
Food Restriction Induces Changes in Ovarian Folliculogenesis, Cell Proliferation, Apoptosis, and Production of Regulatory Peptides in Rabbits
 ,
,  , ,
, ,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
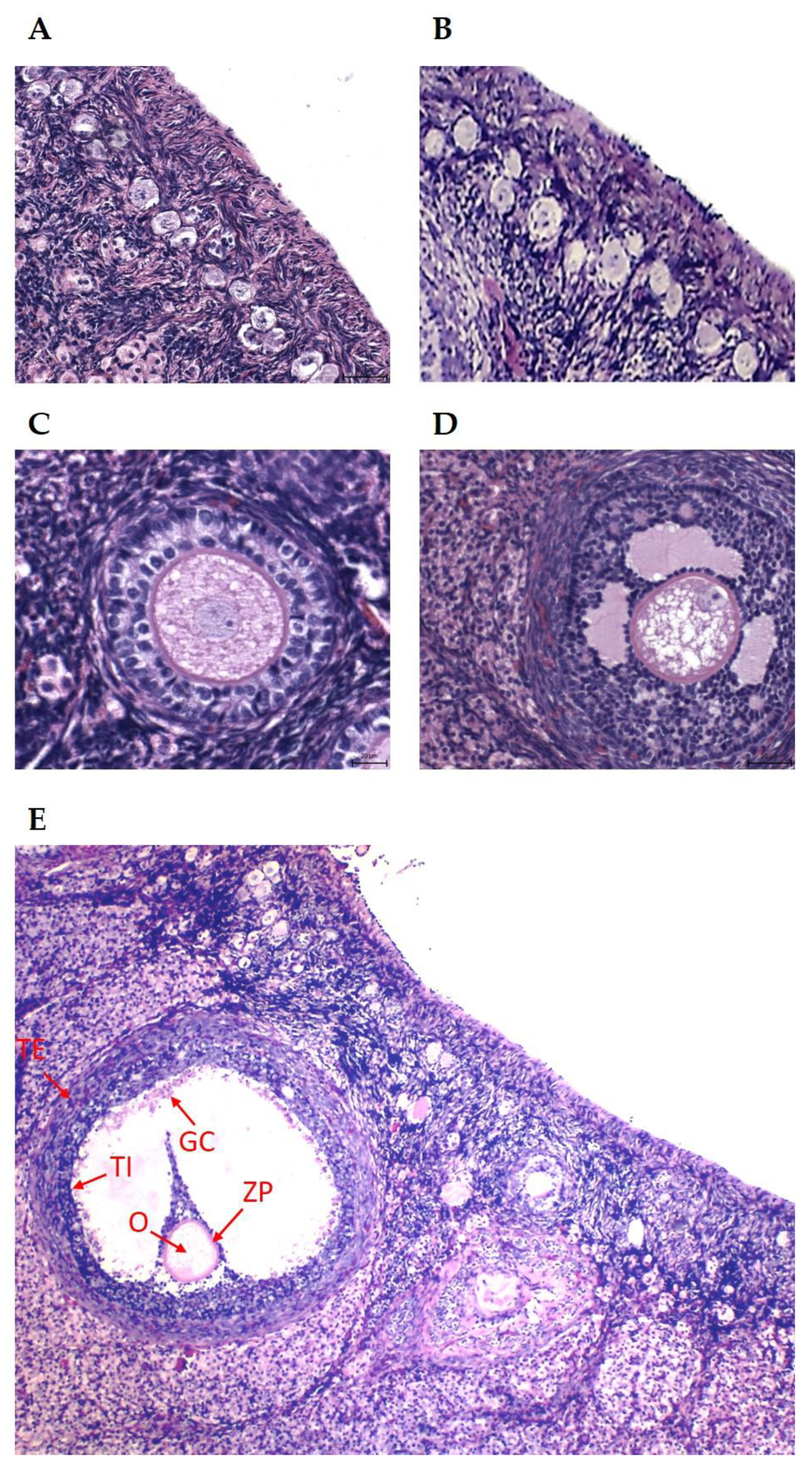
2.2. Histological Processing and Analysis of Ovaries
2.3. Preparation, Culture, and Processing of Ovarian Granulosa Cells
2.4. Preparation, Culture, and Processing of Follicular Fragments
2.5. Cell Viability Test
2.6. BrdU Assay
2.7. Analysis of Nuclear Apoptosis
2.8. Analysis of Proliferation and Cytoplasmic Apoptosis Markers
2.9. Immunoassay of Hormones
2.10. Sample Preparation for Proteomic Evaluation (Profiling)
2.11. Protein Identification by Tandem Mass Spectrometry (Nano HPLC-Chip-MS/MS)
2.12. Gene Ontology Enrichment Analysis
2.13. Statistical Analysis
3. Results
3.1. Fecundity and Anatomical Reproductive Traits of Does Subjected or Not Subjected to Food Restriction
3.2. Histomorphometric Traits in Ovaries of Does Subjected to Normal or Restricted Feeding
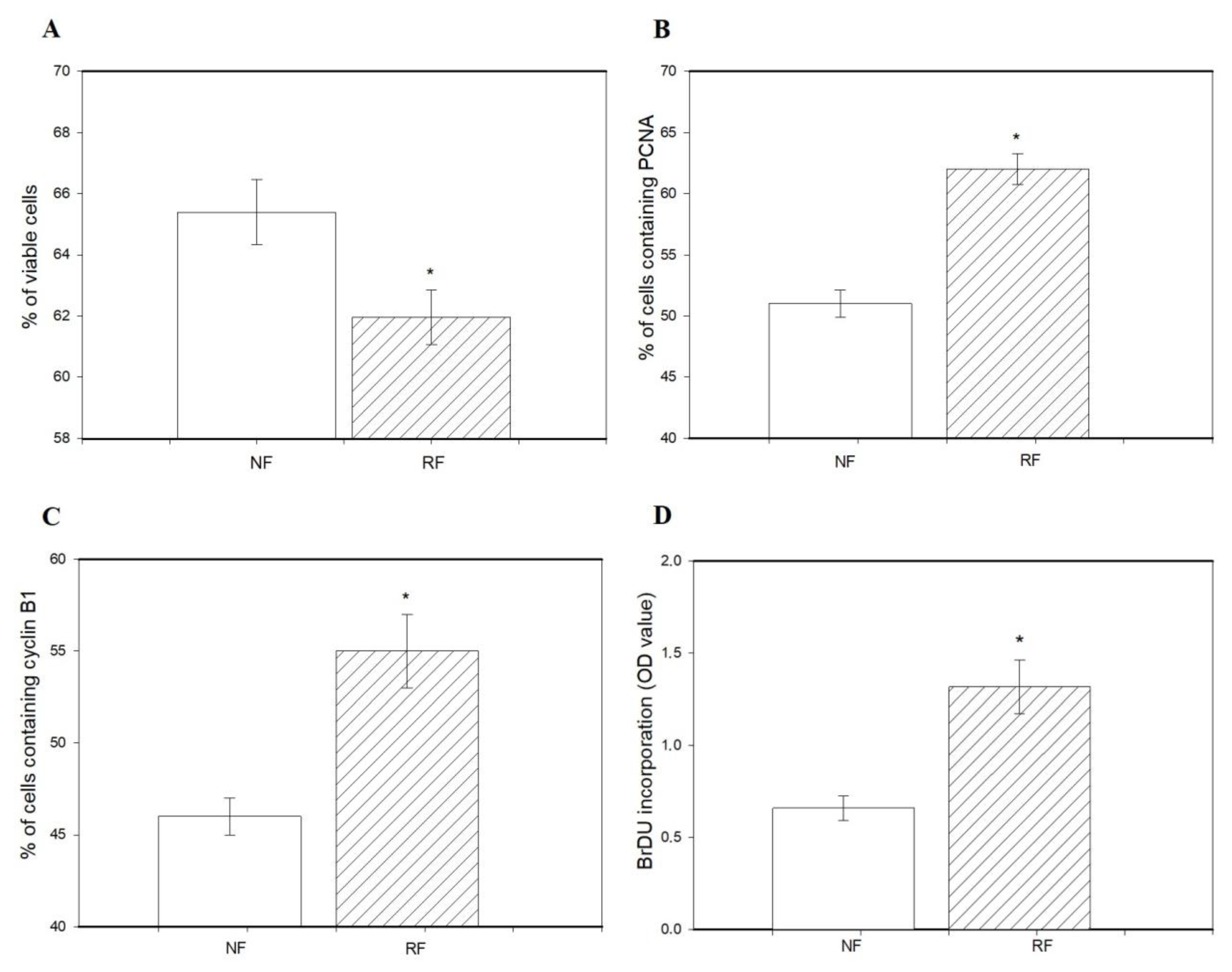
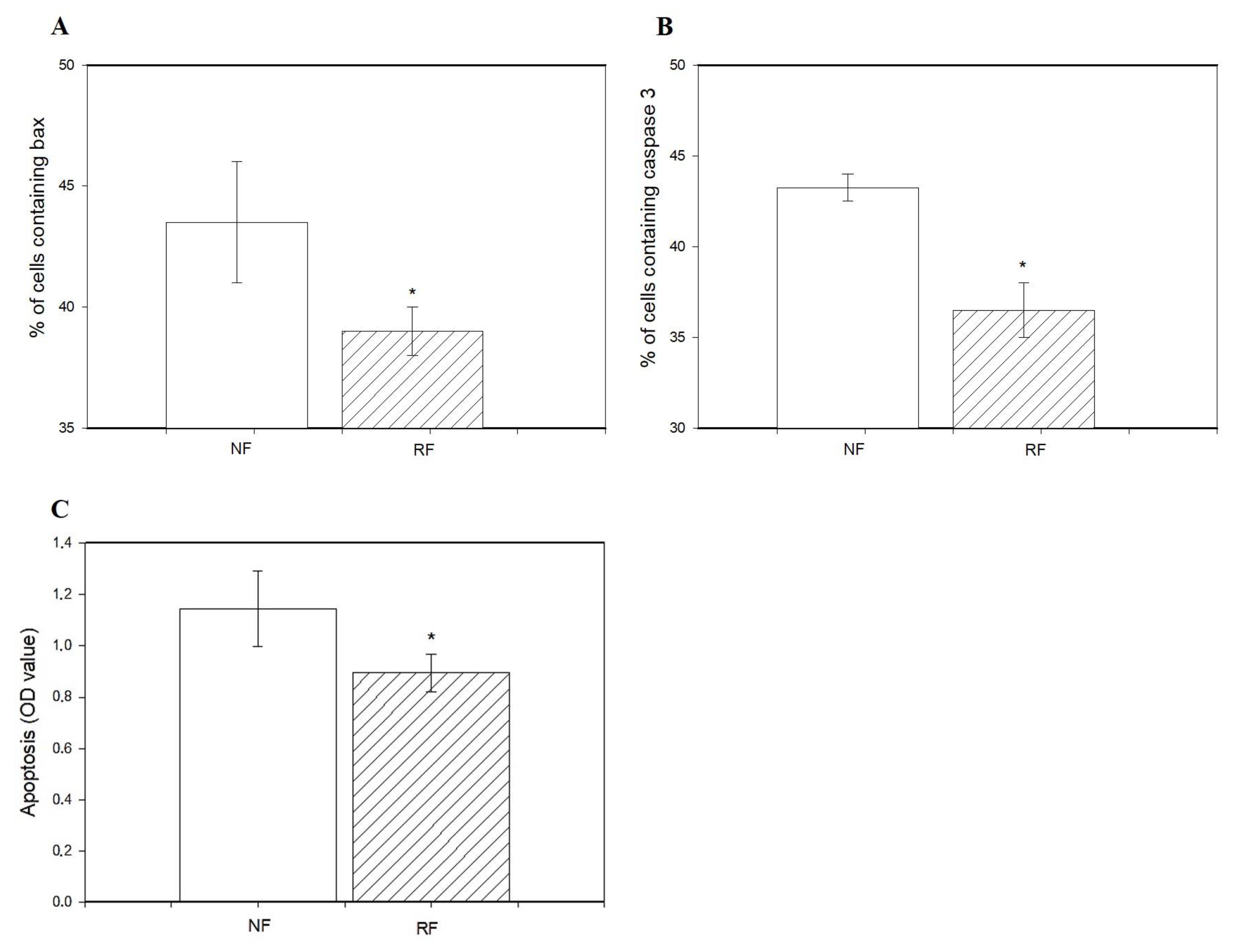
