
Replacing up to 50% of Corn Silage with Triticale Silage Alters the Fecal Microbiome but Not Milk Yield or Composition in Mid-Lactation Holstein Cows


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Location and Ethical Approval
2.2. Silage Management and Chemical Composition
2.3. Cows, Experimental Design, and Feeding Management
2.4. Sample Collection, Laboratory Analyses, and Measurements
2.4.1. Silages and Total Mixed Diets
2.4.2. Milk Yield, Milk Composition and Related Parameters
2.4.3. Fecal Microbiota
2.4.4. Fecal Volatile Fatty Acids
2.5. Statistical Analysis
3. Results
3.1. Effects of Partially Replacing Corn Silage with Triticale Silage on Milk Yield and 4% FCM
3.2. Effects of Partially Replacing Corn Silage with Triticale Silage on Milk Quality of Lactating Holstein Cows
3.3. Effects of Replacing Corn Silage with Triticale Silage on the Fecal Microbiota of Dairy Cows
3.3.1. Sequencing Depth and OTU Distribution
3.3.2. Alpha Diversity
3.3.3. Beta Diversity
3.3.4. Taxonomic Composition and Differential Relative Abundance
3.4. Fecal Volatile Fatty Acids
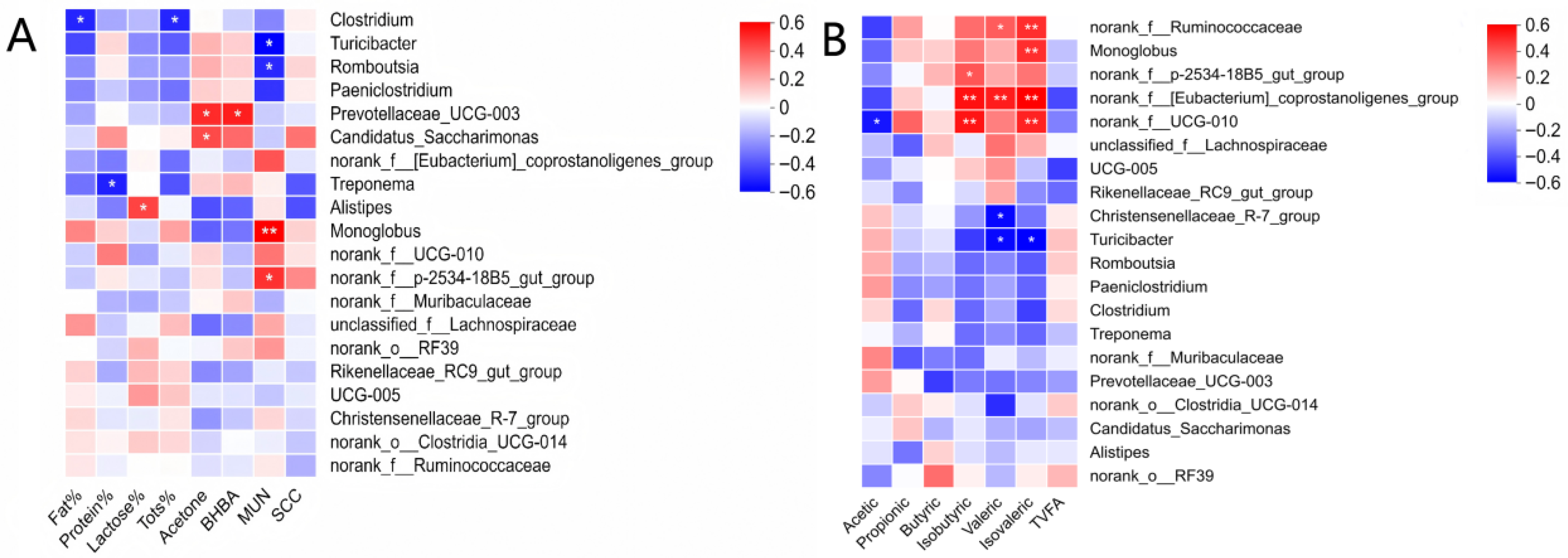
3.5. Correlations Between Fecal Microbiota, Milk Traits, and Fecal Volatile Fatty Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, C.; Cao, Z.; Wu, H.; Wang, B.; Niu, D.; Zheng, M.; Jiang, D.; Xiao, J. The Status of the Forage Utilization Industry in China. In Research Progress on Forage Production, Processing and Utilization in China; Yang, F., Ni, K., Guo, X., Eds.; Springer: Singapore, 2022; pp. 183–207. [Google Scholar] [CrossRef]
- Bach, A.; Joulie, I.; Chevaux, E.; Elcoso, G.; Ragués, J. Milk performance and rumen microbiome of dairy cows as affected by the inclusion of corn silage or corn shredlage in a total mixed ration. Animal 2021, 15, 100014. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.T.; Oh, J.; Giallongo, F.; Roth, G.W.; Hristov, A.N. Inclusion of wheat and triticale silage in the diet of lactating dairy cows. J. Dairy Sci. 2017, 100, 6151–6163. [Google Scholar] [CrossRef] [PubMed]
- Plaizier, J.C.; Krause, D.O.; Gozho, G.N.; McBride, B.W. Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. Vet. J. 2008, 176, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Zebeli, Q.; Aschenbach, J.R.; Tafaj, M.; Boguhn, J.; Ametaj, B.N.; Drochner, W. Invited review: Role of physically effective fiber and estimation of dietary fiber adequacy in high-producing dairy cattle. J. Dairy Sci. 2012, 95, 1041–1056. [Google Scholar] [CrossRef]
- Ayalew, H.; Kumssa, T.T.; Butler, T.J.; Ma, X.F. Triticale improvement for forage and cover crop uses in the Southern Great Plains of the United States. Front. Plant Sci. 2018, 9, 1130. [Google Scholar] [CrossRef]
