
Evaluation of Tricaine (MS-222) and Eugenol for Sedation of Peruvian Grunt Anisotremus scapularis

 ,
,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Conditioning of Juvenile A. scapularis
2.2. Determination of Anesthetic Induction Time
2.3. Determination of Anesthetic Recovery Time
2.4. Data Analysis
3. Results
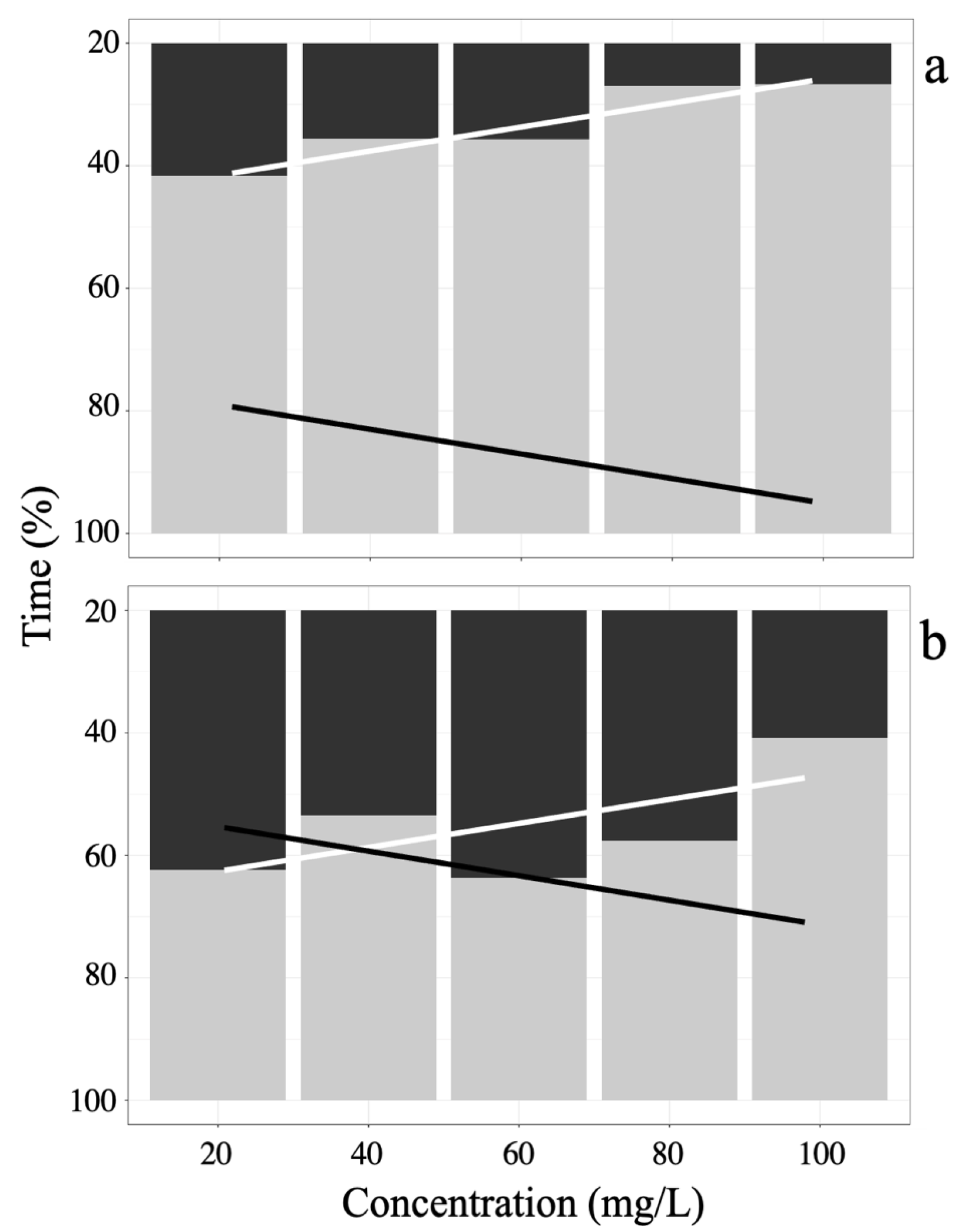
3.1. Eugenol
3.2. Tricaine MS-222
4. Discussion
4.1. Eugenol
4.2. MS-222 (Tricaine)
4.3. Perspectives and Development of Anesthetics in Aquaculture
5. Conclusions
- We determined that an effective concentration of eugenol in the range of 20 to 60 mg/L and 80 mg/L of MS-222 was sufficient to achieve induction and anesthetic recovery times of less than 6 and 16 min, respectively, with no observed mortality.
