
The Novel Antigenic Epitopes of African Swine Fever Virus Inner Membrane p54 Protein Revealed by Monoclonal Antibodies

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice, Cells, Sera, and Viruses
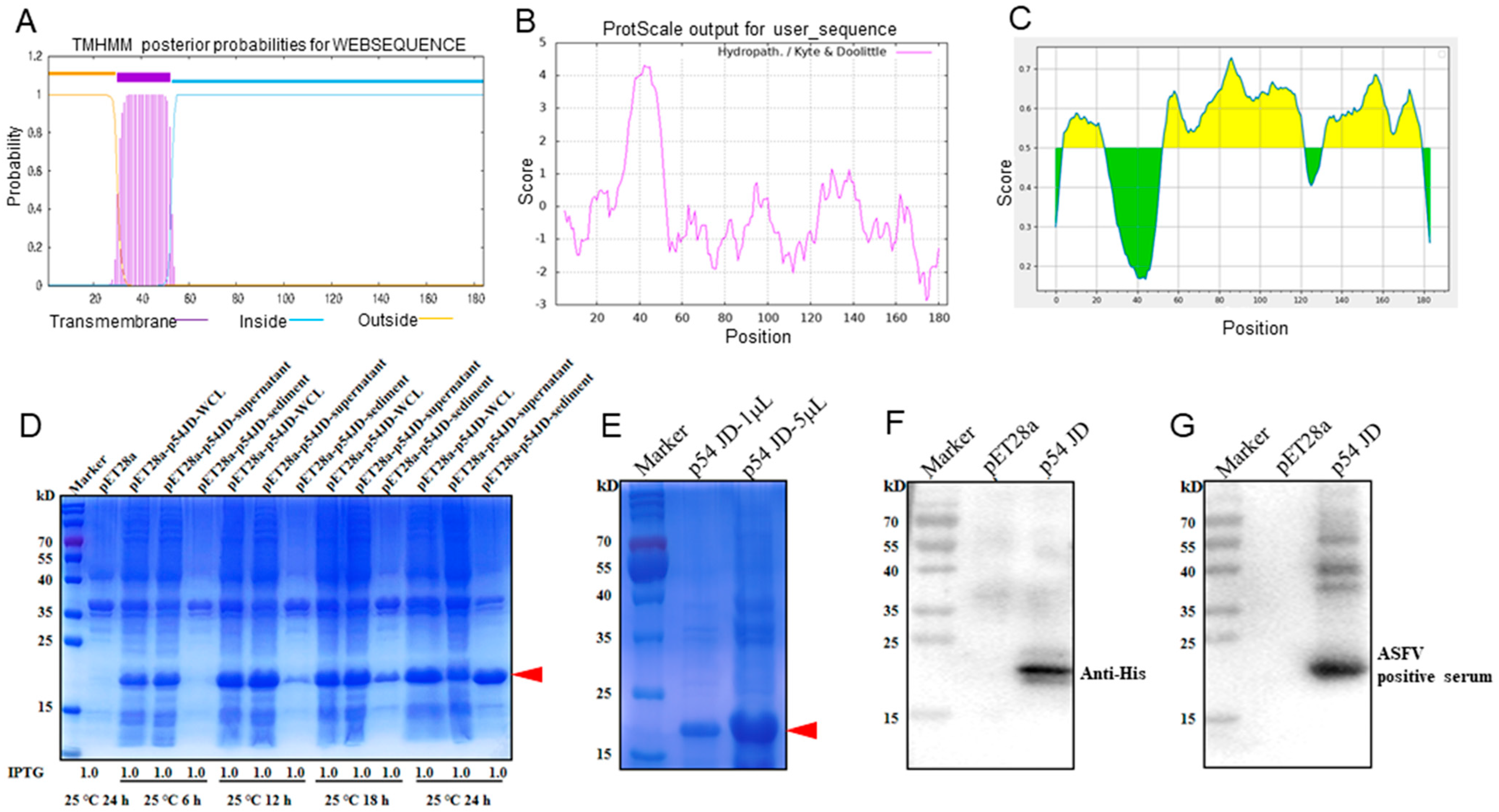
2.2. Expression and Purification of p54 Truncation Protein
2.3. Generation of Anti-p54 Monoclonal Antibodies (mAbs)
2.4. Mapping of the Antigenic Epitopes of the p54 Protein
2.5. Western Blotting (WB) and Immunofluorescence Assay (IFA)
2.6. ASFV p54 Protein Indirect ELISA and Antigenic Peptide Indirect ELISA
2.7. Bioinformatics Analysis
3. Results
3.1. Production and Identification of Recombinant p54 Truncation Protein
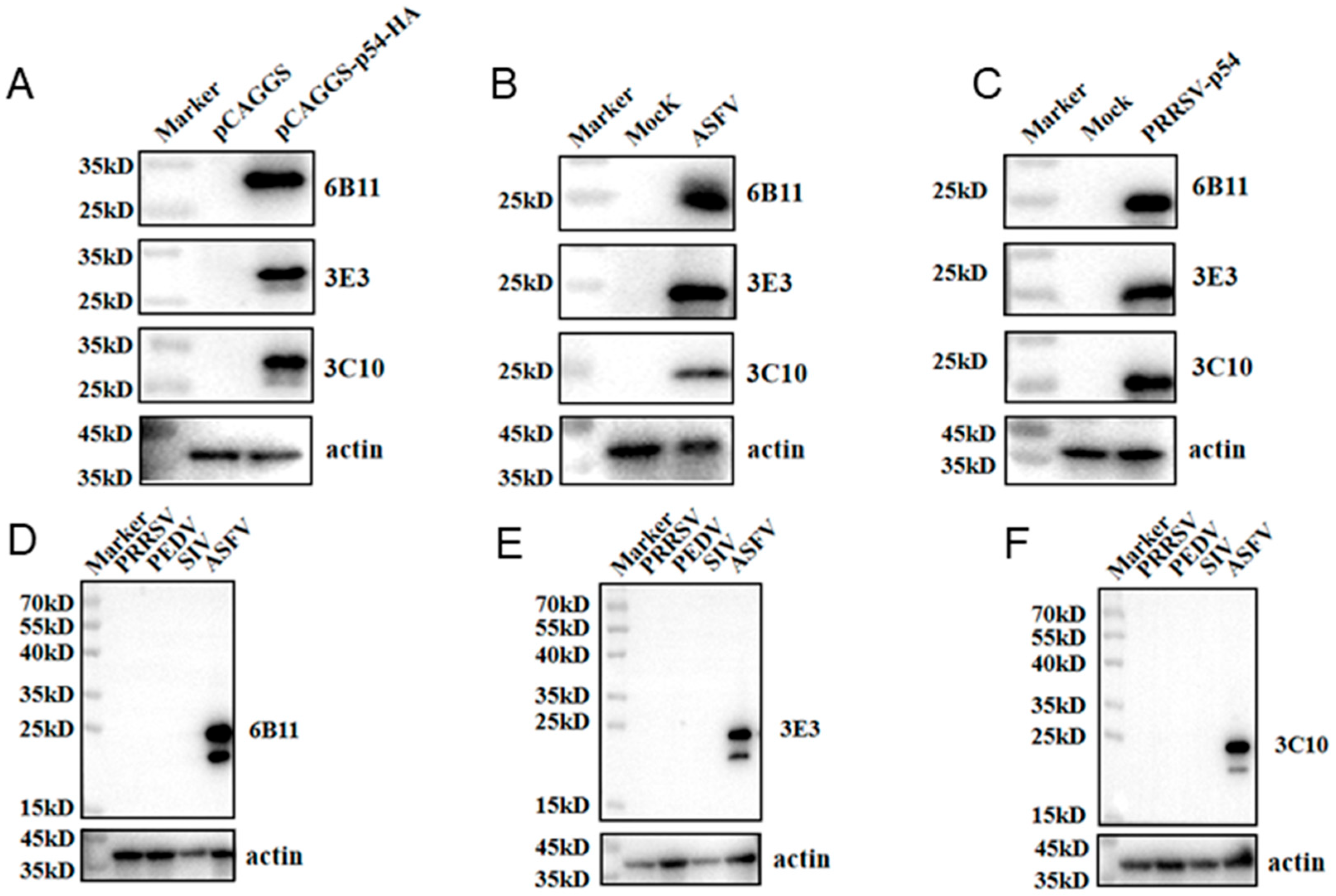
3.2. Generation of Specific Monoclonal Antibodies (mAbs) for p54 Protein
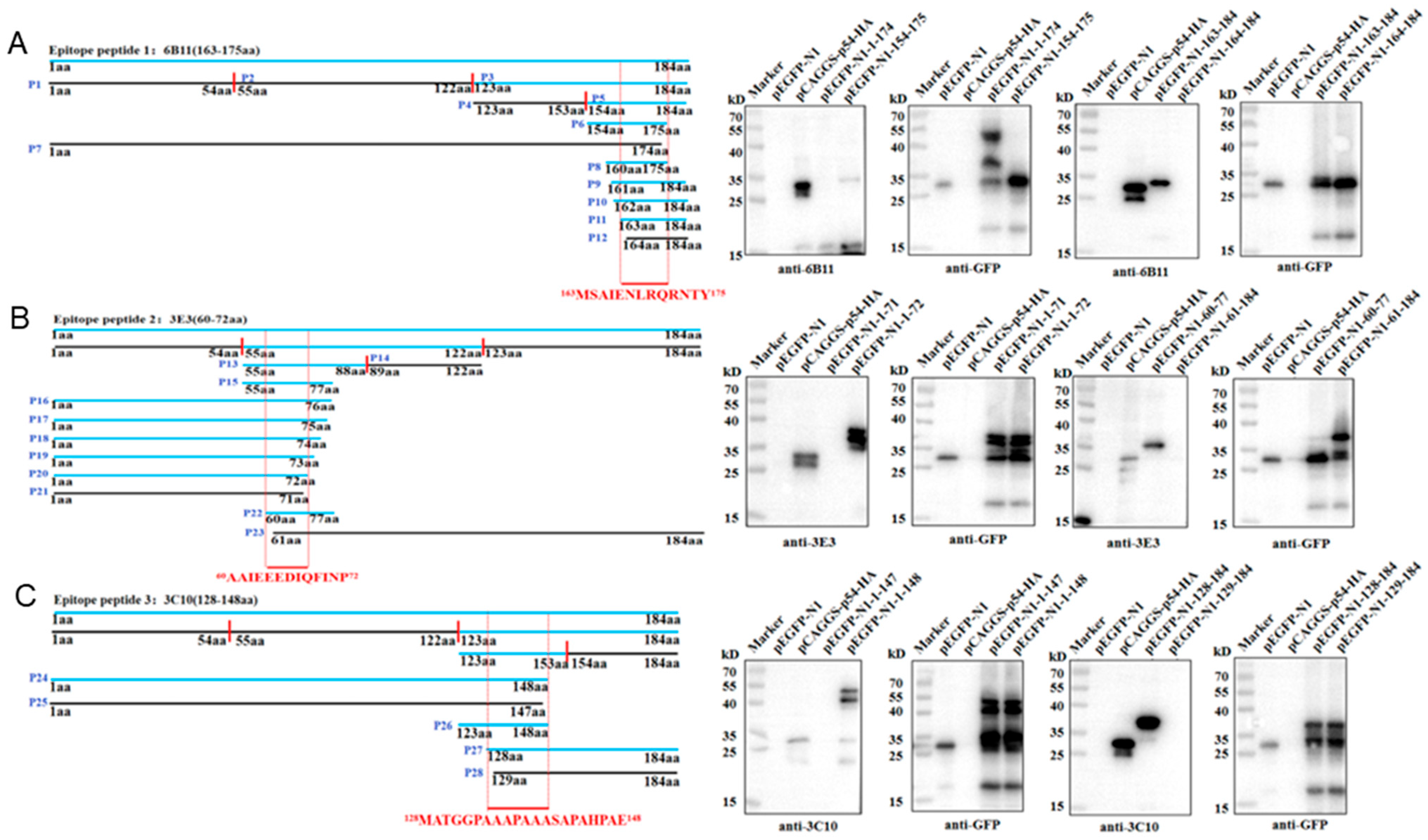
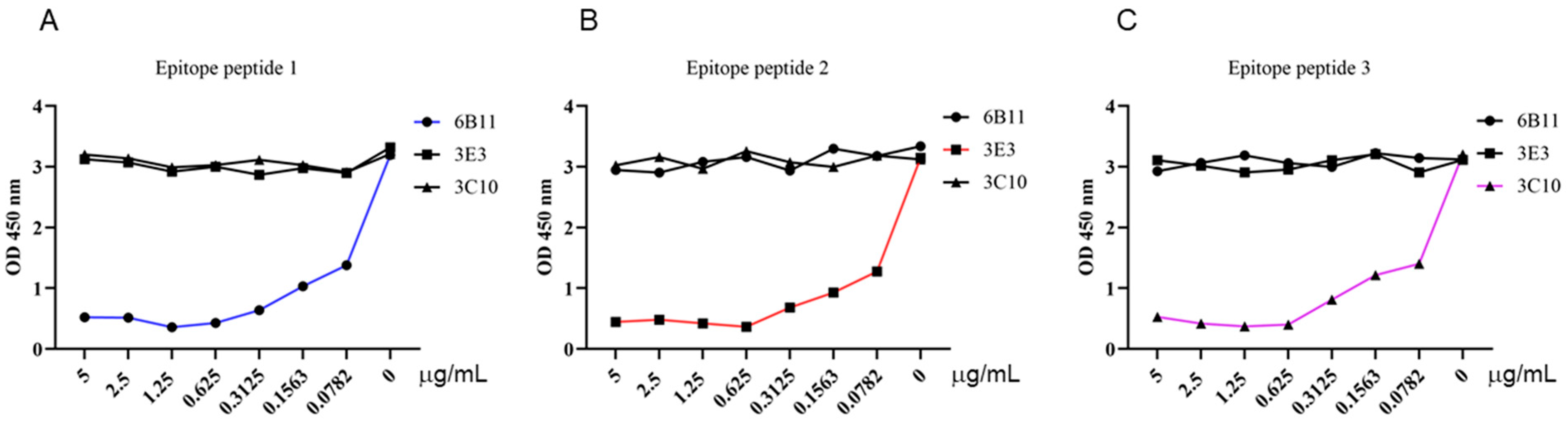
3.3. Identification of the Precise Antigenic Epitopes Targeted by p54 mAbs
3.4. Validation of the p54 Epitopes by Competitive ELISA and Establishment of Indirect Epitope ELISA for Detection of ASF Antibody
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, S.; Miao, C.; Liu, W.; Zhang, G.; Shao, J.; Chang, H. Structure and function of African swine fever virus proteins: Current understanding. Front. Microbiol. 2023, 14, 1043129. [Google Scholar] [CrossRef]
- Salguero, F.J. Comparative Pathology and Pathogenesis of African Swine Fever Infection in Swine. Front. Vet. Sci. 2020, 7, 282. [Google Scholar] [CrossRef] [PubMed]
- Gaudreault, N.N.; Madden, D.W.; Wilson, W.C.; Trujillo, J.D.; Richt, J.A. African Swine Fever Virus: An Emerging DNA Arbovirus. Front. Vet. Sci. 2020, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- Penrith, M.L.; Vosloo, W.; Jori, F.; Bastos, A.D. African swine fever virus eradication in Africa. Virus Res. 2013, 173, 228–246. [Google Scholar] [CrossRef]
