Swooping in the Suburbs; Parental Defence of an Abundant Aggressive Urban Bird against Humans

 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
- if lapwings can discriminate between different human activities, specifically between pedestrians and ground maintenance personnel mowing lawns;
- if the presence of eye stickers (mounted on the back of a person’s head) represents an effective way of managing lapwing attacks on people; and,
2. Methods
2.1. Study Area
2.2. Evoking Defensive Behaviour
2.3. Indexing Parental Defence
2.4. Nests, Eggs and Hatching Success
2.5. Data Analysis

{kind=link}
{kind=link}
| Variables | Low swooping and calling (PC1) | High swooping (PC2) | Separation (PC3) |
|---|---|---|---|
| Percentage of Variance Explained (%) | 34.0 | 22.9 | 16.3 |
| Response | 0.091 | 0.051 | 0.834 |
| Swoopabove | 0.124 | 0.914 | −0.011 |
| Swoopbelow | 0.826 | 0.307 | 0.018 |
| Swoop2above | 0.092 | 0.903 | 0.053 |
| Swoop2below | 0.860 | 0.013 | 0.038 |
| Calling | 0.667 | 0.027 | −0.035 |
| Proximity | −0.231 | −0.008 | 0.773 |
2.6. Characterising Defence
2.7. Effect of Habitat and Egg Age on Defence
| Response variable | Predictor variable | Df | F ratio | P |
|---|---|---|---|---|
| Low swooping and calling (Logged PC1) | Habitat | 1, 72 | 0.005 | 0.942 |
| R² = 0.050 | Days to hatching | 1, 72 | 0.384 | 0.528 |
| Laying date | 1, 72 | 1.709 | 0.195 | |
| Temperature (°C) | 1, 72 | 0.343 | 0.560 | |
| High swooping (Logged PC2) | Habitat | 1, 72 | 0.047 | 0.829 |
| R² = 0.040 | Days to hatching | 1, 72 | 0.392 | 0.533 |
| Laying date | 1, 72 | 2.087 | 0.153 | |
| Temperature | 1, 72 | 0.229 | 0.634 | |
| Separation (PC3) | Habitat | 1, 72 | 31.809 | < 0.001 |
| R² = 0.376 | Days to hatching | 1, 72 | 0.872 | 0.353 |
| Laying date | 1, 72 | 1.111 | 0.295 | |
| Temperature | 1, 72 | 0.997 | 0.321 |
3. Results