- Giuliotti, L.; Benvenuti, M.N.; Martini, A.; Accorsi, P.A.; Lotti, C.; Cappucci, A.; Conte, G. Assessment of blood and productive parameters in mid-lactation dairy cows fed different diets: Replacement of corn silage with triticale silage. Arch. Anim. Breed. 2022, 65, 223–229. [Google Scholar] [CrossRef]
- Iommelli, P.; Zicarelli, F.; Amato, R.; Musco, N.; Sarubbi, F.; Bailoni, L.; Lombardi, P.; Di Bennardo, F.; Infascelli, F.; Tudisco, R. The Effects of Hemp Hay (Cannabis sativa L.) in the Diets of Grazing Goats on Milk Production and Fatty Acid Profile. Animals 2024, 14, 2373. [Google Scholar] [CrossRef]
- Amato, R.; Oteri, M.; Chiofalo, B.; Zicarelli, F.; Musco, N.; Sarubbi, F.; Pacifico, S.; Formato, M.; Lombardi, P.; Di Bennardo, F.; et al. Diet Supplementation with Hemp (Cannabis sativa L.) Inflorescences: Effects on Quanti-Qualitative Milk Yield and Fatty Acid Profile on Grazing Dairy Goats. Vet. Q. 2024, 44, 1–8. [Google Scholar] [CrossRef]
- Hosseinian, F.S.; Mazza, G. Triticale bran and straw: Potential new sources of phenolic acids, proanthocyanidins, and lignans. J. Funct. Foods 2009, 1, 57–64. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census Collaborators; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef]
- Ricci, S.; Rivera-Chacon, R.; Petri, R.M.; Reisinger, N.; Zebeli, Q.; Castillo-Lopez, E. Progressive microbial adaptation of the bovine rumen and hindgut in response to increasing amounts of starch with or without phytogenic feed additive. Front. Microbiol. 2022, 13, 920427. [Google Scholar] [CrossRef]
- Rivera-Chacon, R.; Pacífico, C.; Ricci, S.; Petri, R.M.; Reisinger, N.; Zebeli, Q.; Castillo-Lopez, E. Prolonged feeding of high-concentrate diet remodels the hindgut microbiome and modulates nutrient degradation in the rumen and the total gastrointestinal tract of cows. J. Dairy Sci. 2024, 107, 9235–9250. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle: Seventh Revised Edition, 2001; National Academies Press: Washington, DC, USA, 2001. [CrossRef]
- AOAC International. Official Methods of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2002. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Gaines, W.L.; Davidson, F.A. Relation Between Percentage Fat Content and Yield of Milk: Correction of Milk Yield for Fat Content; University of Illinois Agricultural Experiment Station Bulletin No. 245; University of Illinois: Urbana, IL, USA, 1923; Available online: http://hdl.handle.net/2142/3304 (accessed on 24 January 2026).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.S. Physical constraints on voluntary intake of forages by ruminants. J. Anim. Sci. 1996, 74, 3063–3075. [Google Scholar] [CrossRef] [PubMed]