3.3. Viability, Proliferation, Apoptosis, and Release of Hormones by Ovarian Cells and Fragments of Does Subjected to Nonrestricted or Restricted Feeding
3.4. Proteomic Analysis of Ovarian Cells of Does Subjected to Normal or Restricted Feeding
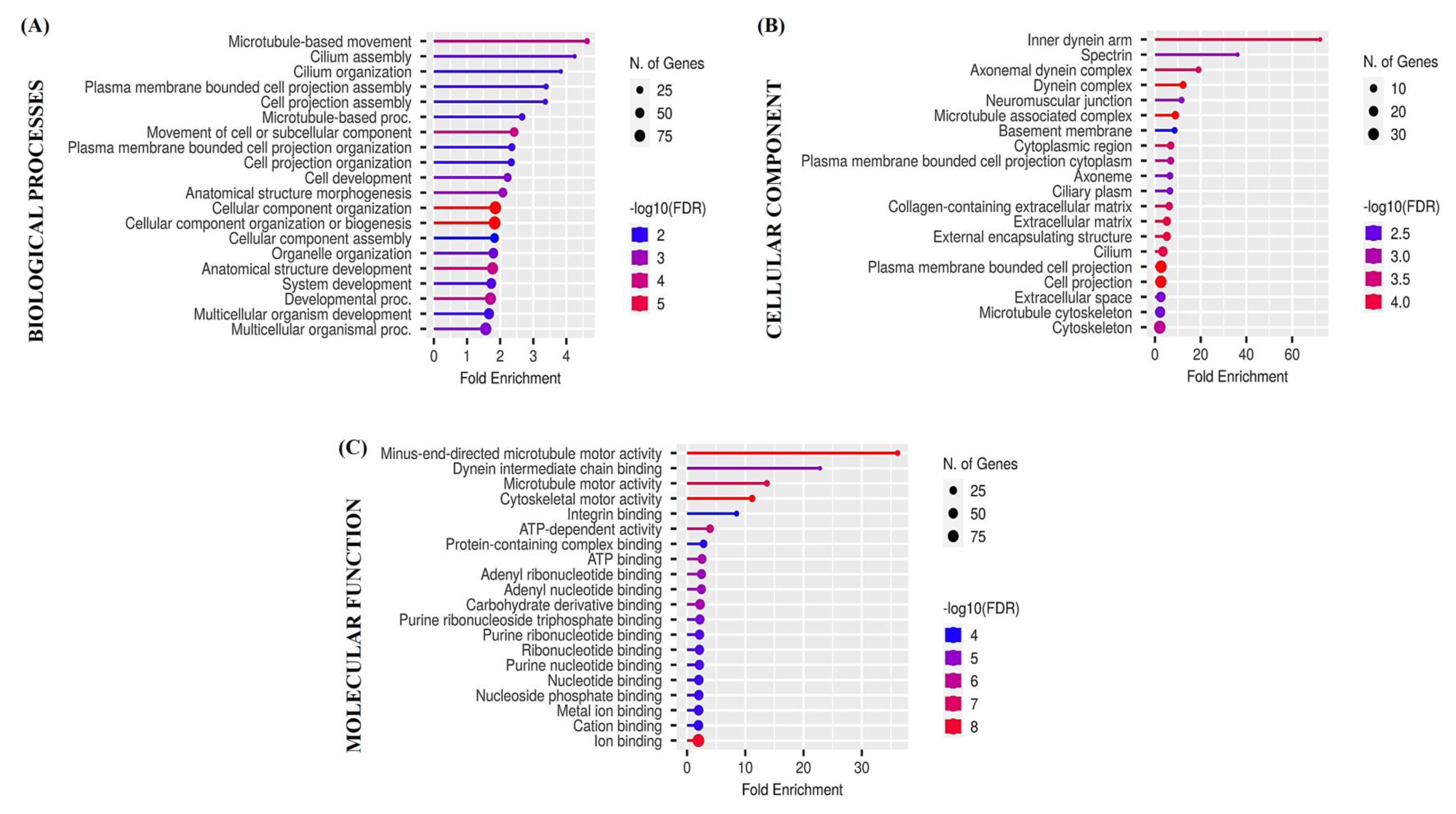
3.5. Functional Enrichment Analysis
4. Discussion
4.1. Anatomical Characterisation of the Reproductive Tract of Females Under Food Restriction
4.2. Effect of Food Restriction on Folliculogenesis and Oogenesis in the Ovaries
4.3. Effect of Food Restriction on Proliferation, Apoptosis, and Hormone Release by Ovarian Cells
4.4. Effect of Food Restriction on Proteomic Profile
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NF | Females fed ad libitum |
| RF | Females subjected to 50% food restriction |
| DAB | 3.3′-diaminobenzidine substrate |
References
- Boland, M.P.; Lonergan, P.; O’Callaghan, D. Effect of nutrition on endocrine parameters, ovarian physiology and oocyte and embryo development. Theriogenology 2001, 55, 1323–1340. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, E.M.; Ashworth, C.J.; Edwards, S.A.; Hawkins, N.; Hepburn, N.; Hunter, M.G. Effect of different nutritional regimens before ovulation on plasma concentrations of metabolic and reproductive hormones and oocyte maturation in gilts. Reproduction 2003, 126, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Kiyma, Z.; Alexander, B.M.; Van Krik, E.A.; Murdoch, W.J.; Hallford, D.M.; Moss, G.E. Effects of feed restriction on reproductive and metabolic hormones in ewes. J. Anim. Sci. 2004, 82, 2548–2557. [Google Scholar] [CrossRef] [PubMed]
- Brecchia, G.; Bonanno, A.; Galeati, G.; Federici, C.; Maranesi, M.; Gobbetti, A.; Boiti, C. Hormonal and metabolic adaptation to fasting: Effects on the hypothalamic-pituitary-ovarian axis and reproductive performance of rabbit does. Domest. Anim. Endocrinol. 2006, 31, 105–122. [Google Scholar] [CrossRef]
- Armstrong, D.G.; McEvoy, T.G.; Baxter, G.; Robinson, J.J.; Hogg, C.O.; Woad, K.J.; Webb, R.; Sinclair, K.D. Effect of dietary energy and protein on bovine follicular dynamics and embryo production in vitro: Associations with the ovarian insulin-like growth factor system. Biol. Reprod. 2001, 64, 1624–1632. [Google Scholar] [CrossRef]