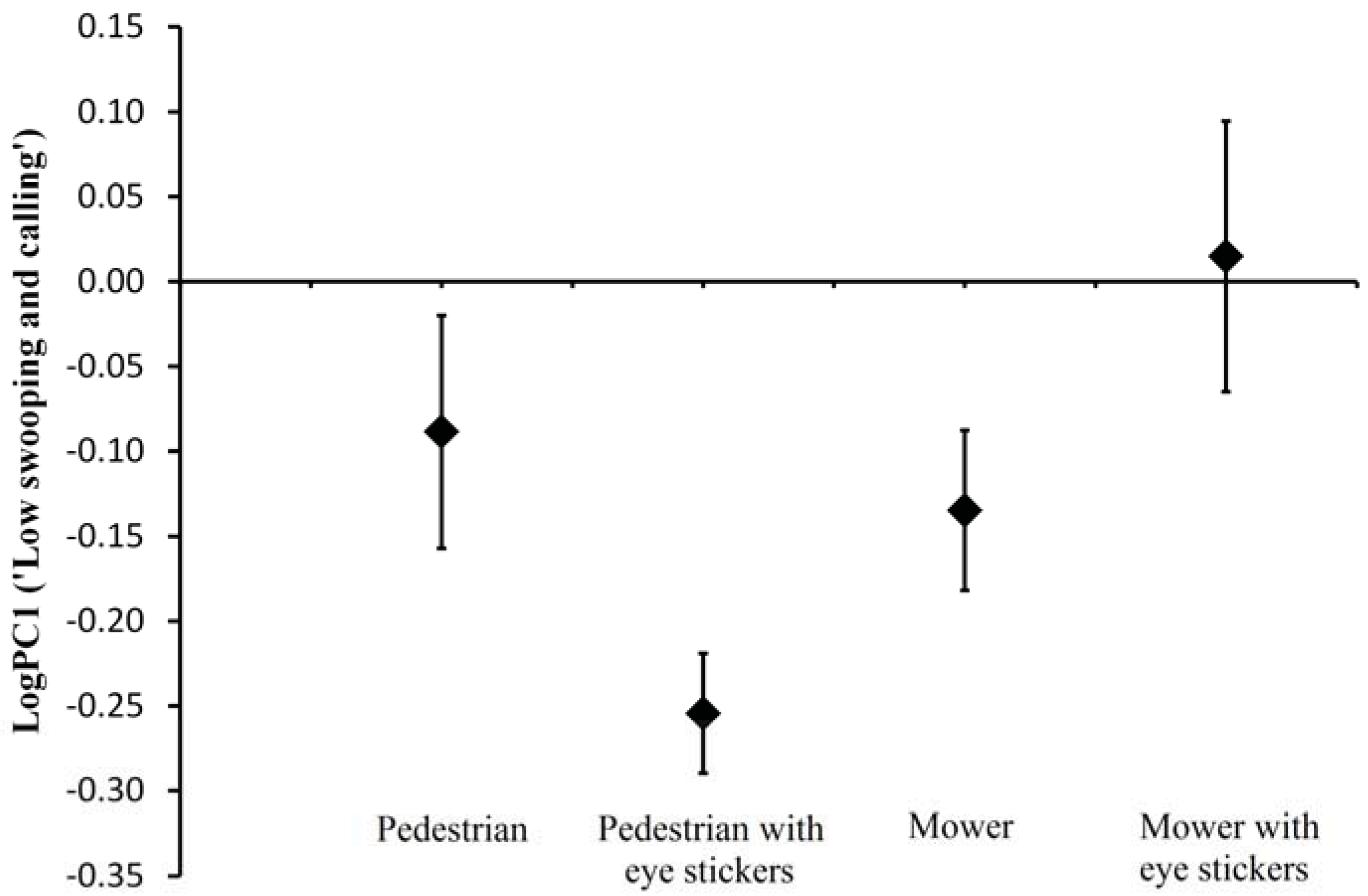
3.1. Effect of Stimulus on Defence
| Response variable | Predictor variable | Df | F ratio | p |
|---|---|---|---|---|
| Low swooping and calling (Logged PC1) | Mower/Pedestrian | 1, 73 | 3.378 | 0.070 |
| R² = 0.130 | Eyes/No eyes | 1, 73 | 0.018 | 0.895 |
| Mower/Pedestrian × Eyes/No eyes | 1, 73 | 6.758 | 0.011 | |
| High swooping (Logged PC2) | Mower/Pedestrian | 1, 73 | 0.165 | 0.686 |
| R² = 0.012 | Eyes/No eyes | 1, 73 | 0.247 | 0.621 |
| Mower/Pedestrian × Eyes/No eyes | 1, 73 | 0.639 | 0.427 | |
| Separation (PC3) | Mower/Pedestrian | 1, 73 | 10.237 | 0.002 |
| Eyes/No eyes | 1, 73 | 1.465 | 0.230 | |
| Mower/Pedestrian × Eyes/No eyes | 1, 73 | 1.362 | 0.247 |

3.2. Effect of Parental Defence on Hatching Success
| Response variable | Predictor variable | Df | C | SE | Z | P |
|---|---|---|---|---|---|---|
| Success | Low swooping and calling | 1, 71 | −0.201 | 0.247 | −0.81 | 0.415 |
| High swooping | 1, 71 | −0.103 | 0.181 | −0.57 | 0.568 | |
| Separation | 1, 71 | 0.611 | 0.351 | 1.74 | 0.082 | |
| Habitat | 1, 71 | 1.164 | 0.795 | 1.46 | 0.143 | |
| Days to hatching | 1, 71 | 0.014 | 0.031 | 0.48 | 0.634 | |
| Laying date | 1, 71 | −0.007 | 0.011 | −0.67 | 0.505 |
4. Discussion
5. Conclusion
Acknowledgements
Conflict of Interest
References
- Ricklefs, R.E. An Analysis of Nesting Mortality in Birds. Smithsonian Contrib. Zool. 1969, 9, 1–48. [Google Scholar] [CrossRef]
- Montgomerie, R.D.; Weatherhead, P.J. Risks and Rewards of Nest Defence by Parent Birds. The Quart. Rev. Biol. 1988, 63, 167–187. [Google Scholar]
- Trivers, R.L. Parental Investment and Sexual Selection, Sexual Selection and the Descent of Man; Campbell, B., Ed.; Aldine: Chicago, IL, USA, 1972; pp. 139–179. [Google Scholar]