- Fessenden, S.W.; Ross, D.A.; Block, E.; Van Amburgh, M.E. Comparison of milk production, intake, and total-tract nutrient digestion in lactating dairy cattle fed diets containing either wheat middlings and urea, commercial fermentation by-product, or rumen-protected soybean meal. J. Dairy Sci. 2020, 103, 5090–5101. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.J. Limiting factors for milk production in dairy cows: Perspectives from physiology and nutrition. J. Anim. Sci. 2022, 100, skac044. [Google Scholar] [CrossRef]
- Zhao, X.; Zang, C.; Zhao, S.; Zheng, N.; Zhang, Y.; Wang, J. Assessing milk urea nitrogen as an indicator of protein nutrition and nitrogen utilization efficiency: A meta-analysis. J. Dairy Sci. 2025, 108, 4851–4862. [Google Scholar] [CrossRef]
- Stocco, G.; Summer, A.; Cipolat-Gotet, C.; Zanini, L.; Vairani, D.; Dadousis, C.; Zecconi, A. Differential somatic cell count as a novel indicator of milk quality in dairy cows. Animals 2020, 10, 753. [Google Scholar] [CrossRef]
- National Mastitis Council. Guidelines on Normal and Abnormal Raw Milk Based on Somatic Cell Counts and Signs of Clinical Mastitis; National Mastitis Council: Verona, WI, USA, 2001. [Google Scholar]
- Wang, D.; Tang, G.; Zhao, L.; Wang, M.; Chen, L.; Zhao, C.; Liang, Z.; Chen, J.; Cao, Y.; Yao, J. Potential roles of the rectum keystone microbiota in modulating the microbial community and growth performance in goat model. J. Anim. Sci. Biotechnol. 2023, 14, 55. [Google Scholar] [CrossRef]
- Petri, R.M.; Münnich, M.; Zebeli, Q.; Klevenhusen, F. Graded replacement of corn grain with molassed sugar beet pulp modulates the fecal microbial community and hindgut fermentation profile in lactating dairy cows. J. Dairy Sci. 2019, 102, 5019–5030. [Google Scholar] [CrossRef]
- Lyu, J.; Yang, Z.; Wang, E.; Liu, G.; Wang, Y.; Wang, W.; Li, S. Possibility of using by-products with high NDF content to alter the fecal short chain fatty acid profiles, bacterial community, and digestibility of lactating dairy cows. Microorganisms 2022, 10, 1731. [Google Scholar] [CrossRef]
- Flint, H.J.; Bayer, E.A.; Rincon, M.T.; Lamed, R.; White, B.A. Polysaccharide utilization by gut bacteria: Potential for new insights from genomic analysis. Nat. Rev. Microbiol. 2008, 6, 121–131. [Google Scholar] [CrossRef]
- Gérard, P. Metabolism of cholesterol and bile acids by the gut microbiota. Pathogens 2014, 3, 14–24. [Google Scholar] [CrossRef]
