/blog/animal-emotions/201503/do-orcas-go-crazy-because-petting-pools-and-false-hopes (accessed on 2 April 2016).
- Jacobs, S. Incidents by Orca. Available online: http://www.orcahome.de/incidentanalysis.htm (accessed on 14 August 2016).
- Robeck, T.R.; Monfort, S.L. Characterization of male killer whale (Orcinus orca) sexual maturation and reproductive seasonality. Theriogenology 2005, 66, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Olesiuk, P.F.; Ellis, G.M.; Ford, J.K.B. Life History and Population Dynamics of Northern Resident Killer Whales (Orcinus orca) in British Columbia; Canadian Science Advisory Secretariat: Nanaimo, BC, Canada, 2005. [Google Scholar]
- Robeck, T.R.; Willis, K.; Scarpuzzi, M.R.; O’Brien, J.K. Comparisons of life-history parameters between free-ranging and captive killer whale (Orcinus orca) populations for application toward species management. J. Mammal. 2015, 96, 1055–1070. [Google Scholar] [CrossRef] [PubMed]
- Latham, N.; Mason, G. Frustration and perseveration in stereotypic captive animals: Is a taste of enrichment worse than none at all? Behav. Brain Res. 2010, 211, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Rollin, B.E. An ethicist’s commentary on animal intelligence and animal welfare. Can. Vet. J. 2011, 52, 702, 704. [Google Scholar] [PubMed]
- Emery, N.J.; Clayton, N.S. Effects of experience and social context on prospective caching strategies by scrub jays. Nature 2001, 414, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Stroffrengen, T.A.; Pittenger, J.B. Human echolocation as a basic form of perception and action. Ecol. Psychol. 2010, 7. [Google Scholar] [CrossRef]
- Lilly, J.C. Lilly on Dolphins; Anchor Press/Doubleday: Garden City, NY, USA, 1975. [Google Scholar]
- Ridgway, S.; Carter, D. Features of dolphin skin with potential hydrodynamic importance. IEEE Eng. Med. Biol. 1993, 12, 83–88. [Google Scholar] [CrossRef]
- Kershenbaum, A.; Sayigh, L.S.; Janik, V.M. The encoding of individual identity in dolphin signature whistles: How much information is needed? PLoS ONE 2013, 8, e77671. [Google Scholar] [CrossRef] [PubMed]
- Anon. The Fate of Captive Orcas. Available online: http://us.whales.org/wdc-in-action/fate-of-captive-orcas (accessed on 14 August 2016).
- Roth, G.; Dicke, U. Evolution of the brain and intelligence. Trends Cogn. Sci. 2005, 9, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Marino, L. Cetacean brains: How aquatic are they? Anat. Rec. 2007, 290, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Wood, F.G.; Evans, W.E. Adaptiveness and ecology of echolocation in toothed whales. In Animal Sonar Systems; Busnel, R.G., Fish, J.F., Eds.; Plenum Press: New York, NY, USA, 1980; pp. 381–425. [Google Scholar]
- Marino, L. Cetacean brain evolution: Multiplication generates complexity. Int. J. Comp. Psychnol. 2004, 17, 1–16. [Google Scholar]
- Parfit, M.; Chisholm, S. The Lost Whale; St. Martin’s Press: New York, NY, USA, 2013. [Google Scholar]
| Average Incidents Per Orca */Average Incidents Per Orca ** | Group | Number in Group with Incidents/All Orcas in Group | Percent with Incidents |
|---|---|---|---|
| 7.57/3.31 | Petting Pool Orcas and Offspring | 7/16 | 44% |
| 3.67/2.20 | Tank Mates and Offspring | 3/5 | 60% |
| 3.50/3.50 | Orcas originally at Marineland | 2/2 | 100% |
| 2.40/1.50 | Sealand Orcas and Offspring | 5/8 | 62% |
| 2.00/0.67 | Joint Offspring of Tilikum and Petting Pool Orcas | 4/12 | 33% |
| 1.62/0.59 | Orcas with no Known Relationships | 8/22 | 38% |
| Sex | Maturity | Number in Group |
|---|---|---|
| M | Sexually Mature | 3 |
| F | Sexually Mature | 6 |
| M | In Puberty | 4 |
| F | In Puberty | 11 |
| M | Pre-puberty | 5 |
| F | Pre-Puberty | 7 |
| Sex | Maturity | Number in Group |
|---|---|---|
| M | Sexually Mature | 13 |
| F | Sexually Mature | 13 |
| M | In Puberty | 0 |
| F | In Puberty | 2 |
| M | Pre-puberty | 0 |
| F | Pre-Puberty | 1 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anderson, R.; Waayers, R.; Knight, A. Orca Behavior and Subsequent Aggression Associated with Oceanarium Confinement. Animals 2016, 6, 49. https://doi.org/10.3390/ani6080049
Anderson R, Waayers R, Knight A. Orca Behavior and Subsequent Aggression Associated with Oceanarium Confinement. Animals. 2016; 6(8):49. https://doi.org/10.3390/ani6080049
Chicago/Turabian StyleAnderson, Robert, Robyn Waayers, and Andrew Knight. 2016. "Orca Behavior and Subsequent Aggression Associated with Oceanarium Confinement" Animals 6, no. 8: 49. https://doi.org/10.3390/ani6080049
APA StyleAnderson, R., Waayers, R., & Knight, A. (2016). Orca Behavior and Subsequent Aggression Associated with Oceanarium Confinement. Animals, 6(8), 49. https://doi.org/10.3390/ani6080049