


Amino Acids as Dietary Additives for Enhancing Fish Welfare in Aquaculture



Simple Summary
Abstract
1. Introduction
2. General Amino Acid Physiological Roles in Animals
3. Stress Responses Depending on Amino Acids
3.1. Immunological Responses
3.2. Effects Involving Energy Metabolism
3.3. Endocrine and Neuroendocrine Processes
3.4. Responses Related to Oxidative Stress
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA | amino acid |
| ACH50 | alternative complement pathway |
| Ala-Gln | alanine–glutamine |
| ALT | alanine transaminase |
| Arg | arginine |
| Asp | aspartate |
| ATP | adenosine triphosphate |
| CAT | catalase |
| EAA | essential amino acid |
| FW | final weight |
| GALT | gut-associated lymphoid tissue |
| gLYS | g-type lysozyme |
| GPT | alanine aminotransferase |
| Gpx | glutathione peroxidase |
| GOT | aspartate aminotransferase |
| GSH | glutathione |
| Ile | isoleucine |
| IMTA | integrated multitrophic aquaculture |
| HAMP-1 | hepcidin antimicrobial peptide 1 |
| HIF-1 | hypoxia-inducible factor 1 |
| HPI | hypothalamic–pituitary–interrenal |
| Hsp70 | mitochondrial heat shock protein 70 |
| Leu | leucine |
| LDL-C | low-density lipoprotein cholesterol |
| Met | methionine |
| MIP1-alpha | macrophage inflammatory protein 1α |
| NEAA | nutritionally non-essential amino acids |
| NEFA | non-esterified fatty acid |
| NO | nitric oxide |
| ONOO− | peroxynitrite |
| PER | protein efficiency ratio |
| Phe | phenylalanine |
| pomc-a | proopiomelanocortin-derived hormone |
| PWG | percent weight gain |
| RGR | relative growth rate |
| ROS | oxygen-containing reactive species |
| SGR | specific growth rate |
| SOD | superoxide dismutase |
| Tau | taurine |
| TC | total cholesterol |
| TG | triglyceride |
| Trp | tryptophan |
| Tyr | tyrosine |
| Val | valine |
| WG | weight gain |
References
- Botreau, R.; Veissier, I.; Perny, P. Overall assessment of animal welfare: Strategy adopted in welfare quality. Animal Welfare 2009, 18, 363–370. [Google Scholar] [CrossRef]
- Saraiva, J.L.; Faccenda, F.; Cabrera-Álvarez, M.J.; Povinelli, M.; Hubbard, P.C.; Cerqueira, M.; Farinha, A.P.; Secci, G.; Tignani, M.V.; Pulido Rodriguez, L.F.; et al. Welfare of rainbow trout at slaughter: Integrating behavioural, physiological, proteomic and quality indicators and testing a novel fast-chill stunning method. Aquaculture 2024, 581, 740443. [Google Scholar] [CrossRef]
- Wolke, R.E.; Bengston, D.A.; Rice, M.A.; Whittaker, M. Diseases of Cultured Flatfish: Diagnosis and Prevention; NRAC Publication No. 00-0001; Northeastern Regional Aquaculture Center, University of Massachusetts: Dartmouth, MA, USA, 2000. [Google Scholar]
- Read, N. Fish Farmer’s Perspective of Welfare. In Fish Welfare.; Branson, E.B., Ed.; Blackwell Publishing: Oxford, UK, 2008; pp. 101–110. [Google Scholar]
- Valcarce, D.G.; Riesco, M.F.; Martínez-Vázquez, J.M.; Rodríguez, J.L.; Robles, V. Impact of different levels of handling on Solea senegalensis culture: Effects on growth and molecular markers of stress. Fish Physiol. Biochem. 2024, 50, 1987–2000. [Google Scholar] [CrossRef]
- Montero, D.; Izquierdo, M.S.; Tort, L.; Robaina, L.; Vergara, J.M. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiol. Biochem. 1999, 20, 53–60. [Google Scholar] [CrossRef]
- Herrera, M.; Ruiz-Jarabo, I.; Hachero, I.; Vargas-Chacoff, L.; Amo, A.; Mancera, J.M. Stocking density affects growth and metabolic parameters in the brill (Scophthalmus rhombus). Aquac. Int. 2012, 20, 1041–1152. [Google Scholar] [CrossRef]
- Herrera, M.; Ruiz-Jarabo, I.; Vargas-Chacoff, L.; De La Roca, E.; Mancera, J.M. Metabolic enzyme activities in relation to crowding stress in the wedge sole (Dicologoglossa cuneata). Aquacult. Res. 2015, 46, 2808–2818. [Google Scholar] [CrossRef]