- Kolbasov, D.; Titov, I.; Tsybanov, S.; Gogin, A.; Malogolovkin, A. African Swine Fever Virus, Siberia, Russia, 2017. Emerg. Infect. Dis. 2018, 24, 796–798. [Google Scholar] [CrossRef]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K.; et al. Emergence of African Swine Fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zheng, H. Insights and progress on epidemic characteristics, pathogenesis, and preventive measures of African swine fever virus: A review. Virulence 2025, 16, 2457949. [Google Scholar] [CrossRef]
- Salas, M.L.; Andrés, G. African swine fever virus morphogenesis. Virus Res. 2013, 173, 29–41. [Google Scholar] [CrossRef]
- Jia, N.; Ou, Y.; Pejsak, Z.; Zhang, Y.; Zhang, J. Roles of African Swine Fever Virus Structural Proteins in Viral Infection. J. Vet. Res. 2017, 61, 135–143. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; García-Escudero, R.; Salas, M.L.; Andrés, G. African swine fever virus structural protein p54 is essential for the recruitment of envelope precursors to assembly sites. J. Virol. 2004, 78, 4299–4313. [Google Scholar] [CrossRef]
- Alcaraz, C.; Brun, A.; Ruiz-Gonzalvo, F.; Escribano, J.M. Cell culture propagation modifies the African swine fever virus replication phenotype in macrophages and generates viral subpopulations differing in protein p54. Virus Res. 1992, 23, 173–182. [Google Scholar] [CrossRef]
- Alonso, C.; Miskin, J.; Hernáez, B.; Fernandez-Zapatero, P.; Soto, L.; Cantó, C.; Rodríguez-Crespo, I.; Dixon, L.; Escribano, J.M. African swine fever virus protein p54 interacts with the microtubular motor complex through direct binding to light-chain dynein. J. Virol. 2001, 75, 9819–9827. [Google Scholar] [CrossRef] [PubMed]
