An Attempt at Captive Breeding of the Endangered Newt Echinotriton andersoni, from the Central Ryukyus in Japan

Abstract
:Simple Summary
Abstract

1. Introduction

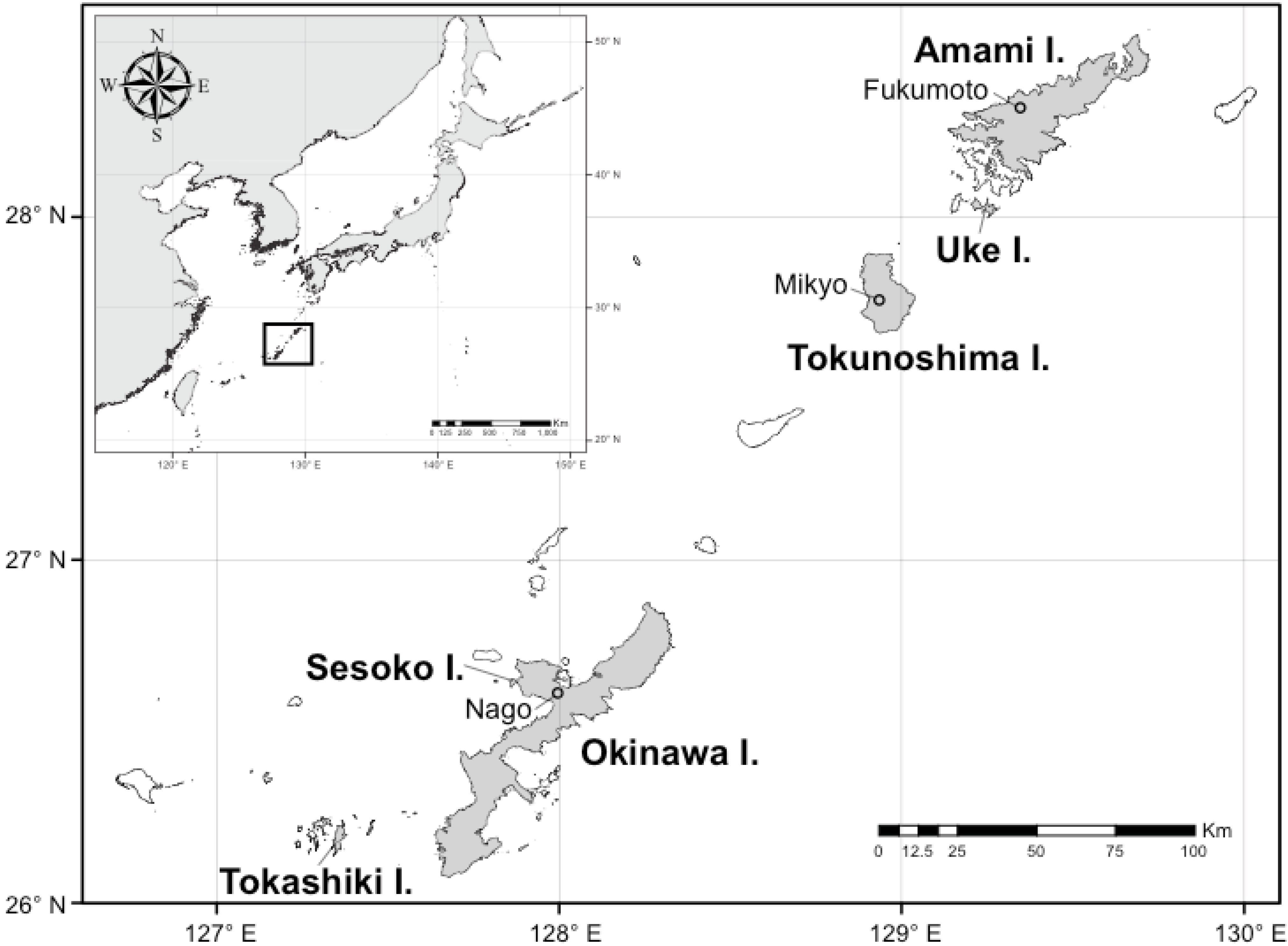
2. Materials and Methods




3. Results
3.1. Development of Naturally Deposited Embryos
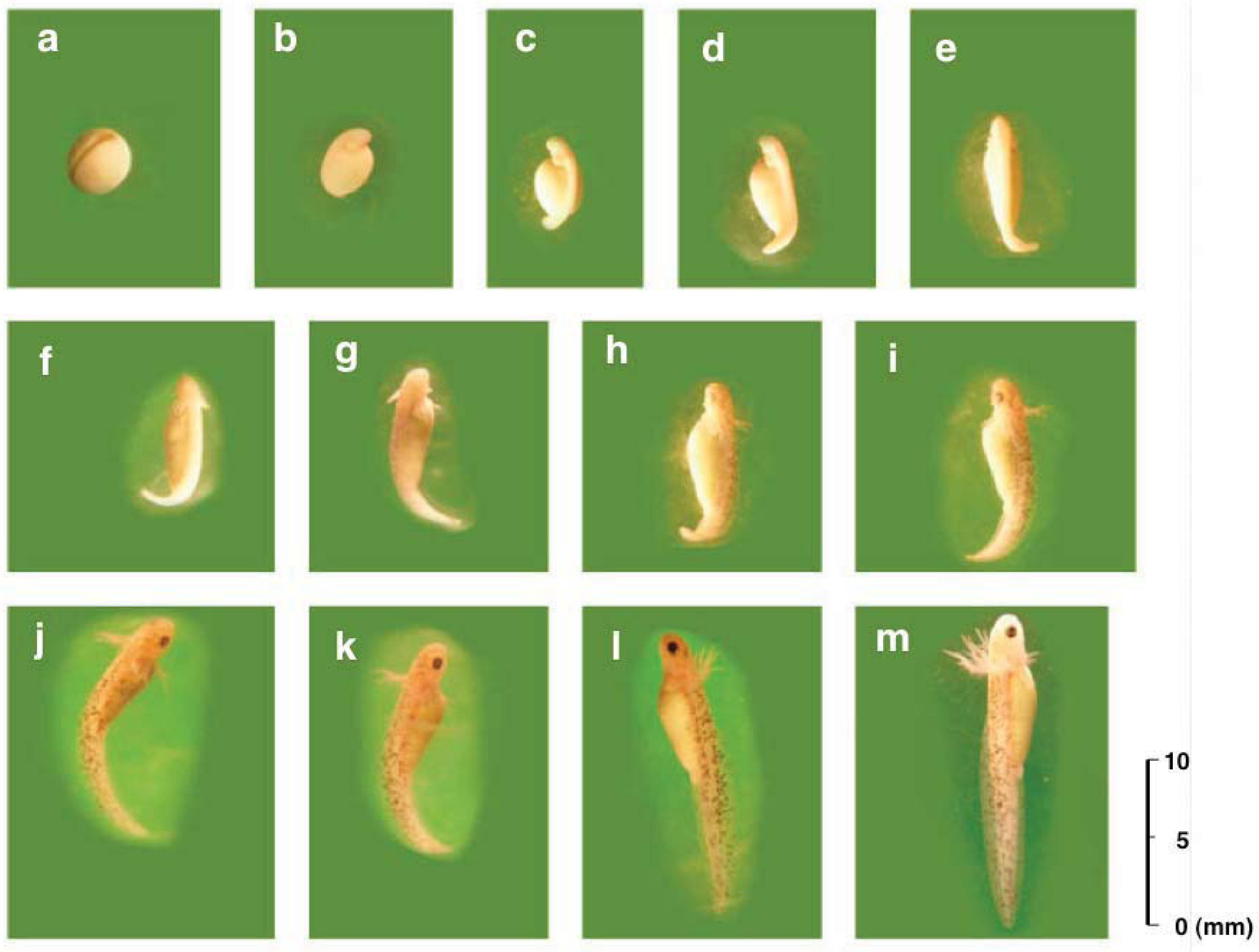
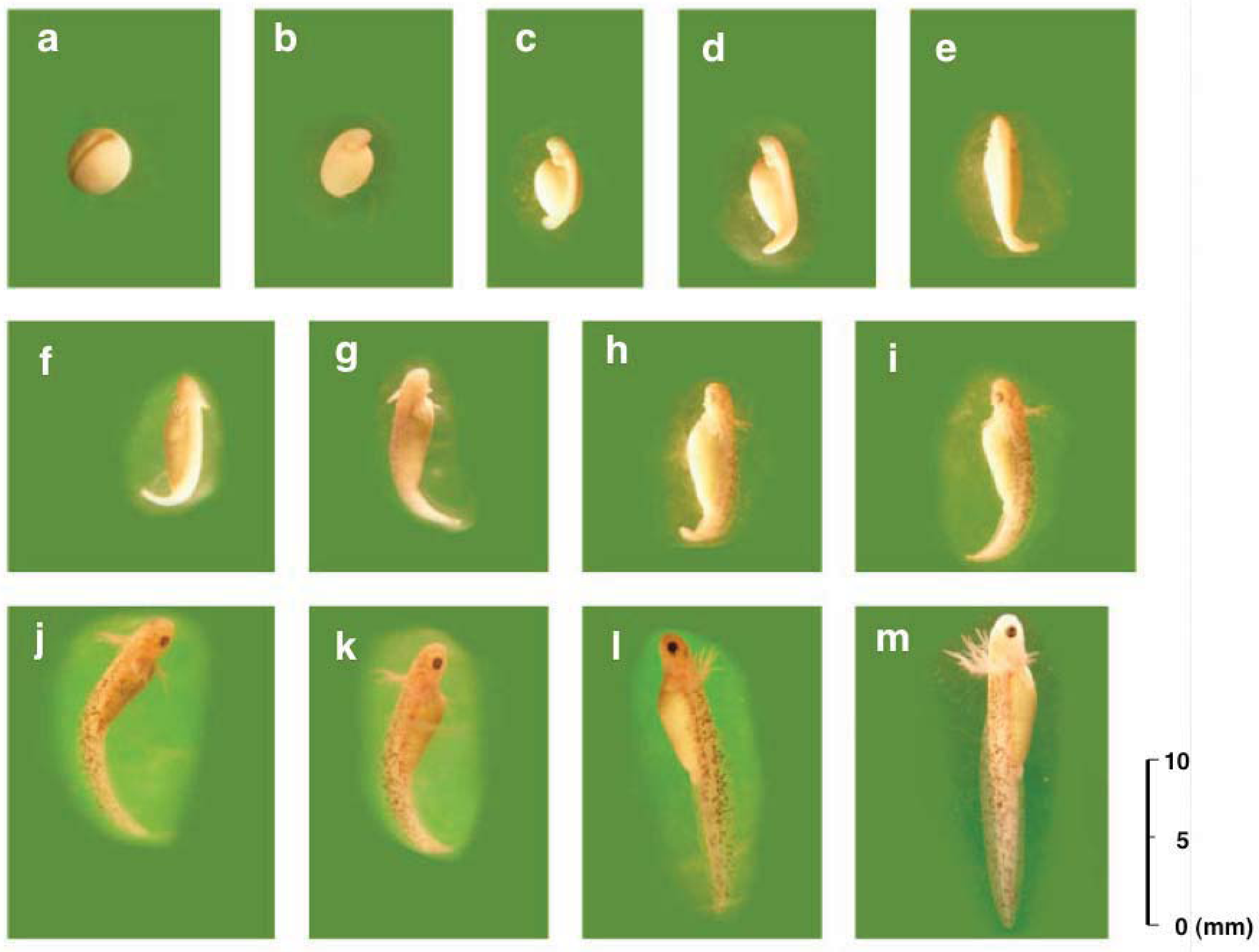
3.2. Developmental Capacity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
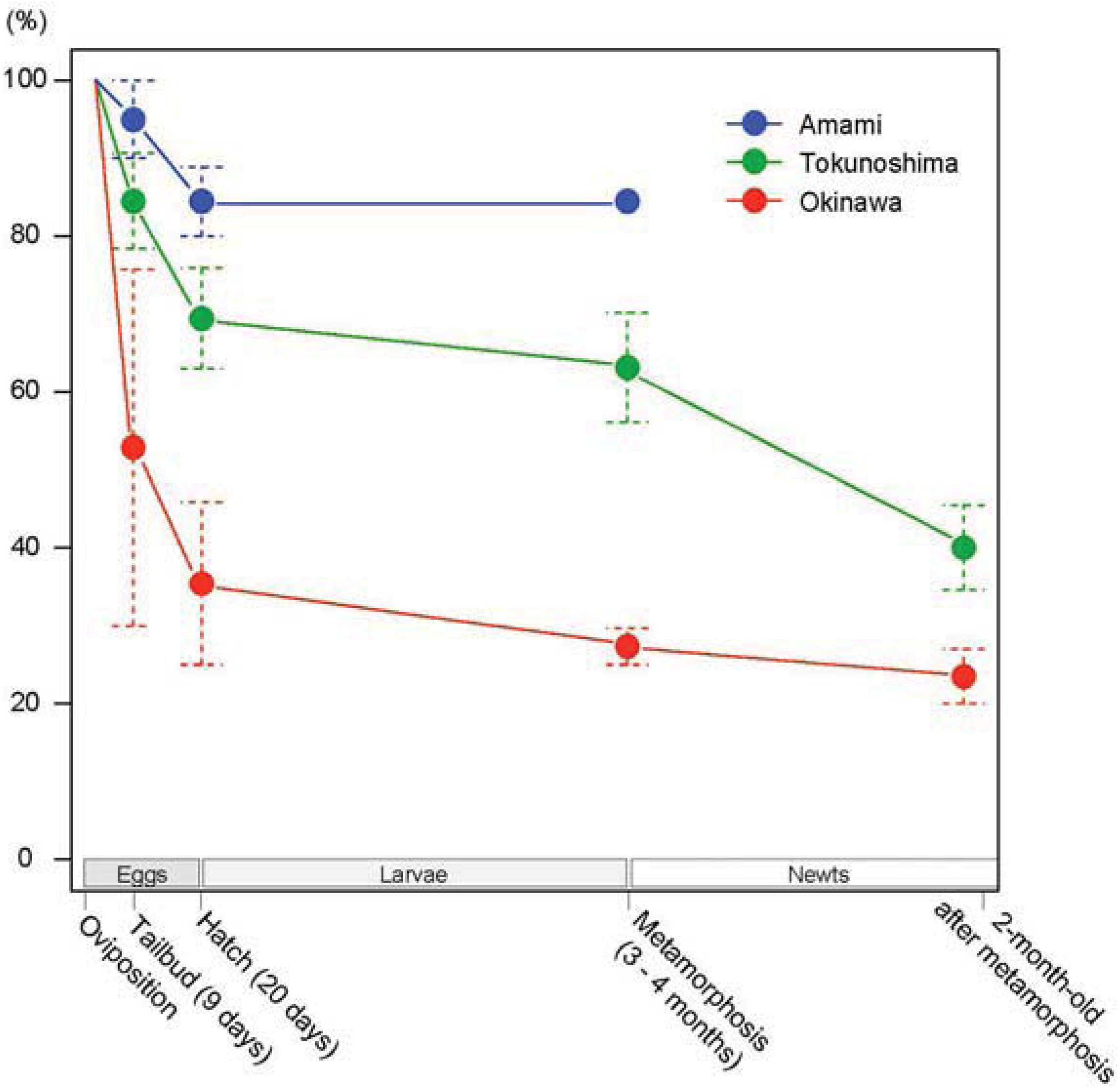
| Island | Date | No. of Eggs | Normal Tailbud Embryos (%) | Normally Hatched Larvae (%) | Metamor- phosed Newts (%) | 2-Month-Old Newts (%) |