- Li, X.; Zheng, S.; Wu, G. Nutrition and functions of amino acids in fish. In Amino Acids in Nutrition and Health; Advances in Experimental Medicine and Biology; Wu, G., Ed.; Springer: Cham, Switzerland, 2021; pp. 133–168. [Google Scholar]
- Castilla-Gavilán, M.; Guerra-García, J.M.; Hachero-Cruzado, I.; Herrera, M. Understanding carbon footprint in sustainable land-based marine aquaculture: Exploring production techniques. J. Mar. Sci. Eng. 2024, 12, 1192. [Google Scholar] [CrossRef]
- Ellis, T.; North, B.; Scott, A.P.; Bromage, N.R.; Porter, M.; Gadd, D. The relationships between stocking density and welfare in farmed rainbow trout. J. Fish Biol. 2002, 61, 493–531. [Google Scholar] [CrossRef]
- Liu, B.; Fei, F.; Li, X.; Wang, X.; Huang, B. Effects of stocking density on stress response, innate immune parameters, and welfare of turbot (Scophthalmus maximus). Aquac. Int. 2019, 27, 1599–1612. [Google Scholar] [CrossRef]
- Long, L.; Zhang, H.; Ni, Q.; Liu, H.; Wu, F.; Wang, X. Effects of stocking density on growth, stress, and immune responses of juvenile Chinese sturgeon (Acipenser sinensis) in a recirculating aquaculture system. CBPC 2019, 219, 25–34. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Cabrera-Álvarez, M.J.; Maia, C.M.; Saraiva, J.L. Environmental enrichment in fish aquaculture: A review of fundamental and practical aspects. Rev. Aquacult. 2022, 14, 704–728. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, L.; Zhang, X. Environmental enrichment increases aquatic animal welfare: A systematic review and meta-analysis. Rev. Aquacult. 2022, 14, 1120–1135. [Google Scholar] [CrossRef]
- Branson, E.B. Fish Welfare; Blackwell Publlishing: Oxford, UK, 2008; p. 316. [Google Scholar]
- Ciji, A.; Akhtar, M.S. Stress management in aquaculture: A review of dietary interventions. Rev. Aquacult. 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- Herrera, M.; Mancera, J.M.; Costas, B. The use of dietary additives in fish stress mitigation: Comparative endocrine and physiological responses. Front. Endocrinol. 2019, 10, 447. [Google Scholar] [CrossRef]
- Conceição, L.E.C.; Aragão, C.; Dias, J.; Costas, B.; Terova, G.; Martins, C.; Tort, L. Dietary nitrogen and fish welfare. Fish Physiol. Biochem. 2012, 38, 119–141. [Google Scholar] [CrossRef]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino. Acids. 2009, 37, 43–53. [Google Scholar] [CrossRef]
- Xing, S.; Liang, X.; Zhang, X.; Oliva-Teles, A.; Peres, H.; Li, M.; Wang, H.; Mai, K.; Kaushik, S.; Xue, M. Essential amino acid requirements of fish and crustaceans, a meta-analysis. Rev. Aquac. 2024, 16, 1069–1086. [Google Scholar] [CrossRef]
- Wu, G. Dietary requirements of synthesizable amino acids by animals: A paradigm shift in protein nutrition. JASB 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Zhang, Y.; Baranov, P.V.; Atkins, J.F.; Gladyshev, V.N. Pyrrolysine and selenocysteine use dissimilar decoding strategies. JBC 2005, 280, 20740–20751. [Google Scholar] [CrossRef]
- Wu, G. Functional amino acids in nutrition and health. Amino. Acids. 2013, 45, 407–411. [Google Scholar] [CrossRef]
- Gutiérrez-Preciado, A.; Romero, H.; Peimbert, M. An evolutionary perspective on amino acids. Nat. Educ. 2010, 3, 29. [Google Scholar]
- Carvalho, D.P.; Dupuy, C. Thyroid hormone biosynthesis and release. Mol. Cell. Endocrinol. 2017, 458, 6–15. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, J.P.F. Amino Acids in Animal Nutrition; CABI Publishing: Wallingford, UK, 2003. [Google Scholar]
- Beaumont, M.; Blachier, F. Amino Acids in Intestinal Physiology and Health. In Amino Acids in Nutrition and Health; Advances in Experimental Medicine and Biology; Wu, G., Ed.; Springer: Cham, Switzerland, 2020; pp. 1–20. [Google Scholar]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Ball, R.O.; Atkinson, J.L.; Bayley, H.S. Proline as an essential amino acid for the young pig. Br. J. Nutr. 1986, 55, 659–668. [Google Scholar] [CrossRef]