- García-Mayoral, M.F.; Rodríguez-Crespo, I.; Bruix, M. Structural models of DYNLL1 with interacting partners: African swine fever virus protein p54 and postsynaptic scaffolding protein gephyrin. FEBS Lett. 2011, 585, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Hernáez, B.; Díaz-Gil, G.; García-Gallo, M.; Ignacio Quetglas, J.; Rodríguez-Crespo, I.; Dixon, L.; Escribano, J.M.; Alonso, C. The African swine fever virus dynein-binding protein p54 induces infected cell apoptosis. FEBS Lett. 2004, 569, 224–228. [Google Scholar] [CrossRef]
- Neilan, J.G.; Zsak, L.; Lu, Z.; Burrage, T.G.; Kutish, G.F.; Rock, D.L. Neutralizing antibodies to African swine fever virus proteins p30, p54, and p72 are not sufficient for antibody-mediated protection. Virology 2004, 319, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.L.; Parkhouse, R.M.E.; Penedos, A.R.; Martins, C.; Leitão, A. Systematic analysis of longitudinal serological responses of pigs infected experimentally with African swine fever virus. J. Gen. Virol. 2007, 88 Pt 9, 2426–2434. [Google Scholar] [CrossRef]
- Xu, Z.; Hu, Y.; Li, J.; Wang, A.; Meng, X.; Chen, L.; Wei, J.; Tong, W.; Kong, N.; Yu, L.; et al. Screening and identification of the dominant antigens of the African swine fever virus. Front. Vet. Sci. 2023, 10, 1175701. [Google Scholar] [CrossRef]
- Gallardo, C.; Reis, A.L.; Kalema-Zikusoka, G.; Malta, J.; Soler, A.; Blanco, E.; Parkhouse, R.M.; Leitão, A. Recombinant antigen targets for serodiagnosis of African swine fever. Clin. Vaccine Immunol. 2009, 16, 1012–1020. [Google Scholar] [CrossRef]
- Onyilagha, C.; Quizon, K.; Zhmendak, D.; El Kanoa, I.; Truong, T.; Ambagala, T.; Clavijo, A.; Le, V.P.; Babiuk, S.; Ambagala, A. Development and Validation of an Indirect and Blocking ELISA for the Serological Diagnosis of African Swine Fever. Pathogens 2024, 13, 981. [Google Scholar] [CrossRef]