- Gerritsen, J.; Hornung, B.; Renckens, B.; van Hijum, S.A.F.T.; Martins dos Santos, V.A.P.; Rijkers, G.T.; Schaap, P.J.; de Vos, W.M.; Smidt, H. Genomic and functional analysis of Romboutsia ilealis CRIBT reveals adaptation to the small intestine. PeerJ 2017, 5, e3698. [Google Scholar] [CrossRef]
- Bosshard, P.P.; Zbinden, R.; Altwegg, M. Turicibacter sanguinis gen. nov., sp. nov., a novel anaerobic, Gram-positive bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52, 1263–1266. [Google Scholar] [CrossRef]
- Benedet, A.; Manuelian, C.L.; Zidi, A.; Penasa, M.; De Marchi, M. Invited review: β-hydroxybutyrate concentration in blood and milk and its associations with dairy cow health and performance. Animal 2019, 13, 1676–1689. [Google Scholar] [CrossRef]
- Tatone, E.H.; Gordon, J.L.; Hubbs, J.; LeBlanc, S.J.; DeVries, T.J.; Duffield, T.F. A systematic review and meta-analysis of the diagnostic accuracy of point-of-care tests for the detection of hyperketonemia in dairy cows. Prev. Vet. Med. 2016, 130, 18–32. [Google Scholar] [CrossRef]
- Tatone, E.H.; Duffield, T.F.; LeBlanc, S.J.; DeVries, T.J.; Gordon, J.L. Investigating the within-herd prevalence and risk factors for ketosis in dairy cattle in Ontario as diagnosed by the test-day concentration of β-hydroxybutyrate in milk. J. Dairy Sci. 2017, 100, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Rigout, S.; Hurtaud, C.; Lemosquet, S.; Bach, A.; Rulquin, H. Lactational effect of propionic acid and duodenal glucose in cows. J. Dairy Sci. 2003, 86, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB Life 2010, 62, 869–877. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Items 1 | Silage 2 | SEM 3 | p-Value | |
|---|---|---|---|---|
| CS | TS | |||
| DM, % | 29.85 | 27.20 | 0.342 | <0.001 |
| CP, % of DM | 8.08 | 11.22 | 0.314 | <0.001 |
| EE, % of DM | 2.78 | 3.82 | 0.042 | 0.002 |
| NDF, % of DM | 47.00 | 50.35 | 0.423 | 0.032 |
| ADF, % of DM | 31.68 | 33.93 | 0.382 | 0.037 |
| Ash, % of DM | 7.09 | 8.03 | 0.043 | 0.004 |
| Items | CON | TS25 | TS50 |
|---|---|---|---|
| Ingredients, % of DM | |||
| Corn silage | 41.16 | 30.87 | 20.58 |
| Triticale silage | 0.00 | 10.29 | 20.58 |
| Alfalfa silage | 7.89 | 7.89 | 7.89 |
| Alfalfa hay | 7.82 | 7.82 | 7.82 |
| Brewers’ grains | 3.29 | 3.29 | 3.29 |
| Brewers’ yeast | 3.18 | 3.18 | 3.18 |
| Corn | 18.92 | 18.92 | 18.92 |
| Wheat | 4.02 | 4.02 | 4.02 |
| Soybean meal | 5.59 | 5.59 | 5.59 |
| Corn distillers dried grains with solubles | 2.46 | 2.46 | 2.46 |
| Premix 1 | 1.00 | 1.00 | 1.00 |
| Limestone powder | 0.65 | 0.65 | 0.65 |
| Dicalcium phosphate | 0.10 | 0.10 | 0.10 |
| Na2HPO4 | 0.05 | 0.05 | 0.05 |
| NaCl | 0.24 | 0.24 | 0.24 |
| Wheat bran | 2.19 | 2.19 | 2.19 |
| Corn germ meal | 0.30 | 0.30 | 0.30 |
| Beet pulp | 0.26 | 0.26 | 0.26 |