- Wu, G. Functional amino acids in growth, reproduction, and health. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef]
- Yoneda, J.; Andou, A.; Takehana, K. Regulatory roles in amino acids in immune response. Curr. Rheumatol. Rev. 2009, 5, 252–258. [Google Scholar] [CrossRef]
- Cowey, C.B. Amino acid requirements of fish: A critical appraisal of present values. Aquaculture 1994, 124, 1–11. [Google Scholar] [CrossRef]
- Costas, B.; Rêgo, P.C.N.P.; Conceição, L.E.C.; Dias, J.; Afonso, A. Dietary arginine supplementation decreases plasma cortisol levels and modulates immune mechanisms in chronically stressed turbot (Scophthalmus maximus). Aquac. Nutr. 2013, 19, 25–38. [Google Scholar] [CrossRef]
- Höglund, E.; Sørensen, C.; Bakke, M.J.; Nilsson, G.E.; Øverli, Ø. Attenuation of stress-induced anorexia in brown trout (Salmo trutta) by pre-treatment with dietary L-tryptophan. Br. J. Nutr. 2007, 97, 786–789. [Google Scholar] [CrossRef]
- Yang, B.; Shen, Y.; Monroig, O.; Zhao, W.; Bao, Y.; Tao, S.; Jiao, L.; Zhou, Q.; Min, J. The ameliorative role of methionine in hepatic steatosis and stress response in juvenile black seabream (Acanthopagrus schlegelii) fed with a high-fat diet. Aquaculture 2024, 580, 740306. [Google Scholar] [CrossRef]
- Souto, C.N.; de Menezes, L.L.C.; Proença, D.C.; Bueno, G.W.; Guimarães, I.G. Pirarucu requires taurine to maximize growth and antioxidant status when fed diets high in plant-based ingredients. Aquaculture 2024, 590, 741022. [Google Scholar] [CrossRef]
- Schreck, C.B.; Tort, L.; Farrell, A.; Brauner, C. Biology of Stress in Fish; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Wolkers, C.P.B.; Serra, M.; Hoshiba, M.A.; Urbinati, E.C. Dietary L-tryptophan alters aggression in juvenile matrinxa Brycon amazonicus. Fish Physiol. Biochem. 2012, 38, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Papoutsoglou, S.E.; Karakatsouli, N.; Chiras, G. Dietary L-tryptophan and tank colour effects on growth performance of rainbow trout (Oncorhynchus mykiss) juveniles reared in a recirculating water system. Aquacult. Eng. 2005, 32, 277–284. [Google Scholar] [CrossRef]
- Tejpal, C.S.; Pal, A.K.; Sahu, N.P.; Kumar, J.A.; Muthappa, N.A.; Vidya, S.; Rajan, M.G. Dietary supplementation of L-tryptophan mitigates crowding stress and augments the growth in Cirrhinus mrigala fingerlings. Aquaculture 2009, 293, 272–277. [Google Scholar] [CrossRef]
- Teixeira, C.; Pedrosa, R.; Castro, C.; Magalhães, R.; Matos, E.; Oliva-Teles, A.; Peres, H.; Pérez-Jiménez, A. Dietary tryptophan supplementation implications on performance, plasma metabolites, and amino acid catabolism enzymes in meagre (Argyrosomus regius). Fishes 2023, 8, 141. [Google Scholar] [CrossRef]
- Herrera, M.; Matias, A.C.; Soares, F.; Ribeiro, L.; Moreira, M.; Salamanca, N.; Jerez-Cepa, I.; Mancera, J.M.; Astola, A. Effect of amino acid supplementation and stress on expression of molecular markers in meagre (Argyrosomus regius). Aquaculture 2021, 534, 736238. [Google Scholar] [CrossRef]
- Herrera, M.; Fernández-Alacid, L.; Sanahuja, I.; Ibarz, A.; Salamanca, N.; Morales, E.; Giráldez, I. Physiological and metabolic effects of a tryptophan-enriched diet to face up chronic stress in meagre (Argyrosomus regius). Aquaculture 2020, 522, 735102. [Google Scholar] [CrossRef]
- Fernández-Alacid, L.; Sanahuja, I.; Ordóñez-Grande, B.; Sánchez-Nuño, S.; Herrera, M.; Ibarz, A. Skin mucus metabolites and cortisol in meagre fed acute stress-attenuating diets: Correlations between plasma and mucus. Aquaculture 2019, 499, 185–194. [Google Scholar] [CrossRef]