- Huang, C.; Cao, C.; Xu, Z.; Lin, Y.; Wu, J.; Weng, Q.; Liu, Z.; Jin, Y.; Chen, P.; Hua, Q. A blocking ELISA based on virus-like nanoparticles chimerized with an antigenic epitope of ASFV P54 for detecting ASFV antibodies. Sci. Rep. 2023, 13, 19928. [Google Scholar] [CrossRef]
- Jin, J.; Bai, Y.; Zhang, Y.; Lu, W.; Zhang, S.; Zhao, X.; Sun, Y.; Wu, Y.; Zhang, A.; Zhang, G.; et al. Establishment and characterization of a novel indirect ELISA method based on ASFV antigenic epitope-associated recombinant protein. Int. J. Biol. Macromol. 2023, 253 Pt 7, 127311. [Google Scholar] [CrossRef]
- Xu, L.; Cao, C.; Yang, Z.; Jia, W. Identification of a conservative site in the African swine fever virus p54 protein and its preliminary application in a serological assay. J. Vet. Sci. 2022, 23, e55. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.C.; Nguyen, T.N.; Poonsuk, K.; McVey, D.S.; Vu, H.L.X. Lipid nanoparticle-encapsulated DNA vaccine encoding African swine fever virus p54 antigen elicits robust immune responses in pigs. Vet. Microbiol. 2025, 305, 110508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.Y.; Zhang, K.; Deng, D.; Peng, K.; Zheng, W.; Chen, N.; Liu, P.; Shang, S.; Li, R.; Zhu, J. Characterization of three African swine fever viruses from different clinical settings revealed a potential attenuation mechanism. Animal Dis. 2024, 4, 24. [Google Scholar] [CrossRef]
- Xia, N.; Cao, Q.; Liu, A.; Zhang, J.; Han, H.; Jiao, J.; He, P.; Sun, Z.; Xu, Z.; Zheng, W.; et al. Identification of a New Conserved Antigenic Epitope by Specific Monoclonal Antibodies Targeting the African Swine Fever Virus Capsid Protein p17. Vet. Sci. 2024, 11, 650. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Li, S.; Cao, Q.; Gu, H.; Zhang, J.; Zhang, Y.; Zhang, K.; Zheng, W.; Chen, N.; Shang, S.; et al. Development of Specific Monoclonal Antibodies against Porcine RIG-I-like Receptors Revealed the Species Specificity. Int. J. Mol. Sci. 2023, 24, 4118. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, K.; Sun, S.; He, P.; Deng, D.; Zhang, P.; Zheng, W.; Chen, N.; Zhu, J. Specific Monoclonal Antibodies against African Swine Fever Virus Protease pS273R Revealed a Novel and Conserved Antigenic Epitope. Int. J. Mol. Sci. 2024, 25, 8906. [Google Scholar] [CrossRef]
- Wu, K.; Liu, J.; Wang, L.; Fan, S.; Li, Z.; Li, Y.; Yi, L.; Ding, H.; Zhao, M.; Chen, J. Current State of Global African Swine Fever Vaccine Development under the Prevalence and Transmission of ASF in China. Vaccines 2020, 8, 531. [Google Scholar] [CrossRef]
- Sun, E.; Zhang, Z.; Wang, Z.; He, X.; Zhang, X.; Wang, L.; Wang, W.; Huang, L.; Xi, F.; Huangfu, H.; et al. Emergence and prevalence of naturally occurring lower virulent African swine fever viruses in domestic pigs in China in 2020. Sci. China Life Sci. 2021, 64, 752–765. [Google Scholar] [CrossRef]
- Petrovan, V.; Murgia, M.V.; Wu, P.; Lowe, A.D.; Jia, W.; Rowland, R.R.R. Epitope mapping of African swine fever virus (ASFV) structural protein, p54. Virus Res. 2020, 279, 197871. [Google Scholar] [CrossRef]
- Heimerman, M.E.; Murgia, M.V.; Wu, P.; Lowe, A.D.; Jia, W.; Rowland, R.R. Linear epitopes in African swine fever virus p72 recognized by monoclonal antibodies prepared against baculovirus-expressed antigen. J. Vet. Diagn. Invest. 2018, 30, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qiao, S.; Liu, J.; Zhou, Y.; Tong, W.; Dong, S.; Liu, C.; Jiang, Y.; Guo, Z.; Zheng, H.; et al. A highly efficient indirect ELISA and monoclonal antibody established against African swine fever virus pK205R. Front. Immunol. 2022, 13, 1103166. [Google Scholar] [CrossRef] [PubMed]