- Brunton, D. The Effects of Nesting Stage, Sex, and Type of Predator on Parental Defense by Killdeer (Charadrius vociferous): Testing Models of Avian Parental Defense. Behav. Ecol. Sociobiol. 1990, 26, 181–190. [Google Scholar] [CrossRef]
- Buitron, D. Variability in the Responses of Black-Billed Magpies to Natural Predators. Behaviour 1983, 87, 209–236. [Google Scholar] [CrossRef]
- Galeotti, P.; Tavecchia, G.; Bonetti, A. Parental Defence in Long-eared Owls (Asio otus): Effects of Breeding Stage, Parent Sex and Human Persecution. J. Avian Biol. 2000, 31, 431–440. [Google Scholar]
- Gottfried, B.M. Anti-Predator Aggression in Birds Nesting in Old Field Habitats: An Experimental Analysis. The Condor 1979, 81, 251–257. [Google Scholar] [CrossRef]
- Jones, D.N.; Thomas, L.K. Attacks on Humans by Australian Magpies: Management of an Extreme Suburban Human-Wildlife Conflict. Wildlife Soc. Bull. 1999, 27, 473–478. [Google Scholar]
- Warne, R.M.; Jones, D.N. Evidence of Target Specificity in Attacks by Australian Magpies on Humans. Wildlife Res. 2003, 30, 265–267. [Google Scholar] [CrossRef]
- Jones, D.N.; Nealson, T. Management of Aggressive Australian Magpies by Translocation. Wildlife Res. 2003, 30, 167–177. [Google Scholar] [CrossRef]
- Wallin, K. Defence as Parental Care in Tawny Owls (Strix aluco). Behaviour 1987, 102, 213–230. [Google Scholar] [CrossRef]
- Kofron, C.P. Attacks to Humans and Domestic Animals by the Southern Cassowary (Casuarius casuarius johnsonii) in Queensland, Australia. J. Zool. 1999, 249, 375–381. [Google Scholar] [CrossRef]
- Cilento, N.J.; Jones, D.N. Aggression by Australian Magpies (Gymnorhina tibicen) Toward Human Intruders. Emu 1999, 99, 85–90. [Google Scholar] [CrossRef]
- Buchman, B.; Wilson, M.; Holland, S. Swoop! Available online: http://www.dse.vic.gov.au/plants-and-animals/native-plants-and-animals/problem-wildlife/swooping-birds (accessed on 9 August 2013).