- Asencio-Alcudia, G.; Andree, K.B.; Giraldez, I.; Tovar-Ramirez, D.; Alvarez-González, A.; Herrera, M.; Gisbert, E. Stressors due to handling impair gut immunity in meagre (Argyrosomus regius): The compensatory role of dietary L-tryptophan. Front. Physiol. 2019, 10, 547. [Google Scholar] [CrossRef]
- González-Silvera, D.; Herrera, M.; Giráldez, I.; Esteban, M.A. Effects of the dietary tryptophan and aspartate on the immune response of meagre (Argyrosomus regius) after stress. Fishes 2018, 3, 6. [Google Scholar] [CrossRef]
- Cabanillas-Gámez, M.; López, L.M.; Galaviz, M.A.; True, C.D.; Bardullas, U. Effect of L-tryptophan supplemented diets on serotonergic system and plasma cortisol in Totoaba macdonaldi (Gilbert, 1890) juvenile exposed to acute stress by handling and hypoxia. Aquac. Res. 2018, 49, 847–857. [Google Scholar] [CrossRef]
- Herrera, M.; Herves, M.A.; Giráldez, I.; Skar, K.; Mogren, H.; Mortensen, A.; Puvanendran, V. Effects of amino acid supplementations on metabolic and physiological parameters in Atlantic cod (Gadus morhua) under stress. Fish Physiol. Biochem. 2017, 43, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Salamanca, N.; Morales, E.; Ruiz-Azcona, P.; Herrera, M. Endocrine and metabolic effects of Trp-enriched diets for attenuation of chronic stress in the Senegal soles (Solea senegalensis). Aquaculture 2020, 523, 735173. [Google Scholar] [CrossRef]
- Azeredo, R.; Machado, M.; Afonso, A.; Fierro-Castro, C.; Reyes-López, F.E.; Tort, L.; Gesto, M.; Conde-Sieira, M.; Míguez, J.M.; Soengas, J.L.; et al. Neuroendocrine and immune responses undertake different fates following tryptophan or methionine dietary treatment: Tales from a teleost model. Front. Immunol. 2017, 8, 1226. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Mirghaed, A.T.; Mazandarani, M.; Zoheiri, F. Serum cortisol, glucose, thyroid hormones’ and non-specific immune responses of Persian sturgeon, Acipenser persicus to exogenous tryptophan and acute stress. Aquaculture 2016, 462, 17–23. [Google Scholar] [CrossRef]
- Kumar, P.; Saurabh, S.; Pal, A.K.; Sahu, N.P.; Arasu, A.R.T. Stress mitigating and growth enhancing effect of dietary tryptophan in rohu (Labeo rohita, Hamilton, 1822) fingerlings. Fish Physiol. Biochem. 2014, 40, 1325–1338. [Google Scholar] [CrossRef]
- Shi, L.; Zhao, Y.; Zhou, J.; Zhai, H.; Wei, L.; Liu, Z.; Wang, W.; Jiang, Z.; Ren, T.; Han, Y. Dietary taurine impacts the growth, amino acid profile and resistance against heat stress of tiger puffer (Takifugu rubripes). Aquac. Nutr. 2020, 26, 1691–1701. [Google Scholar] [CrossRef]
- Tian, Q.; Hu, Y.; Mao, P.; Xie, J.; Fang, B.; Peng, H. Effect of dietary taurine supplementation on growth, intestine structure and resistance to acute crowding stress in juvenile black carp (Mylopharyngodon piceus) fed low fish meal diets. J. Fish. China 2016, 40, 1330–1339. [Google Scholar]
- Ceccotti, C.; Al-Sulaivany, B.S.A.; Al-Habbib, O.A.M.; Saroglia, M.; Rimoldi, S.; Terova, G. Protective effect of dietary taurine from ROS production in European seabass under conditions of forced swimming. Animals 2019, 9, 607. [Google Scholar] [CrossRef]
- Salamanca, N.; Giráldez, I.; Morales, E.; de La Rosa, I.; Herrera, M. Phenylalanine and tyrosine as feed additives for reducing stress and enhancing welfare in gilthead seabream and meagre. Animals 2021, 11, 45. [Google Scholar] [CrossRef]
- Costas, B.; Conceição, L.E.; Dias, J.; Novoa, B.; Figueras, A.; Afonso, A. Dietary arginine and repeated handling increase disease resistance and modulate innate immune mechanisms of Senegalese sole (Solea senegalensis Kaup, 1858). Fish Shellfish Immunol. 2011, 31, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Guo, G.L.; Sun, L.; Yang, Q.S.; Wang, G.Q.; Qin, G.X.; Zhang, D.M. Effects of Ala-Gln feeding strategies on growth, metabolism, and crowding stress resistance of juvenile Cyprinus carpio var. Jian. Fish Shellfish. Immunol. 2016, 51, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Zhen, L.; Feng, L.; Jiang, W.; Wu, P.; Liu, Y.; Tang, L.; Li, S.W.; Zhong, C.B.; Zhou, X.O. Exploring the novel benefits of leucine: Protecting nitrite-induced liver damage in sub-adult grass carp (Ctenopharyngodon idella) through regulating mitochondria quality control. Fish Shellfish. Immunol. 2024, 151, 109690. [Google Scholar] [CrossRef]