- Cubillos, C.; Gómez-Sebastian, S.; Moreno, N.; Nuñez, M.C.; Mulumba-Mfumu, L.K.; Quembo, C.J.; Heath, L.; Etter, E.M.; Jori, F.; Escribano, J.M.; et al. African swine fever virus serodiagnosis: A general review with a focus on the analyses of African serum samples. Virus Res. 2013, 173, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Netherton, C.L.; Goatley, L.C.; Reis, A.L.; Portugal, R.; Nash, R.H.; Morgan, S.B.; Gault, L.; Nieto, R.; Norlin, V.; Gallardo, C.; et al. Identification and Immunogenicity of African Swine Fever Virus Antigens. Front. Immunol. 2019, 10, 1318. [Google Scholar] [CrossRef]
- Potocnakova, L.; Bhide, M.; Pulzova, L.B. An Introduction to B-Cell Epitope Mapping and In Silico Epitope Prediction. J. Immunol. Res. 2016, 2016, 6760830. [Google Scholar] [CrossRef]
- Desmet, C.; Coelho-Cruz, B.; Mehn, D.; Colpo, P.; Ruiz-Moreno, A. ASFV epitope mapping by high density peptides microarrays. Virus Res. 2024, 339, 199287. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Han, D.; Zhang, Y.; Zhang, K.; Du, N.; Tong, W.; Li, G.; Zheng, H.; Liu, C.; Gao, F.; et al. Identification of one novel epitope targeting p54 protein of African swine fever virus using monoclonal antibody and development of a capable ELISA. Res. Vet. Sci. 2021, 141, 19–25. [Google Scholar] [CrossRef]
- Wang, A.; Jiang, M.; Liu, H.; Liu, Y.; Zhou, J.; Chen, Y.; Ding, P.; Wang, Y.; Pang, W.; Qi, Y.; et al. Development and characterization of monoclonal antibodies against the N-terminal domain of African swine fever virus structural protein, p54. Int. J. Biol. Macromol. 2021, 180, 203–211. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, G.; Dong, H.; Wu, S.; Du, Y.; Wan, B.; Ji, P.; Wu, Y.; Jiang, D.; Zhuang, G.; et al. Identification of a Linear B Cell Epitope on p54 of African Swine Fever Virus Using Nanobodies as a Novel Tool. Microbiol. Spectr. 2023, 11, e0336222. [Google Scholar] [CrossRef]