- In this study, eugenol was a more effective anesthetic than MS-222. This efficacy was defined as a combination of: (a) short induction times, (b) complete recovery with no visible adverse effects, and (c) an adequate duration of deep anesthesia (stage I-3) that allowed handling procedures to be performed with minimal interference from fish movement. In addition, eugenol showed better performance in terms of stability and duration of the deep anesthetic state.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carrera, L.; Cota, N.; Linares, J.; Castro, A.; Orihuela, L.; Silva, E.; Montes, M. Manual Para Acondicionamiento de Reproductores de Chita Anisotremus scapularis. Inf. Inst. Mar Perú 2018, 45, 263–276. [Google Scholar]
- Ramos, L.A.E.; Layme, V.F.C.; Victoriano, R.G.P.; Choqueapaza, J.P.; Mamani, Z.C. Captura, Acondicionamiento y Primer Desove de Sargo Anisotremus scapularis en la Región Tacna. Cienc. Desarro. 2019, 25, 68–74. [Google Scholar] [CrossRef]
- Montes, M.; Castro, A.M.; Linares, J.F.; Orihuela, L.I.; Carrera, L.J. Embryonic Development of Peruvian Grunt Anisotremus scapularis (Perciformes: Haemulidae). Rev. Biol. Mar. Oceanogr. 2019, 54, 166–173. [Google Scholar] [CrossRef]
- Instituto del Mar del Perú: IMARPE. Ciclo de Vida de la Chita Anisotremus scapularis. Ser. Divulg. Científica IMARPE 2015, 1, 1–20. [Google Scholar]
- Chirichigno, N. Clave para Identificar los Peces Marinos del Perú; FAO: Lima, Peru, 1974; Volume 44. [Google Scholar]
- Froese, R.; Pauly, D. Species List: World Wide Web Electronic Publication. Available online: www.fishbase.org (accessed on 17 January 2025).
- IMARPE. Evaluación Pesquera y Perspectiva de Manejo de Chita Anisotremus scapularis en el Litoral Peruano; IMARPE: Lima, Peru, 2018. [Google Scholar]
- Pepe-Victoriano, R.; Huanacuni, J.I.; Presa, P.; Espinoza-Ramos, L.A. Reproductive Management: Conditioning, Spawning and Development of Peruvian Grunt Anisotremus scapularis in Southern Peru. PeerJ 2025, 13, e18655. [Google Scholar] [CrossRef]
- He, R.; Lei, B.; Su, Y.; Wang, A.; Cui, K.; Shi, X.; Chen, X. Effectiveness of Eugenol as an Anesthetic for Adult Spotted Sea Bass (Lateolabrax maculatus). Aquaculture 2020, 523, 735180. [Google Scholar] [CrossRef]
- Pepe-Victoriano, R.; Pepe-Vargas, P.; Yañez-Valenzuela, M.; Aravena-Ambrosetti, H.; Olivares-Cantillano, G.; Méndez-Abarca, F.; Huanacuni, J.I.; Méndez, S.; Espinoza-Ramos, L. Growth of Oncorhynchus mykiss (Rainbow Trout) through a Recirculation System in the Foothills of the Extreme North of Chile. Animals 2024, 14, 2567. [Google Scholar] [CrossRef]
- Pepe-Victoriano, R.; Aravena-Ambrosetti, H.; Merino, G.E. Breeding of a Wild Population of South Pacific Bonito Sarda Chiliensis Chiliensis (Cuvier 1832) Broodstock under Laboratory Conditions in Pisagua, Northern Chile. Animals 2021, 12, 24. [Google Scholar] [CrossRef]
- Bell, T.G.; Johnson, M.T.; Jickells, T.D.; Liss, P.S. Ammonia/Ammonium Dissociation Coefficient in Seawater: A Significant Numerical Correction. Environ. Chem. 2007, 4, 183–186, Erratum in Environ. Chem. 2008, 5, 258. [Google Scholar] [CrossRef]