- Shi, L.; Zhai, H.; Wei, L.; Wang, X.; Ren, T.; Wang, W.; Jiang, c.; Han, Y. Interactive effect between dietary valine and isoleucine on growth performance, blood parameters and resistance against low salinity stress of Japanese flounder Paralichthys olivaceus. J. Appl. Ichthyol. 2021, 37, 285–294. [Google Scholar] [CrossRef]
- Bae, J.; Moniruzzaman, M.; Je, H.W.; Lee, S.; Choi, W.; Min, T.; Kim, K.W.; Bai, S.C. Evaluation of Gamma-Aminobutyric Acid (GABA) as a functional feed ingredient on growth performance, immune enhancement, and disease resistance in olive flounder (Paralichthys olivaceus) under high stocking density. Antioxidants 2024, 13, 647. [Google Scholar] [CrossRef]
- Tort, L.; Balasch, J.C. Stress and immunity in fish. In Principles of Fish Immunology; Buchmann, K., Secombes, C.J., Eds.; Springer: Cham, Switzerland, 2022; pp. 609–655. [Google Scholar]
- Nardocci, G.; Navarro, C.; Cortés, P.P.; Imarai, M.; Montoya, M.; Valenzuela, B.; Jara, P.; Acuña-Castillo, C.; Fernández, R. Neuroendocrine mechanisms for immune system regulation during stress in fish. Fish Shellfish. Immunol. 2014, 40, 531–538. [Google Scholar] [CrossRef]
- Saeij, J.P.J.; Verburg-van Kemenade, L.B.M.; van Muiswinkel, W.B.; Wiegertjes, G.F. Daily handling stress reduces resistance of carp to Trypanoplasma borreli: In vitro modulatory effects of cortisol on leukocyte function and apoptosis. Dev. Comp. Immunol. 2003, 27, 233–245. [Google Scholar] [CrossRef]
- Law, W.Y.; Chen, W.H.; Song, Y.L.; Dufour, S.; Chang, C.F. Differential in vitro suppressive effects of steroids on leukocyte phagocytosis in two teleosts, tilapia and common carp. Gen. Comp. Endocrinol. 2001, 121, 163–172. [Google Scholar] [CrossRef]
- Yao, K.; Fang, J.; Yin, Y.L.; Feng, Z.M.; Tang, Z.R.; Wu, G. Tryptophan metabolism in animals: Important roles in nutrition and health. Front. Biosci. 2011, 3, 286–297. [Google Scholar]
- Azeredo, R.; Machado, M.; Guardiola, F.A.; Cerezuela, R.; Afonso, A.; Peres, H.; Oliva- Teles, A.; Esteban, M.A.; Costas, B. Local immune response of two mucosal surfaces of the European seabass, Dicentrarchus labrax, fed tryptophan- or methionine supplemented diets. Fish Shellfish. Immunol. 2017, 70, 76–86. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Seve, B. Biological roles of tryptophan and its metabolism: Potential implications for pig feeding. Livest. Sci. 2007, 112, 23–32. [Google Scholar] [CrossRef]
- Mamane, Y.; Heylbroeck, C.; Génin, P.; Algarté, M.; Servant, M.J.; LePage, C.; DeLuca, C.; Kwon, H.; Lin, R.; Hiscott, J. Interferon regulatory factors: The next generation. Gene 1999, 237, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Satriano, J.; Ishizuka, S.; Archer, D.C.; Blantz, R.C.; Kelly, C.J. Regulation of intracelular polyamine biosynthesis and transport by NO and cytokines TNF-alpha and IFN-gamma. Am. J. Phys. 1999, 276, C892–C899. [Google Scholar]
- Buentello, J.A.; Gatlin, D.M. Nitric oxide production in activated macrophages from channel catfish (Ictalurus punctatus): Influence of dietary arginine and culture media. Aquaculture 1999, 179, 513–521. [Google Scholar] [CrossRef]
- Tafalla, C.; Novoa, B. Requirements for nitric oxide production by turbot (Scophthalmus maximus) head kidney macrophages. Dev. Comp. Immunol. 2000, 24, 623–631. [Google Scholar] [CrossRef]