|---|---|---|---|---|---|---|
| Okinawa | 2009.4.19 | 37 | 28 (75.7) | 17 (45.9) | 11 (29.7) | 10 (27.0) |
| 2010.4.10 | 20 | 6 (30.0) | 5 (25.0) | 5 (25.0) | 4 (20.0) | |
| Total | 57 | 34 (59.6) | 22 (38.6) | 16 (28.1) | 14 (24.6) | |
| Tokunoshima | 2010.2.22 | 33 | 29 (87.9) | 29 (87.9) | 29 (87.9) | 22 (66.7) |
| 2010.3.23 | 21 | 21 (100) | 21 (100) | 21 (100) | 16 (76.2) | |
| 2010.4.12 | 30 | 25 (83.3) | 24 (80.0) | 23 (76.7) | 13 (43.3) | |
| 2010.4.16 | 17 | 9 (52.9) | 9 (52.9) | 9 (52.9) | 4 (23.5) | |
| 2010.6. 4 | 36 | 25 (69.4) | 25 (69.4) | 25 (69.4) | 14 (38.9) | |
| 2011.1.23 | 44 | 37 (61.4) | 37 (61.4) | 25 (56.8) | 14 (31.8) | |
| 2011.3.11 | 20 | 20 (100) | 10 (50.0) | 10 (50.0) | 6 (30.0) | |
| 2011.4.13 | 3 | 3 (100) | 3 (100) | 3 (100) | 2 (66.7) | |
| 2011.4.30 | 10 | 10 (100) | 5 (50.0) | 4 (40.0) | 2 (20.0) | |
| 2011.5. 7 | 17 | 17 (100) | 7 (41.2) | 4 (23.5) | 2 (11.8) | |
| 2011.5.29 | 7 | 7 (100) | 6 (85.7) | 6 (85.7) | 4 (57.1) | |
| 2012.3. 7 | 46 | 46 (100) | 44 (95.7) | 38 (82.6) | 28 (60.9) | |
| Total | 284 | 249 (87.7) | 220 (77.5) | 197 (69.4) | 127 (44.7) | |
| Amami | 2013.4.17 | 27 | 27(100) | 24(88.9) | 24(88.9) | – |
| 2013.5.21 | 10 | 9(90.0) | 8(80.0) | – | – | |