- Kwasek, K.; Rimoldi, S.; Cattaneo, A.G.; Parker, T.; Dabrowski, K.; Terova, G. The Expression of Hypoxia-Inducible Factor-1α Gene Is Not Affected by Low-Oxygen Conditions in Yellow Perch (Perca flavescens) Juveniles. Fish Physiol. Biochem. 2017, 43, 849–862. [Google Scholar] [CrossRef]
- Mancera, J.M.; Martínez-Rodríguez, G.; Skrzynska, A.K.; Martos-Sitcha, J.A. Osmoregulatory Role of Vasotocinergic and Isotocinergic Systems in the Gilthead Sea Bream (Sparus aurata L). Gen. Comp. Endocrinol. 2018, 257, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Chacoff, L.; Ruiz-Jarabo, I.; Páscoa, I.; Gonçalves, O.; Mancera, J.M. Yearly Growth and Metabolic Changes in Earthen Pond-Cultured Meagre Argyrosomus regius. Sci. Mar. 2014, 78, 193–202. [Google Scholar] [CrossRef]
- Tacchi, L.; Lowrey, L.; Musharrafieh, R.; Crossey, K.; Larragoite, E.T.; Salinas, I. Effects of Transportation Stress and Addition of Salt to Transport Water on the Skin Mucosal Homeostasis of Rainbow Trout (Oncorhynchus mykiss). Aquaculture 2015, 435, 120–127. [Google Scholar] [CrossRef]
- Priborsky, J.; Velisek, J. A Review of Three Commonly Used Fish Anesthetics. Rev. Fish. Sci. Aquac. 2018, 26, 417–442. [Google Scholar] [CrossRef]
- Altun, T.; Bilgin, R.; Danabaş, D. Efectos del Bicarbonato de Sodio en la Anestesia de los Juveniles de Carpa Común (Cyprinus Carpio L., 1758). Rev. Turca Pesca Cienc. Acuáticas 2009, 9, 29–31. [Google Scholar]
- Hajek, G.J.; Kłyszejko, B.; Dziaman, R. The Anaesthetic Effect of Clove Oil on Common Carp, Cyprinus carpio L. Acta Ichthyol. Piscat. 2006, 36, 93–97. [Google Scholar] [CrossRef]
- Sneddon, L.U. Clinical Anesthesia and Analgesia in Fish. J. Exot. Pet Med. 2012, 21, 32–43. [Google Scholar] [CrossRef]
- Opiyo, M.A.; Ogello, E.O.; Charo-Karisa, H. Effectiveness of Sodium Bicarbonate as an Anaesthetic for Different Sizes of Nile Tilapia (Oreochromis niloticus L., 1758) Juveniles. Int. J. Aquat. Sci. 2013, 4, 14–22. [Google Scholar]
- Githuka, C.M. Anaesthetic Effects of Sodium Bicarbonate at Different Concentrations on African Catfish (Clarias gariepinus) Juveniles. J. Aquac. Eng. Fish. Res. 2016, 2, 151–158. [Google Scholar] [CrossRef]
- Hasimuna, O.J.; Maulu, S.; Monde, C.; Mweemba, M. Cage Aquaculture Production in Zambia: Assessment of Opportunities and Challenges on Lake Kariba, Siavonga District. Egypt. J. Aquat. Res. 2019, 45, 281–285. [Google Scholar] [CrossRef]
- Ross, L.G.; Ross, B. Anaesthetic and Sedative Techniques for Aquatic Animals, 3rd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2008; ISBN 978-1-405-14938-9. [Google Scholar]
- Nascimento, H.d.S.; Crispim, B.D.A.; Francisco, L.F.V.; Merey, F.M.; Kummrow, F.; Viana, L.F.; Inoue, L.A.K.A.; Barufatti, A. Genotoxicity Evaluation of Three Anesthetics Commonly Employed in Aquaculture Using Oreochromis niloticus and Astyanax lacustris. Aquac. Rep. 2020, 17, 100357. [Google Scholar] [CrossRef]
- Flores, C. Respuestas Neuroendócrinas al Estrés En Peces Teleósteos. Rev. Ictiol. 2002, 10, 57–78. [Google Scholar]
- Carter, K. The Effects of Dissolved Oxygen on Steelhead Trout, Coho Salmon, and Chinook Salmon Biology and Function by Life Stage. Quality 2005, 1–10. Available online: https://www.noaa.gov/sites/default/files/legacy/document/2020/Oct/0.7.115.29756-000004.pdf (accessed on 20 April 2025).