- Dai, B.; Zhang, J.; Liu, M.; Lu, J.; Zhang, Y.; Xu, Y.; Miao, J.; Yin, Y. The role of Ca2+ mediated signaling pathways on the effect of taurine against Streptococcus uberis infection. Vet. Microbiol. 2016, 192, 26–33. [Google Scholar] [CrossRef]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; M’as, D.; Yin, Y. The role of methionine on metabolism, oxidative stress, and diseases. Amino. Acids. 2017, 49, 2091–2098. [Google Scholar] [CrossRef]
- Abe, T.; Takeya, K. Difference in gamma-aminobutyric acid content in vegetable soybean seeds. J. Jpn. Soc. Food Sci. Technol. 2005, 52, 545–549. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, L.; Li, H.; Chen, Q.; Li, N.; Li, J.; Zhao, Z.; Xiao, D.; Tang, T.; Bi, C.; et al. Insights and progress on the biosynthesis, metabolism, and physiological functions of gamma-aminobutyric acid (GABA): A review. PeerJ 2024, 12, e18712. [Google Scholar] [CrossRef]
- Kim, S.K.; Takeuchi, T.; Yokoyama, M.; Murata, Y. Effect of dietary supplementation with taurine, β-alanine and GABA on the growth of juvenile and fingerling Japanese flounder Paralichthys olivaceus. Fish. Sci. 2003, 69, 242–248. [Google Scholar] [CrossRef]
- Wu, F.; Liu, M.; Chen, C.; Chen, J.; Tan, Q. Effects of dietary Gamma aminobutyric acid on growth performance, antioxidant status, and feeding related gene expression of juvenile-grass carp, Ctenopharyngodon idellus. J. World Aquac. Soc. 2016, 47, 820–829. [Google Scholar] [CrossRef]
- Temu, V.; Kim, H.; Hamidoghli, A.; Park, M.; Won, S.; Oh, M.; Han, J.K.; Bai, S.C. Effects of dietary gamma aminobutyric acid in juvenile Nile tilapia, Orechromis niloticus. Aquaculture 2019, 507, 475–480. [Google Scholar] [CrossRef]
- Liu, H.; Yang, R.; Fu, Z.; Yu, G.; Li, M.; Dai, S.; Ma, Z.; Zong, H. Acute thermal stress increased enzyme activity and muscle energy distribution of yellowfin tuna. PLoS ONE 2023, 18, e0289606. [Google Scholar] [CrossRef]
- Hao, R.; Li, H.; Tian, Y.; Ru, X.; Deng, Q.; Zhu, K.; Yang, T.; Huang, Y.; Zhu, C. The effect of heat stress on energy metabolism, immune function, and oxidative stress of juvenile greater amberjack Seriola dumerili. Aquac. Res. 2024, 4406151. [Google Scholar] [CrossRef]
- Shafik, M.; Ibrahime, H.; Elyazeid, I.A.; Abass, O.; Saad, H.M. The stress of phenylalanine on rats to study the phenylketonuria at biochemical and molecular level. J. Appl. Pharm. Sci. 2014, 4, 24–29. [Google Scholar]
- Salamanca, N.; Moreno, O.; Giráldez, I.; Morales, E.; de la Rosa, I.; Herrera, M. Effects of dietary phenylalanine and tyrosine supplements on the chronic stress response in the seabream (Sparus aurata). Front. Physiol. 2022, 12, 775771. [Google Scholar] [CrossRef]
- Craig, P.M.; Moon, T.W. Methionine restriction affects the phenotypic and transcriptional response of rainbow trout (Oncorhynchus mykiss) to carbohydrate-enriched diets. Br. J. Nutr. 2013, 109, 402–412. [Google Scholar] [CrossRef]
- Hasek, B.E.; Stewart, L.K.; Henagan, T.M.; Boudreau, A.; Lenard, N.R.; Black, C.; Shin, J.; Huypens, P.; Malloy, V.L.; Plaisance, E.P.; et al. Dietary methionine restriction enhances metabolic flexibility and increases uncoupled respiration in both fed and fasted states. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R728–R739. [Google Scholar] [CrossRef]
- Kimball, S.R.; Jefferson, L.S. New functions for amino acids: Effects on gene transcription and translation. AJCN 2006, 83, 500–507. [Google Scholar] [CrossRef]
- Sharf, Y.; Khan, M.A. Effect of dietary isoleucine level on growth, protein retention efficiency, haematological parameter, lysozyme activity and serum antioxidant status of fingerling Channa punctatus (Bloch). Aquac. Nutr. 2020, 26, 908–920. [Google Scholar] [CrossRef]
- MacCormack, T.J.; Callaghan, N.I.; Sykes, A.V.; Driedzic, W.R. Taurine depresses cardiac contractility and enhances systemic heart glucose utilization in the cuttlefish, Sepia officinalis. J. Comp. Physiol. 2016, 186, 215–227. [Google Scholar] [CrossRef]