| Total | 37 | 36 (97.3) | 32 (865) | – | – | |


4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Utsunomiya, T. Searching for a ‘living fossil’, Tylotriton andersoni. (in Japanese). Asahi-Larousse Weekly Animal Encyclopedia 1973, 144, I–III. [Google Scholar]
- Matayoshi, S.; Oshiro, N.; Kiyuna, T.; Hoshikawa, H.; Mitsui, K.; Kumagai, E. A note on the distribution of Tylotriton andersoni Boulenger in Okinawa Island. (in Japanese). Biol. Mag. Okinawa 1997, 15, 1–4. [Google Scholar]
- Zhao, E-M.; Adler, K. Herpetology of China; Society for the Study of Amphibians and Reptiles: Oxford, OH, USA, 1993; p. 111. [Google Scholar]
- Zhao, E. China Red Data Book of Endangered Animals: Amphibia and Reptilia; Science Press: Beijing, China, 1998. [Google Scholar]
- Matsui, M.; Ota, H. On Chinese herpetology. Herpetologica 1995, 51, 234–250. [Google Scholar]
- Yamada, F.; Sugimura, K. Negative impact of an invasive small Indian mongoose Herpestes javanicus on native wildlife species and evaluation of a control project in Amami-Ohshima and Okinawa Islands. Glob. Environ. Res. (Japan) 2004, 8, 117–124. [Google Scholar]
- IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org (accessed on 25 July 2013).