- Moraes, G.; Afonso, L.B.; Barton, B.A.; Iwama, G.K.; Mackinlay, D. Stress in Fish: Consequences to Performance. Symposium Proceedings; Physiology Section, Ed.; American Fisheries Society: Bethesda, MD, USA, 2004. [Google Scholar]
- Velisek, J.; Svobodova, Z.; Piackova, V.; Groch, L.; Nepejchalova, L. Effects of Clove Oil Anaesthesia on Common Carp (Cyprinus Carpio L.). Veter Med. 2005, 50, 269–275. [Google Scholar] [CrossRef]
- da Cunha, M.A.; Zeppenfeld, C.C.; Garcia, L.d.O.; Loro, V.L.; da Fonseca, M.B.; Emanuelli, T.; Veeck, A.P.d.L.; Copatti, C.E.; Baldisserotto, B. Anesthesia of Silver Catfish with Eugenol: Time of Induction, Cortisol Response and Sensory Analysis of Fillet. Cienc. Rural. 2010, 40, 2107–2114. [Google Scholar] [CrossRef]
- McFarland, W.N. A Study of the Effects of Anaesthetics on the Behaviour and Physiology of Fishes. Publ. Inst. Mar. Sci. 1959, 6, 22–55. [Google Scholar]
- Hikasa, Y.; Takase, K.; Ogasawara, T.; Ogasawara, S. Anesthesia and Recovery with Tricaine Methanesulfonate, Eugenol and Thiopental Sodium in the Carp, Cyprinus carpio. Jpn. J. Vet. Sci. 1986, 48, 341–351. [Google Scholar] [CrossRef]
- Iwama, G.K.; Thomas, P.T.; Forsyth, R.B.; Vijayan, M.M. Heat Shock Protein Expression in Fish. Rev. Fish Biol. Fish. 1998, 8, 35–56. [Google Scholar] [CrossRef]
- Cooke, S.J.; Suski, C.D.; Ostrand, K.G.; Tufts, B.L.; Wahl, D.H. Behavioral and Physiological Assessment of Low Concentrations of Clove Oil Anaesthetic for Handling and Transporting Largemouth Bass (Micropterus salmoides). Aquaculture 2004, 239, 509–529. [Google Scholar] [CrossRef]
- Park, I.-S. The Anesthetic Effects of Clove Oil and MS-222 on Far Eastern Catfish, Silurus asotus. Dev. Reprod. 2019, 23, 183–191. [Google Scholar] [CrossRef]
- Wang, W.; Dong, H.; Sun, Y.; Cao, M.; Duan, Y.; Li, H.; Liu, Q.; Gu, Q.; Zhang, J. The Efficacy of Eugenol and Tricaine Methanesulphonate as Anaesthetics for Juvenile Chinese Sea Bass (Lateolabrax maculatus) during Simulated Transport. J. Appl. Ichthyol. 2019, 35, 551–557. [Google Scholar] [CrossRef]
- Lee, S.; Nguyen, P.T.; Song, H.-K.; Hur, S.P.; Kim, J.-H. Effect of Water Temperature, Fish Age, and MS-222 Concentration on the Anesthetization of River Pufferfish, Takifugu obscurus. Korean J. Ichthyol. 2023, 35, 67–74. [Google Scholar] [CrossRef]
- Cao, J.; Wang, Q.; Qiu, W.; Mei, J.; Xie, J. Transport and Recovery of Turbot (Scophthalmus Maximus) Sedated with MS-222 and Eugenol: Effects on Intermediary Metabolism and Osmoregulation. Animals 2021, 11, 2228. [Google Scholar] [CrossRef]
- Teles, M.; Oliveira, M.; Jerez-Cepa, I.; Franco-Martínez, L.; Tvarijonaviciute, A.; Tort, L.; Mancera, J.M. Transport and Recovery of Gilthead Sea Bream (Sparus aurata L.) Sedated With Clove Oil and MS222: Effects on Oxidative Stress Status. Front. Physiol. 2019, 10, 523. [Google Scholar] [CrossRef]
- Villamizar, N.; De Luque, A.; Gaitán-Ibarra, S. Evaluation of Eugenol as a Sedative for the Transportation of Common Snook Centropomus undecimalis (Bloch, 1792). Aquac. Res. 2021, 52, 5898–5902. [Google Scholar] [CrossRef]
- Honorato, L.; Batista, J.V.d.S.; Durigon, E.G.; de Mello, G.L.; Medeiros, M.V. Eugenol and Tricaine Methanesulfonate as Anhestesics for the Pearl Cichlid. Acta Sci. Biol. Sci. 2021, 43, e53422. [Google Scholar] [CrossRef]