- Cardilini, A.P.A. The Effects of Urbanisation on Reproductive Success and Parental Care in the Masked Lapwing (Vanellus miles). Deakin University Unpublished Honours MS: Waurn Ponds, Australia, 2010. [Google Scholar]
- Marchant, S.; Higgins, P.J. Handbook of Australian, New Zealand and Antartic Birds; Oxford University Press Australia: Melbourne, Australia, 1993; Volume 2. [Google Scholar]
- Glover, H.K. Deakin University. Persional communication, 2012. [Google Scholar]
- Glover, H.K.; Weston, M.A.; Maguire, G.S.; Miller, K.K.; Christie, B.A. Towards Ecologically Meaningful and Socially Acceptable Buffers: Response Distances of Shorebirds in Victoria, Australia, to Human Disturbance. Landscape Urban Plan. 2011, 103, 326–334. [Google Scholar] [CrossRef]
- Cardilini, A.P.A.; Weston, M.A.; Nimo, D.G.; Dann, P.; Sherman, C.D.H. Surviving in Sprawling Suburbs: Suburban Environments Represent High Quality Breeding Habitat for a Widespread Shorebird. Landscape Urban Plan. 2013, 115, 72–80. [Google Scholar] [CrossRef]
- Hampel, T. Pesky Plovers. Dealing with Swooping Birds. Available online: http://www.trevorsbirding.com/pesky-plovers-dealing-with-swooping-birds/ (accessed on 9 August 2013).
- Ristau, C.A. Language, Cognition, and Awareness in Animals? Ann. N. Y. Acad. Sci. 1983, 406, 170–186. [Google Scholar] [CrossRef]
- Bugnyar, T.; Stöwe, M.; Heinrich, B. Ravens, Corvus corax, Follow Gaze Direction of Humans Around Obstacles. Proc. Roy. Soc. Lond. B Biol. Sci. 2004, 271, 1331–1336. [Google Scholar] [CrossRef]
- Jones, D.N. Living with a Dangerous Neighbour: Australian Magpies in a Suburban Environment. In Proceedings of the 4th International Symposium on Urban Wildlife Conservation, Tucson, AZ, USA, 1–5 May 1999.
- Andersson, M.; Wiklund, C.G.; Rundgren, H. Parental Defence of Offspring: A Model and an Example. Anim. Behav. 1980, 28, 536–542. [Google Scholar] [CrossRef]
- Arnold, K.E. Group Mobbing Behaviour and Nest Defence in a Cooperatively Breeding Australian Bird. Ethology 2000, 106, 385–393. [Google Scholar] [CrossRef]
- Dann, P. Breeding of the Banded and Masked Lapwings in Southern Victoria. Emu 1981, 81, 121–127. [Google Scholar] [CrossRef]
- Thomas, D.G. Breeding Biology of the Australian Spur-winged Plover. Emu 1969, 69, 81–102. [Google Scholar] [CrossRef]
- Chambers, L.E.; Gibbs, H.; Weston, M.A.; Ehmke, G.C. Spatial and Temporal Variation in the Breeding of Masked Lapwings (Vanellus miles) in Australia. Emu 2008, 108, 115–124. [Google Scholar] [CrossRef]
- Weston, M.A.; Dodge, F.; Bunce, A.; Nimmo, D.G.; Miller, K.K. Do Temporary Beach Closures Assist in the Conservation of Breeding Shorebirds on Recreational Beaches? Pac. Conserv. Biol. 2012, 18, 47–55. [Google Scholar]
- Liebezeit, J.R.; Smith, P.A.; Lanctot, R.B.; Schekkerman, H.; Tulp, I.; Kendall, S.J.; Tracy, D.M.; Rodrigues, R.J.; Meltofte, H.; Robinson, J.A.; Gratto-Trevor, C.; McCaffery, B.J.; Morse, J.; Zack, S.W. Assessing the Development of Shorebird Eggs Using the Floatation Method: Spesies-specific and Generalized Regression Models. The Condor 2007, 109, 32–47. [Google Scholar] [CrossRef]
- Székely, T.; Kosztolányi, A.; Küpper, C. Practical Guide for Investigating Breeding Ecology of Kentish Plover (Charadrius alexandrinus); University of Bath: Bath, UK, 2008; Volume 3. [Google Scholar]
- Lees, D.; Weston, M.A.; Sherman, C.; Maguire, G.; Dann, P.; Cardilini, A.P.; Tan, L. Occurrences of Cooperative Breeding in the Masked Lapwing Vanellus miles. The Victorian Naturalist 2013, 130, 84–85. [Google Scholar]
- Giles, N.; Huntingford, F.A. Predation Risk and Inter-population Variation in Antipredator Behaviour in the Three-spined Stickleback, Gasterosteus aculeatus. Anim. Behav. 1984, 32, 264–275. [Google Scholar] [CrossRef]
- Huntingford, F.A. An Investigation of the Territorial Behaviour of the Three-spined Stickleback (Gasterosteus aculeatus) Using Principal Components Analysis. Anim. Behav. 1976, 24, 822–834. [Google Scholar] [CrossRef]
- SPSS, SPSS Stastics, 20.0; SPSS Inc.: Chicago, IL, USA, 2011.