- Uchiyama, R.; Maeda, N.; Numata, K.; Seki, S. A Photograpic Guide. In Amphibians and Reptiles in Japan(in Japanese), 4th ed.; Heibonsha: Tokyo, Japan, 2005; pp. 62–63. [Google Scholar]
- Matayoshi, S.; Oshiro, N.; Kiyuna, T.; Hoshikawa, H.; Mitsui, K.; Kumagai, E. A note on the spawning of Tylotriton andersoni Boulenger in Okinawa Island. (in Japanese). Biol. Mag. Okinawa 1978, 16, 11–16. [Google Scholar]
- Tanaka, S. Preliminary observations on ecology of the subtropical newts Tylotriton (Echinotriton) andersoni Tylotriton and Cynopus ensicauda in Sesokojima Island, the Ryukyu Islands. (in Japanese). J. Biol. Educ. 1994, 26, 13–21. [Google Scholar]
- Utsunomiya, T.; Matsui, M. Male courtship behavior of Tylotriton (Echinotriton) andersoni Boulenger under laboratory condition. Current Herpetol. 2002, 21, 67–74. [Google Scholar] [CrossRef]
- Sugawara, H.; Igawa, T.; Iwai, N.; Yamamuro, K.; Oumi, S.; Sumida, M. Record of the egg-laying behavior of Anderson’s crocodile newt (Echinotriton andersoni) from Tokunoshima (in Japanese). Bull. Herpetol. Soc. Japan 2011, 2011, 115–118. [Google Scholar]
- Fleck, J. Bemerkungen zu Echinotriton andersoni von den Japanischen Inseln Tokunoshima und Okinawa sowie zur Nachzucht von Tieren der Tokunoshima-Form. Amphibia 2006, 5, 4–6. [Google Scholar]
- Snider, A.; Zippel, K. Amphibian Conservation at the Detroit Zoological Institute. Froglog 2000, 40, 2. [Google Scholar]
- Kuramoto, M.; Satou, N.; Oumi, S.; Kurabayashi, A.; Sumida, M. Inter- and intra-island divergence in Odorrana ishikawae (Anura, Ranidae) of the Ryukyu Archipelago of Japan, with description of a new species. Zootaxa 2011, 2767, 25–40. [Google Scholar]
- Matsui, M. On the brown frogs from the Ryukyu Archipelago, Japan, with descriptions of two new species (Amphibia, Anura). Current Herpetol. 2012, 30, 111–128. [Google Scholar] [CrossRef]
- Igawa, T.; Oumi, S.; Katsuren, S.; Sumida, M. Population structure and landscape genetics of two endangered frog species of genus Odorrana: Different scenarios on two islands. Heredity 2013, 110, 46–56. [Google Scholar] [CrossRef]
- Hayashi, T.; Matsui, M.; Utsunomiya, T.; Tanaka, S.; Ota, H. Allozyme variation in the newt Tylototriton andersoni from three islands of the Ryukyu Archipelago. Herpetologica 1992, 48, 178–184. [Google Scholar]
- Kurabayashi, A.; Nishitani, T.; Katsuren, S.; Oumi, S.; Sumida, M. Mitochondrial genomes and divergence times of crocodile newts: Inter-islands distribution of Echinotriton andersoni and the origin of a unique repetitive sequence found in Tylototriton mt genomes. Genes Genet. Syst. 2012, 87, 39–51. [Google Scholar] [CrossRef]
- Honda, M.; Matsui, M.; Tominaga, A.; Ota, H.; Tanaka, S. Phylogeny and biogeography of the Anderson’s crocodile newt, Echinotriton andersoni (Amphibia: Caudata), as revealed by mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2012, 65, 642–653. [Google Scholar] [CrossRef]
- Sugawara, H.; Igawa, T.; Yokogawa, M.; Okuda, M.; Oumi, S.; Katsuren, S.; Kaneko, S.; Umino, T.; Isagi, Y.; Sumida, M. Isolation and characterization of ten microsatellite loci of endangered Anderson’s crocodile newt. Echinotriton andersoni. Conserv. Genet. Resour. 2012, 4, 595–598. [Google Scholar] [CrossRef]