- Alexander, B.M.; Kiyma, Z.; McFarland, M.; Van Krik, E.A.; Hallford, D.M.; Hawkins, D.E.; Kane, K.K.; Moss, G.E. Influence of short-term fasting during the luteal phase of the oestrous cycle on ovarian follicular development during the ensuing proestrus of ewes. Anim. Reprod. Sci. 2007, 97, 356–363. [Google Scholar] [CrossRef]
- Szendrő, Z.; Szendrő, K.; Zotte, A.D. Management of reproduction on small, medium and large rabbit farms: A review. Asian Australas. J. Anim. Sci. 2012, 25, 738–748. [Google Scholar] [CrossRef]
- Manal, A.F.; Tony, M.A.; Ezzo, O.H. Feed restriction of pregnant nulliparous rabbit does: Consequences on reproductive performance and maternal behaviour. Anim. Reprod. Sci. 2010, 120, 179–186. [Google Scholar] [CrossRef]
- García-García, R.M.; Rebollar, P.G.; Arias-Alvarez, M.; Sakr, O.G.; Bermejo-Alvarez, P.; Brecchia, G.; Gutierres-Adan, A.; Zerani, M.; Boiti, C.; Lorenzo, P.L. Acute fasting before conception affects metabolic and endocrine status without impacting follicle and oocyte development and embryo gene expression in the rabbit. Reprod. Fertil. Dev. 2011, 23, 759–768. [Google Scholar] [CrossRef]