- Zheng, N.; Li, C.; Hou, H.; Chen, Y.; Zhang, A.; Han, S.; Wan, B.; Wu, Y.; He, H.; Wang, N.; et al. A Novel Linear B-Cell Epitope on the P54 Protein of African Swine Fever Virus Identified Using Monoclonal Antibodies. Viruses 2023, 15, 867. [Google Scholar] [CrossRef]
- Gao, Y.; Xia, T.; Bai, J.; Zhang, L.; Zheng, H.; Jiang, P. Preparation of Monoclonal Antibodies against the Viral p54 Protein and a Blocking ELISA for Detection of the Antibody against African Swine Fever Virus. Viruses 2022, 14, 2335. [Google Scholar] [CrossRef] [PubMed]
- Imdhiyas, M.; Sen, S.; Barman, N.; Buragohain, L.; Malik, Y.; Kumar, S. Computational analysis of immunogenic epitopes in the p30 and p54 proteins of African swine fever virus. J. Biomol. Struct. Dyn. 2023, 41, 7480–7489. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Nurmoja, I.; Cano-Gómez, C.; Cvetkova, S.; Frant, M.; Woźniakowski, G.; Simón, A.; Pérez, C.; Nieto, R.; et al. Dynamics of African swine fever virus (ASFV) infection in domestic pigs infected with virulent, moderate virulent and attenuated genotype II ASFV European isolates. Transbound. Emerg. Dis. 2021, 68, 2826–2841. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Nurmoja, I.; Soler, A.; Delicado, V.; Simón, A.; Martin, E.; Perez, C.; Nieto, R.; Arias, M. Evolution in Europe of African swine fever genotype II viruses from highly to moderately virulent. Vet. Microbiol. 2018, 219, 70–79. [Google Scholar]