- Schaffer, S.W.; Ju Jong, C.; Kc, R.; Azuma, J. Physiological roles of taurine in heart and muscle. J. Biomed. Sci. 2010, 17, S2. [Google Scholar] [CrossRef] [PubMed]
- Dixon, T.A.M.; Rhyno, E.L.M.; El, N.; McGaw, S.P.; Otley, N.A.; Parker, K.S.; Buldo, E.C.; Pabody, C.M.; Savoie, M.; Cockshutt, A.; et al. Taurine depletion impairs cardiac function and affects tolerance to hypoxia and high temperatures in brook char (Salvelinus fontinalis). J. Exp. Biol. 2023, 226, jeb245092. [Google Scholar] [CrossRef] [PubMed]
- Farris, N.W.; Hamidoghli, A.; Bae, J.; Won, S.; Choi, W.; Biró, J.; Lee, S.; Bai, S.C. Dietary supplementation with γ-aminobutyric acid improves growth, digestive enzyme activity, non-specific immunity and disease resistance against Streptococcus iniae in juvenile olive flounder, Paralichthys olivaceus. Animals 2022, 12, 248. [Google Scholar] [CrossRef]
- Lee, S.; Moniruzzaman, M.; Farris, N.; Min, T.; Bai, S.C. Interactive effect of dietary gamma-aminobutyric acid (GABA) and water temperature on growth performance, blood plasma indices, heat shock proteins and GABAergic gene expression in juvenile olive flounder Paralichthys olivaceus. Metabolites 2023, 13, 619. [Google Scholar] [CrossRef]
- Matus-Ortega, G.; Romero-Aguilar, L.; Luqueno-Bocardo, O.I.; Hernandez-Morfin, K.; Guerra-Sanchez, G.; Matus-Ortega, M.; Pardo-Vazquez, J.P. Metabolic, Endocrine, and Regulatory Functions of Lactate in Gene Expression. Rev. Fac. Med. 2020, 63.5, 7–17. [Google Scholar] [CrossRef]
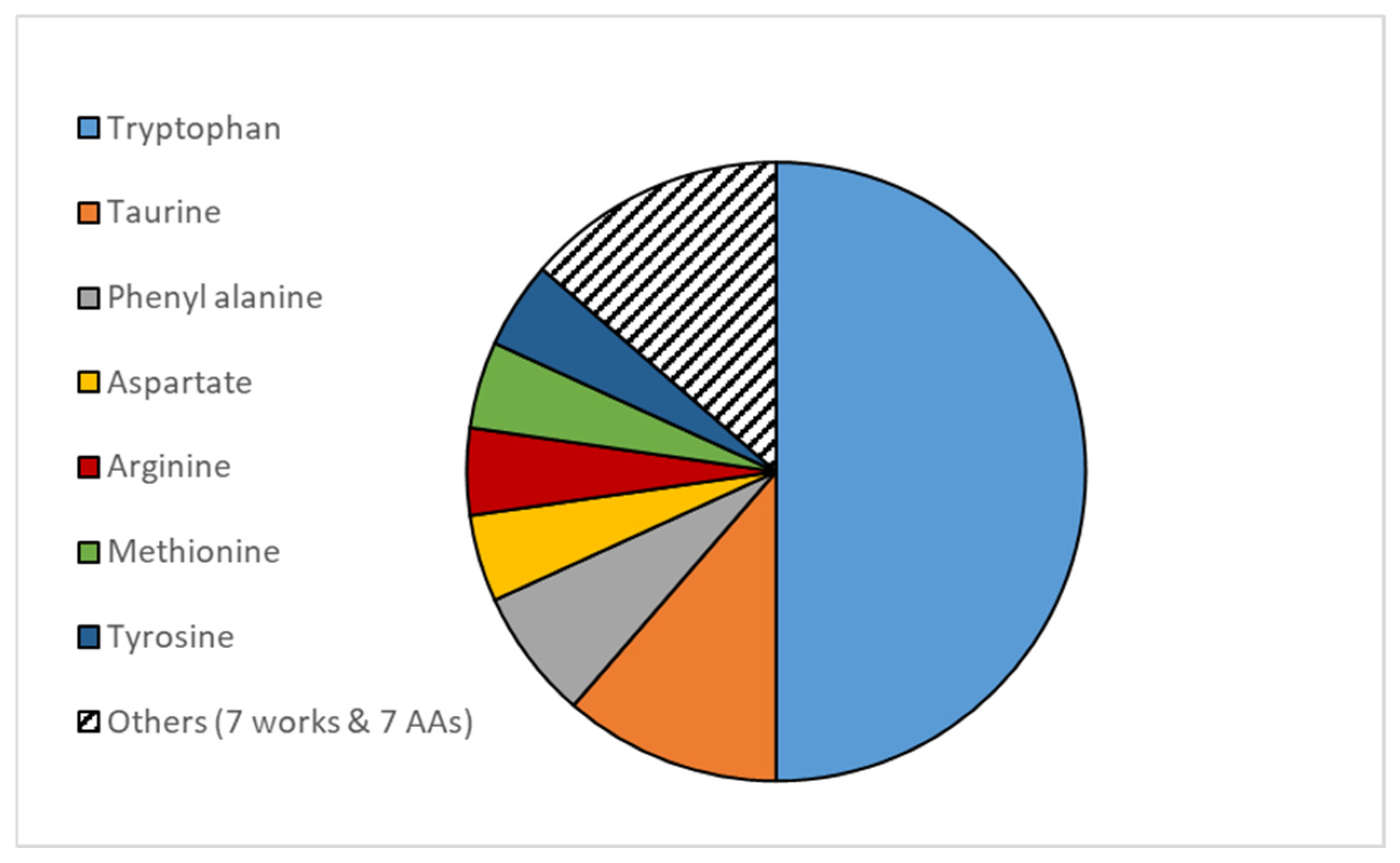

{kind=link}
| Amino Acid | Relevant Physiological Stress Responses | Species | References |
|---|---|---|---|
| Tryptophan | Improves feeding intake | Salmo trutta | Höglund et al. [35] |
| Reduces aggressiveness without affecting stress indicators | Brycon amazonicus | Wolkers et al. [39] | |
| Decreases growth Increases food consumption | Oncorhynchus mykiss | Papoutsoglou et al. [40] | |
| Increases growth parameters under stress conditions Decreases cortisol and glucose levels Decreases energy requirements | Cirrhinus mrigala | Tejpal et al. [41] | |
| Decrease cortisol, glucose levels and energy requirements | Argyrosomus regius | Teixeira et al. [42] | |
| Alters the prl and gh expressions after stress Increases Hsp70 expression | Argyrosomus regius | Herrera et al. [43] | |
| Decreases enzyme activities related to amino acid and carbohydrate metabolism Increases the liver kynurenine concentration | Argyrosomus regius | Herrera et al. [44] | |
| Decreases plasma lactate and mucus glucose | Argyrosomus regius | Fernández-Alacid et al. [45] | |
| Upregulates immune-related gene expressions | Argyrosomus regius | Asencio-Alcudia et al. [46] | |
| Keeps levels of protease, antiprotease, peroxidase and lysozyme unchanged | Argyrosomus regius | Gonzalez-Silvera et al. [47] | |
| Reduces SOD and CAT activities Increases serotonergic activity and plasma cortisol | Totoaba macdonaldi | Cabanillas-Gámez et al. [48] | |
| Decreased plasma cortisol levels Increased liver transaminase activity Raises enzyme activity in glycolysis and gluconeogenesis | Gadus morhua | Herrera et al. [49] | |
| Reduces plasma cortisol and lactate | Solea senegalensis | Salamanca et al. [50] | |
| Increases plasma cortisol Modulates plasma glucose and lactate | Solea senegalensis | Herrera et al. [18] | |
| Increases plasma cortisol Higher brain monoamine content | Dicentrarchus labrax | Azeredo et al. [51] | |
| Increases plasma cortisol Decreases plasma glucose Decreases lysozyme and ACH50 Decreases serum thyroid hormone Inhibits post-stress immunosuppression Decrease serum thyroid hormones. | Acipenser persicus | Hoseini et al. [52] | |
| Decreases aminotransferase and lactate dehydrogenase activities Reduces enzyme activities related to oxidative stress Higher growth, RGR and PER | Labeo rohita | Kumar et al. [53] | |