- Igawa, T.; Sugawara, H.; Honda, M.; Tominaga, A.; Oumi, S.; Katsuren, S.; Ota, H.; Matsui, M.; Sumida, M. Fine scale population structure and genetic divergence of Anderson’s crocodile newt, Echinotriton andersoni. 2013. in preparation. [Google Scholar]
- Utsunomiya, Y.; Utsunomiya, T. On the development of Tylototriton andersoni. (in Japanese). J. Fac. Fish. Anim. Husb. 1997, 16, 65–76. [Google Scholar]
- Nishioka, M. Amphibians. In Lecture for Experimental Biology. Method for Preparing Experimental Materials; (in Japanese). Egami, N., Katsumi, M., Eds.; Maruzen Co.: Tokyo, Japan, 1982; Volume 1, pp. 75–94. [Google Scholar]
- Sumida, M.; Satou, N.; Yoshikawa, N.; Kurabayashi, A.; Islam, M.M.; Igawa, T.; Oumi, S.; Katsuren, S.; Ota, H.; Shintani, N.; Fukuniwa, H.; Sano, N.; Fujii, T. Artificial production and natural breeding of the endangered frog species Odorrana ishikawae, with special reference to fauna conservation in the laboratory. Zool. Sci. 2011, 28, 834–839. [Google Scholar] [CrossRef]
- Utsunomiya, T. Data Book on Rare Wild Aquatic Organism in Japan; (in Japanese). Japan Fisheries Resource Conservation Association: Tokyo, Japan, 1998; pp. 220–221. [Google Scholar]
- Orton, F.; Routledge, E. Agricultural intensity in ovo affects growth, metamorphic development and sexual differentiation in the common toad (Bufo bufo). Ecotoxicology 2011, 20, 901–911. [Google Scholar] [CrossRef]
- Maeda, N.; Matsui, M. Frogs and Toads of Japan; Bunichi-Sogo Shuppan: Tokyo, Japan, 1999. [Google Scholar]
- Ota, H. Anderson’s crocodile newt. In Endangered Wild Animals and Plants in Kagoshima Prefecture; (in Japanese). Kagoshima Environmental Research and Service: Kagoshima, Japan, 2003; p. 105. [Google Scholar]
- Matsui, M.; Shimada, T.; Ota, H.; Tanaka-Ueno, T. Multiple invasions of the Ryukyu archipelago by Oriental frogs of the subgenus Odorrana with phylogenetic reassessment of the related subgenera of the genus Rana. Mol. Phylogenet. Evol. 2005, 37, 733–742. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Igawa, T.; Sugawara, H.; Tado, M.; Nishitani, T.; Kurabayashi, A.; Islam, M.M.; Oumi, S.; Katsuren, S.; Fujii, T.; Sumida, M. An Attempt at Captive Breeding of the Endangered Newt Echinotriton andersoni, from the Central Ryukyus in Japan. Animals 2013, 3, 680-692. https://doi.org/10.3390/ani3030680
Igawa T, Sugawara H, Tado M, Nishitani T, Kurabayashi A, Islam MM, Oumi S, Katsuren S, Fujii T, Sumida M. An Attempt at Captive Breeding of the Endangered Newt Echinotriton andersoni, from the Central Ryukyus in Japan. Animals. 2013; 3(3):680-692. https://doi.org/10.3390/ani3030680
Chicago/Turabian StyleIgawa, Takeshi, Hirotaka Sugawara, Miyuki Tado, Takuma Nishitani, Atsushi Kurabayashi, Mohammed Mafizul Islam, Shohei Oumi, Seiki Katsuren, Tamotsu Fujii, and Masayuki Sumida. 2013. "An Attempt at Captive Breeding of the Endangered Newt Echinotriton andersoni, from the Central Ryukyus in Japan" Animals 3, no. 3: 680-692. https://doi.org/10.3390/ani3030680