- Nuanmanee, S.; Sriwanayos, P.; Boonyo, K.; Chaisri, W.; Saengsitthisak, B.; Tajai, P.; Pikulkaew, S. Synergistic Effect between Eugenol and 1,8-Cineole on Anesthesia in Guppy Fish (Poecilia reticulata). Veter Sci. 2024, 11, 165. [Google Scholar] [CrossRef]
- Vidal, L.V.O.; Albinati, R.C.B.; Albinati, A.C.L.; de Lira, A.D.; de Almeida, T.R.; Santos, G.B. Eugenol Como Anestésico Para a Tilápia-Do-Nilo. Pesqui. Agropecu. Bras. 2008, 43, 1069–1074. [Google Scholar] [CrossRef]
- Vidal, L.V.O.; Albinati, R.C.B.; Albinati, A.C.L.; de Mecêdo, G.R. Utilização Do Eugenol Como Anestésico Para o Manejo de Juvenis de Pintado (Pseudoplatystoma corruscans). Acta Sci. Biol. Sci. 2007, 28, 275–279. [Google Scholar] [CrossRef]
- Jia, Y.; Xie, T.; Gao, Y.; Qin, H.; Guan, C. Anesthetics Efficacy and Physiological Response of MS222 and Clove Oil in Spotted Knifejaw Oplegnathus punctatus. Aquac. Rep. 2022, 25, 101201. [Google Scholar] [CrossRef]
- Gomes, D.P.; Chaves, B.W.; Becker, A.G.; Baldisserotto, B. Water Parameters Affect Anaesthesia Induced by Eugenol in Silver Catfish, Rhamdia quelen. Aquac. Res. 2011, 42, 878–886. [Google Scholar] [CrossRef]
- Meinertz, J.R.; Schreier, T.M.; Porcher, S.T.; Smerud, J.R.; Gaikowski, M.P. Depletion of Eugenol Residues from the Skin-on Fillet Tissue of Rainbow Trout Exposed to 14C-Labeled Eugenol. Aquaculture 2014, 430, 74–78. [Google Scholar] [CrossRef]
- Cupp, A.R.; Hartleb, C.F.; Fredricks, K.T.; Gaikowski, M.P. Effectiveness of Eugenol Sedation to Reduce the Metabolic Rates of Cool and Warm Water Fish at High Loading Densities. Aquac. Res. 2016, 47, 234–242. [Google Scholar] [CrossRef]
- Cupp, A.R.; Schreier, T.M.; Schleis, S.M. Live Transport of Yellow Perch and Nile Tilapia in AQUI-S 20E (10% Eugenol) at High Loading Densities. N. Am. J. Aquac. 2017, 79, 176–182. [Google Scholar] [CrossRef]
- Cunha, F.E.A.; Rosa, I.L. Anaesthetic Effects of Clove Oil on Seven Species of Tropical Reef Teleosts. J. Fish Biol. 2006, 69, 1504–1512. [Google Scholar] [CrossRef]
- Roohi, Z.; Imanpoor, M.R. The Efficacy of the Oils of Spearmint and Methyl Salicylate as New Anesthetics and Their Effect on Glucose Levels in Common Carp (Cyprinus carpio L., 1758) Juveniles. Aquaculture 2015, 437, 327–332. [Google Scholar] [CrossRef]
- Woody, C.A.; Nelson, J.; Ramstad, K. Clove Oil as an Anaesthetic for Adult Sockeye Salmon: Field Trials. J. Fish Biol. 2002, 60, 340–347. [Google Scholar] [CrossRef]
- Purbosari, N.; Warsiki, E.; Syamsu, K.; Santoso, J. Natural versus Synthetic Anesthetic for Transport of Live Fish: A Review. Aquac. Fish. 2019, 4, 129–133. [Google Scholar] [CrossRef]
- Uehara, S.; Andrade, D.; Takata, R.; Júnior, A.G.; Vidal, M. The Effectiveness of Tricaine, Benzocaine, Clove Oil, and Menthol as Anesthetics for Lambari-Bocarra Oligosarcus argenteus. Aquaculture 2019, 502, 326–331. [Google Scholar] [CrossRef]
- Weber, R.; Peleteiro, J.; Martín, L.G.; Aldegunde, M. The Efficacy of 2-Phenoxyethanol, Metomidate, Clove Oil and MS-222 as Anaesthetic Agents in the Senegalese Sole (Solea senegalensis Kaup 1858). Aquaculture 2009, 288, 147–150. [Google Scholar] [CrossRef]