- Lord, A.; Waas, J.R.; Innes, J.; Whittingham, M.J. Effects of Human Approaches to Nests of Northern New Zealand Dotterels. Biol. Conserv. 2001, 98, 233–240. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Walls, J.; Cornell, H.N.; Withey, J.C.; Craig, D.P. Lasting Recognition of Threatening People by Wild American Crows. Anim. Behav. 2010, 79, 699–707. [Google Scholar] [CrossRef]
- Stankowich, T.; Blumstein, D.T. Fear in Animals: A Meta-analysis and Review of Risk Assessment. Proc. Roy. Soc. B Biol. Sci. 2005, 272, 2627–2634. [Google Scholar] [CrossRef]
- Lees, D. Deakin University. Persional communication, 2013. [Google Scholar]
- Lee, W.Y.; Choe, J.C.; Jablonski, P.G. Wild Birds Recognize Individual Humans: Experiments on Magpies, Pica pica. Anim. Cogn. 2011, 14, 817–825. [Google Scholar] [CrossRef]
- Møller, A.P. Flight Distance of Urban Birds, Predation, and Selection for Urban Life. Behav. Ecol. Sociobiol. 2008, 63, 63–75. [Google Scholar] [CrossRef]
- Brunton, D. Fatal Antipredator Behavior of a Killdeer. Wilson Bull. 1986, 98, 605–607. [Google Scholar]
- Sordhal, T.A. Evolutionary Aspects of Avian Distraction Display: Variation in American Avocet and Black-necked Stilt Antipredator Behaviour. In Deception: Perspectives on Human and Nonhuman Deceit; Mitchell, R.W., Ed.; SUNY Press: Albany, NY, USA, 1986; pp. 87–107. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lees, D.; Sherman, C.D.H.; Maguire, G.S.; Dann, P.; Cardilini, A.P.A.; Weston, M.A. Swooping in the Suburbs; Parental Defence of an Abundant Aggressive Urban Bird against Humans. Animals 2013, 3, 754-766. https://doi.org/10.3390/ani3030754
Lees D, Sherman CDH, Maguire GS, Dann P, Cardilini APA, Weston MA. Swooping in the Suburbs; Parental Defence of an Abundant Aggressive Urban Bird against Humans. Animals. 2013; 3(3):754-766. https://doi.org/10.3390/ani3030754
Chicago/Turabian StyleLees, Daniel, Craig D. H. Sherman, Grainne S. Maguire, Peter Dann, Adam P. A. Cardilini, and Michael A. Weston. 2013. "Swooping in the Suburbs; Parental Defence of an Abundant Aggressive Urban Bird against Humans" Animals 3, no. 3: 754-766. https://doi.org/10.3390/ani3030754
APA StyleLees, D., Sherman, C. D. H., Maguire, G. S., Dann, P., Cardilini, A. P. A., & Weston, M. A. (2013). Swooping in the Suburbs; Parental Defence of an Abundant Aggressive Urban Bird against Humans. Animals, 3(3), 754-766. https://doi.org/10.3390/ani3030754