| Taurine | Improves growth performance, muscle composition and amino acid composition | Takifugu rubripes | Shi et al. [54] |
| Enhances growth performance Improves intestine structure | Mylopharyngodon piceus | Tian et al. [55] | |
| Increases CAT activity Increases total serum immunoglobulin concentration | Arapaima gigas | Souto et al. [37] | |
| Increases growth Decreases ROS production and antioxidant enzyme gene expressions | Dicentrarchus labrax | Ceccotti et al. [56] | |
| Phenylalanine | Lower plasma cortisol levels Increases liver transaminase activities Raises enzyme activity in glycolysis and gluconeogenesis | Gadus morhua | Herrera et al. [49] |
| Reduces plasma stress markers | Sparus aurata | Salamanca et al. [57] | |
| Reduces plasma glucose and lactate | Argyrosomus regius | Salamanca et al. [57] | |
| Aspartate | Enhances pomc-a expression Increases Hsp70 expression | Argyrosomus regius | Herrera et al. [43] |
| Produces over-exudation of mucus metabolites and cortisol | Argyrosomus regius | Fernández-Alacid et al. [45] | |
| Keeps protease, antiprotease, peroxidase and lysozyme levels stable | Argyrosomus regius | Gonzalez-Silvera et al. [47] | |
| Methionine | Decreases TG, TC, NEFA, LDL-C, and ALT Decreases lipid droplets in liver Increased ampkα and sirt1 expression Improves lipogenesis pathway gene expressions Up-regulates antioxidant enzyme activities and gene expression levels Decreases pro-inflammation and pro-apoptosis gene expressions Up-regulates anti-inflammatory cytokine and anti-apoptosis gene expressions | Acanthopagrus schlegelii | Yang et al. [36] |
| Increases plasma cortisol Upregulates complement factor 3 Increases immune cells | Dicentrarchus labrax | Azeredo et al. [51] | |
| Tyrosine | Reduces plasma stress markers | Sparus aurata | Salamanca et al. [57] |
| Arginine | Decreases plasma cortisol levels | Scophthalmus maximus | Costas et al. [34] |
| Increases respiratory burst activity and nitric oxide production of head kidney leukocytes Enhances HIF-1, HAMP-1, MIP1-alpha and gLYS expressions | Solea senegalensis | Costas et al. [58] | |
| Alanine + Glutamine | Increases body weight Increases fish survival during a bacterial challenge | Cyprinus carpio | Chen et al. [59] |
| Leucine | Promotes FW, WG, PWG, and SGR Decreases activities of serum parameters Decreases ROS, NO and ONOO− activities Increases mRNA levels of mitochondrial biogenesis genes and fusion-related genes Decreases mRNA levels of fission-related genes, mitophagy-related genes and autophagy-related genes | Ctenopharyngodon idella | Zhen et al. [60] |
| Valine + Isoleucine | Enhances growth Increases blood parameter levels | Paralichthys olivaceus | Shi et al. [61] |
| Gamma-aminobutyric acid | Improves macrophage maturation, autophagy activation, and antibacterial response to bacterial infection | Paralichthys olivaceus | Bae et al. [62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salamanca, N.; Herrera, M.; de la Roca, E. Amino Acids as Dietary Additives for Enhancing Fish Welfare in Aquaculture. Animals 2025, 15, 1293. https://doi.org/10.3390/ani15091293
Salamanca N, Herrera M, de la Roca E. Amino Acids as Dietary Additives for Enhancing Fish Welfare in Aquaculture. Animals. 2025; 15(9):1293. https://doi.org/10.3390/ani15091293
Chicago/Turabian StyleSalamanca, Natalia, Marcelino Herrera, and Elena de la Roca. 2025. "Amino Acids as Dietary Additives for Enhancing Fish Welfare in Aquaculture" Animals 15, no. 9: 1293. https://doi.org/10.3390/ani15091293
APA StyleSalamanca, N., Herrera, M., & de la Roca, E. (2025). Amino Acids as Dietary Additives for Enhancing Fish Welfare in Aquaculture. Animals, 15(9), 1293. https://doi.org/10.3390/ani15091293