- Jerez-Cepa, I.; Fernández-Castro, M.; O’Neill, T.J.D.S.; Martos-Sitcha, J.A.; Martínez-Rodríguez, G.; Mancera, J.M.; Ruiz-Jarabo, I. Transport and Recovery of Gilthead Seabream (Sparus aurata L.) Sedated with Clove Oil and MS-222: Effects on Stress Axis Regulation and Intermediary Metabolism. Front. Physiol. 2019, 10, 612. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.M.A.; Wada, S.; Hatai, K.; Yamamoto, A. Antimycotic Activity of Eugenol against Selected Water Molds. J. Aquat. Anim. Health 2000, 12, 224–229. [Google Scholar] [CrossRef]
- Parker-Graham, C.A.; Lima, K.M.; Soto, E. The Effect of Anesthetic Time and Concentration on Blood Gases, Acid-Base Status, and Electrolytes in Koi (Cyprinus Carpio) Anesthetized with Buffered Tricaine Methanesulfonate (MS-222). J. Zoo Wildl. Med. 2020, 51, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Nochetto, C.B.; Reimschuessel, R.; Gieseker, C.; Cheely, C.S.; Carson, M.C. Determination of Tricaine Residues in Fish by Liquid Chromatography. J. AOAC Int. 2009, 92, 1241–1248. [Google Scholar] [CrossRef]
- Rosado, M.; Dionicio, J.; Aguirre-Velarde, A. Evaluación de Diferentes Concentraciones de Tricaína (MS-222) en el Transporte de Chitas (Anisotremus scapularis) Juveniles. Rev. Investig. Vet. Perú 2016, 27, 687. [Google Scholar] [CrossRef]
- Iwama, G.K.; Ackerman, P.A. Anaesthetics. In Biochemistry and Molecular Biology of Fishes; Elsevier: Amsterdam, The Netherlands, 1994; Volume 3, pp. 1–15. [Google Scholar]
- Kreiberg, H. Stress and Anesthesia. In The Laboratory Fish; Elsevier: Amsterdam, The Netherlands, 2000; pp. 503–511. [Google Scholar]
- Neiffer, D.L.; Stamper, M.A. Fish Sedation, Anesthesia, Analgesia, and Euthanasia: Considerations, Methods, and Types of Drugs. ILAR J. 2009, 50, 343–360. [Google Scholar] [CrossRef]
- Carter, K.M.; Woodley, C.M.; Brown, R.S. A Review of Tricaine Methanesulfonate for Anesthesia of Fish. Rev. Fish Biol. Fish. 2011, 21, 51–59. [Google Scholar] [CrossRef]
- Pelkowski, S.D.; Kapoor, M.; Richendrfer, H.A.; Wang, X.; Colwill, R.M.; Creton, R. A Novel High-Throughput Imaging System for Automated Analyses of Avoidance Behavior in Zebrafish Larvae. Behav. Brain Res. 2011, 223, 135–144. [Google Scholar] [CrossRef]
- Readman, G.D.; Owen, S.F.; Murrell, J.C.; Knowles, T.G. Do Fish Perceive Anaesthetics as Aversive? PLoS ONE 2013, 8, e73773. [Google Scholar] [CrossRef]
- Roubach, R.; Gomes, L.d.C.; Val, A.L. Safest Level of Tricaine Methanesulfonate (Ms-222) to Induce Anesthesia in Juveniles of Matrinxã, Brycon cephalus. Acta Amaz. 2001, 31, 159–163. [Google Scholar] [CrossRef]
- Mitjana, O.; Bonastre, C.; Insua, D.; Falceto, M.V.; Esteban, J.; Josa, A.; Espinosa, E. The Efficacy and Effect of Repeated Exposure to 2-Phenoxyethanol, Clove Oil and Tricaine Methanesulphonate as Anesthetic Agents on Juvenile Angelfish (Pterophyllum scalare). Aquaculture 2014, 433, 491–495. [Google Scholar] [CrossRef]
- Brown, L. Anaesthesia in fishes. J. Small Anim. Pract. 1981, 22, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Durhack, T.C.; Jeffrey, J.D.; Enders, E.C. In Search of an Anaesthesia Alternative for Field-Based Research. Aquaculture 2020, 525, 735285. [Google Scholar] [CrossRef]