| NaHCO3 | 0.88 | 0.88 | 0.88 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutrient components, % of DM | |||
| Crude protein (CP) | 14.31 | 14.60 | 14.89 |
| Ether extract (EE) | 3.19 | 3.28 | 3.40 |
| Crude ash (Ash) | 8.02 | 8.12 | 8.21 |
| Neutral detergent fiber (NDF) | 35.1 | 35.52 | 35.93 |
| Acid detergent fiber (ADF) | 23.08 | 23.37 | 23.65 |
| Ca | 0.91 | 0.88 | 0.85 |
| Phosphorus | 0.37 | 0.37 | 0.37 |
| Items 1 | Treatment 2 | SEM | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | TS25 | TS50 | Trt | Time | Trt × Time | L | Q | ||
| Milk yield, kg/d | 29.66 | 29.41 | 29.78 | 0.85 | 0.883 | <0.001 | 0.320 | 0.914 | 0.790 |
| 4% FCM, kg/d | 30.26 | 30.01 | 30.73 | 0.87 | 0.582 | <0.001 | 0.020 | 0.688 | 0.683 |
| Items 1 | Treatment 2 | SEM | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | TS25 | TS50 | Trt | Time | Trt × Time | L | Q | ||
| Fat, % | 4.10 | 4.27 | 4.16 | 0.31 | 0.894 | 0.927 | 0.766 | 0.680 | 0.951 |
| Protein, % | 3.35 | 3.44 | 3.35 | 0.10 | 0.157 | 0.027 | 0.249 | 0.235 | 0.064 |
| Tots, % | 13.31 | 12.41 | 13.18 | 0.88 | 0.283 | 0.468 | 0.224 | 0.933 | 0.115 |
| MUN, mg/dL | 11.99 | 12.76 | 11.81 | 1.86 | 0.631 | 0.034 | 0.356 | 0.344 | 0.873 |
| SCC, ×103 cells/mL | 14.15 | 16.89 | 9.86 | 5.59 | 0.077 | 0.562 | 0.232 | 0.105 | 0.108 |
| Lactose, % | 4.92 | 4.98 | 4.91 | 0.11 | 0.849 | NA | NA | 0.920 | 0.577 |
| Acetone, mmol/L | 0.06 | 0.05 | 0.05 | 0.02 | 0.932 | NA | NA | 0.741 | 0.875 |
| BHBA, mmol/L | 0.06 | 0.05 | 0.04 | 0.01 | 0.453 | NA | NA | 0.228 | 0.730 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Wang, E.; Han, X.; Sun, W.; Zheng, C.; Du, W. Replacing up to 50% of Corn Silage with Triticale Silage Alters the Fecal Microbiome but Not Milk Yield or Composition in Mid-Lactation Holstein Cows. Animals 2026, 16, 1122. https://doi.org/10.3390/ani16071122
Wang E, Han X, Sun W, Zheng C, Du W. Replacing up to 50% of Corn Silage with Triticale Silage Alters the Fecal Microbiome but Not Milk Yield or Composition in Mid-Lactation Holstein Cows. Animals. 2026; 16(7):1122. https://doi.org/10.3390/ani16071122
Chicago/Turabian StyleWang, Erlong, Xiaoxia Han, Weidong Sun, Chen Zheng, and Wenhua Du. 2026. "Replacing up to 50% of Corn Silage with Triticale Silage Alters the Fecal Microbiome but Not Milk Yield or Composition in Mid-Lactation Holstein Cows" Animals 16, no. 7: 1122. https://doi.org/10.3390/ani16071122
APA StyleWang, E., Han, X., Sun, W., Zheng, C., & Du, W. (2026). Replacing up to 50% of Corn Silage with Triticale Silage Alters the Fecal Microbiome but Not Milk Yield or Composition in Mid-Lactation Holstein Cows. Animals, 16(7), 1122. https://doi.org/10.3390/ani16071122