- Tesfagaber, W.; Wang, L.; Tsegay, G.; Hagoss, Y.T.; Zhang, Z.; Zhang, J.; Huangfu, H.; Xi, F.; Li, F.; Sun, E.; et al. Characterization of Anti-p54 Monoclonal Antibodies and Their Potential Use for African Swine Fever Virus Diagnosis. Pathogens 2021, 10, 178. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, J.; Peng, Y.; Xie, Y.; Xiao, Y. A Promising Tool in Serological Diagnosis: Current Research Progress of Antigenic Epitopes in Infectious Diseases. Pathogens 2022, 11, 1095. [Google Scholar] [CrossRef]
- Qu, H.; Ge, S.; Zhang, Y.; Wu, X.; Wang, Z. A systematic review of genotypes and serogroups of African swine fever virus. Virus Genes 2022, 58, 77–87. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhang, K.; Sun, S.; He, P.; Deng, D.; Lv, H.; Xie, M.; Zhang, P.; Zheng, W.; Chen, N.; et al. The Novel Antigenic Epitopes of African Swine Fever Virus Inner Membrane p54 Protein Revealed by Monoclonal Antibodies. Animals 2025, 15, 1296. https://doi.org/10.3390/ani15091296
Zhang J, Zhang K, Sun S, He P, Deng D, Lv H, Xie M, Zhang P, Zheng W, Chen N, et al. The Novel Antigenic Epitopes of African Swine Fever Virus Inner Membrane p54 Protein Revealed by Monoclonal Antibodies. Animals. 2025; 15(9):1296. https://doi.org/10.3390/ani15091296
Chicago/Turabian StyleZhang, Jiajia, Kaili Zhang, Shaohua Sun, Ping He, Dafu Deng, Hanrong Lv, Mingwang Xie, Pingping Zhang, Wanglong Zheng, Nanhua Chen, and et al. 2025. "The Novel Antigenic Epitopes of African Swine Fever Virus Inner Membrane p54 Protein Revealed by Monoclonal Antibodies" Animals 15, no. 9: 1296. https://doi.org/10.3390/ani15091296
APA StyleZhang, J., Zhang, K., Sun, S., He, P., Deng, D., Lv, H., Xie, M., Zhang, P., Zheng, W., Chen, N., Bai, J., & Zhu, J. (2025). The Novel Antigenic Epitopes of African Swine Fever Virus Inner Membrane p54 Protein Revealed by Monoclonal Antibodies. Animals, 15(9), 1296. https://doi.org/10.3390/ani15091296