- Minaz, M.; Er, A.; Ak, K.; Kurtoğlu, İ.Z.; Kayış, Ş. Determining the Appropriate Concentration of an Anesthetic Mixture in Three Different Fish Species with the PROMETHEE Decision Model. Front. Veter Sci. 2024, 11, 1492769. [Google Scholar] [CrossRef]
- Cho, S.; Yang, H.; Jeon, Y.-J.; Lee, C.J.; Jin, Y.-H.; Baek, N.-I.; Kim, D.; Kang, S.-M.; Yoon, M.; Yong, H.; et al. Phlorotannins of the Edible Brown Seaweed Ecklonia cava Kjellman Induce Sleep via Positive Allosteric Modulation of Gamma-Aminobutyric Acid Type A–Benzodiazepine Receptor: A Novel Neurological Activity of Seaweed Polyphenols. Food Chem. 2012, 132, 1133–1142. [Google Scholar] [CrossRef]
- Bushra, R.; Rahila, N.; Iqbal, A.; Somia, G. Neuropharmacological Screening of Iyengaria Stellata Revealed Its Memory Boosting, Anxiolytic and Antidepressant Effects. IRJP 2012, 10, 3. [Google Scholar]
- Charlie-Silva, I.; Feitosa, N.M.; Gomes, J.M.M.; Hoyos, D.C.d.M.; Mattioli, C.C.; Eto, S.F.; Fernandes, D.C.; Belo, M.A.d.A.; Silva, J.d.O.; de Barros, A.L.B.; et al. Potential of Mucoadhesive Nanocapsules in Drug Release and Toxicology in Zebrafish. PLoS ONE 2020, 15, e0238823. [Google Scholar] [CrossRef]
- Bertotto, D.; Poltronieri, C.; Negrato, E.; Richard, J.; Pascoli, F.; Simontacchi, C.; Radaelli, G. Whole Body Cortisol and Expression of HSP70, IGF-I and MSTN in Early Development of Sea Bass Subjected to Heat Shock. Gen. Comp. Endocrinol. 2011, 174, 44–50. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Anesthesia | Stage Code | Description |
|---|---|---|
| Induction | I-1 | Loss of swimming axis |
| Erratic swimming | ||
| Accelerated opercular frequency | ||
| I-2 | Loss of body movement | |
| Decreased opercular frequency | ||
| Regular opercular rhythm | ||
| Absence of response to external mechanical stimulus | ||
| Absence of reflex response | ||
| I-3 | Body immobility | |
| Severe decrease in opercular frequency | ||
| Irregular opercular rhythm | ||
| Recovery | R-1 | Body immobility |
| Increased opercular frequency | ||
| Regular opercular rhythm | ||
| Absence of response to external mechanical stimulus | ||
| R-2 | Onset of body movement | |
| Response to external mechanical stimulus | ||
| R-3 | Recovery of swimming axis | |
| Behavior like that of pre-anesthesia |
| Eugenol Concentrations (mg/L) | Correlation | ||||||
|---|---|---|---|---|---|---|---|
| Stage | 20 | 40 | 60 | 80 | 100 | p-Value | r |
| I-1 | 52.00 ± 3.61 a | 42.67 ± 5.03 b | 40.33 ± 5.13 b | 20.33 ± 3.51 c | 15.67 ± 2.08 c | <0.001 | −0.942 |
| I-2 | 82.33 ± 19.40 a | 71.33 ± 6.66 ab | 86.33 ± 10.69 a | 31.33 ± 2.52 b | 29.00 ± 4.36 b | 0.003 | −0.715 |
| I-3 | 103.33 ± 20.43 a | 94.33 ± 20.60 ab | 100.67 ± 9.29 ab | 44.33 ± 8.39 bc | 35.00 ± 3.61 c | <0.001 | −0.840 |
| I-Total | 237.67 ± 43.36 a | 208.33 ± 18.77 ab | 227.33 ± 23.12 a | 96 ± 3.46 bc | 79.67 ± 6.66 c | <0.001 | −0.859 |
| R-1 | 79.33 ± 17.01 c | 104.00 ± 10.58 bc | 101.33 ± 4.16 bc | 147.00 ± 4.00 a | 114.67 ± 2.08 b | 0.005 | 0.688 |
| R-2 | 170.00 ± 13.23 c | 229.00 ± 9.00 ab | 232.67 ± 4.62 ab | 258.00 ± 19.08 a | 218.33 ± 10.60 bc | 0.159 | 0.383 |
| R-3 | 390.67 ± 9.02 c | 525.67 ± 4.04 bc | 600.00 ± 9.17 a | 597.67 ± 2.52 a | 548.33 ± 9.07 b | 0.010 | 0.639 |
| R-Total | 640 ± 38.69 d | 858.67 ± 23.35 c | 934.00 ± 12.17 b | 1002.67 ± 15.70 a | 881.33 ± 6.11 c | 0.187 | −0.360 |
| MS-222 Concentrations (mg/L) | Correlation | ||||||
|---|---|---|---|---|---|---|---|
| Stage | 20 | 40 | 60 | 80 | 100 | p-Value | r |
| I-1 | 300.33 ± 13.65 a | 186.67 ± 9.29 b | 50.33 ± 3.79 c | 41.67 ± 3.79 d | 24.33 ± 3.21 e | <0.001 | −0.977 |
| I-2 | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 98.00 ± 17.09 a | 110.67 ± 7.37 a | 53.00 ± 1.00 b | 0.011 | 0.637 |
| I-3 | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 204.00 ± 9.64 a | 139.33 ± 11.37 b | 72.67 ± 3.06 c | 0.031 | 0.558 |
| I-Total | 300.33 ± 13.65 ab | 186.67 ± 9.29 bc | 352.33 ± 25.42 a | 291.67 ± 21.96 ab | 150.00 ± 6.56 c | 0.003 | 0.718 |
| R-1 | 21.00 ± 2.65 a | 19.33 ± 5.13 a | 21.33 ± 17.93 a | 19.33 ± 4.04 a | 26.33 ± 3.21 a | 0.477 | 0.199 |
| R-2 | 53.33 ± 1.53 bc | 51.33 ± 3.21 c | 51.33 ± 5.69 b c | 74.67 ± 5.03 ab | 114.67 ± 0.08 a | 0.001 | 0.771 |
| R-3 | 192.00 ± 3.00 c | 188.33 ± 7.64 c | 220.67 ± 15.50 b | 234.33 ± 5.13 ab | 284.33 ± 13.65 a | <0.001 | 0.912 |
| R-Total | 266.33 ± 5.69 bc | 259.00 ± 15.10 c | 293.33 ± 30.24 ab | 328.33 ± 14.05 ab | 425 ± 11.72 a | <0.001 | 0.881 |
| Specie | Habitat | Eugenol (mg/L) | MS-222 (mg/L) | Source |
|---|---|---|---|---|
| Silurus asotus (far eastern catfish) | Freshwater | 500 | 600 | [35] |
| Lateolabrax maculatus (Chinese sea bass) | Marine | 6 | 30 | [36] |
| Takifugu obscurus (river pufferfish) | Freshwater | – | 150–175 | [37] |
| Scophthalmus maximus (turbot) | Marine | 60 | [38] | |
| Sparus aurata (gilthead sea bream) | Marine | 1 | 60 | [39] |
| Centropomus undecimalis (common snook) | Marine | 50 | – | [40] |
| Geophagus brasiliensis (pearl cichlid) | Freshwater | 217 | 294 | [41] |
| Poecilia reticulata (guppy) | Freshwater | 50–75 | – | [42] |
| Oreochromis niloticus (Nile tilapia) | Freshwater | 90 | 300 | [43] |
| Pseudoplatystoma corruscans (spotted sorubim) | Freshwater | 75 | – | [44] |
| Oplegnathus punctatus (spotted knifejaw) | Marine | 35 | 80 | [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinoza-Ramos, L.A.; Puma-Vilca, Y.; Huanacuni, J.I.; Pepe-Victoriano, R. Evaluation of Tricaine (MS-222) and Eugenol for Sedation of Peruvian Grunt Anisotremus scapularis. Animals 2025, 15, 1322. https://doi.org/10.3390/ani15091322
Espinoza-Ramos LA, Puma-Vilca Y, Huanacuni JI, Pepe-Victoriano R. Evaluation of Tricaine (MS-222) and Eugenol for Sedation of Peruvian Grunt Anisotremus scapularis. Animals. 2025; 15(9):1322. https://doi.org/10.3390/ani15091322
Chicago/Turabian StyleEspinoza-Ramos, Luis Antonio, Ydelsa Puma-Vilca, Jordan I. Huanacuni, and Renzo Pepe-Victoriano. 2025. "Evaluation of Tricaine (MS-222) and Eugenol for Sedation of Peruvian Grunt Anisotremus scapularis" Animals 15, no. 9: 1322. https://doi.org/10.3390/ani15091322
APA StyleEspinoza-Ramos, L. A., Puma-Vilca, Y., Huanacuni, J. I., & Pepe-Victoriano, R. (2025). Evaluation of Tricaine (MS-222) and Eugenol for Sedation of Peruvian Grunt Anisotremus scapularis. Animals, 15(9), 1322. https://doi.org/10.3390/ani15091322