- Menchetti, L.; Brecchia, G.; Canali, C.; Cardinali, R.; Polisca, A.; Zerani, M.; Boiti, C. Food restriction during pregnancy in rabbits: Effects on hormones and metabolites involved in energy homeostasis and metabolic programming. Res. Vet. Sci. 2015, 98, 7–12. [Google Scholar] [CrossRef]
- Fortun-Lamothe, L. Effects of pre-mating energy intake on reproductive performance of rabbit does. Anim. Sci. 1998, 66, 263–269. [Google Scholar] [CrossRef]
- Daoud, N.M.; Mahrous, K.F.; Ezzo, O.H. Feed restriction as a biostimulant of the production of oocyte, their quality and GDF-9 gene expression in rabbit oocytes. Anim. Reprod. Sci. 2012, 136, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Naturil-Alfonso, C.; Lavara, R.; Vicente, J.S.; Marco-Jimenez, F. Effects of female dietary restriction in a rabbit growth line during rearing on reproductive performance and embryo quality. Reprod. Domest. Anim. 2016, 51, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V.; Koničková, I.; Østrup, O.; Rafay, J.; Laurincik, J.; Harrath, A.H. Caloric restriction and IGF-I administration promote rabbit fecundity: Possible interrelationships and mechanisms of action. Theriogenology 2017, 90, 252–259. [Google Scholar] [CrossRef]
- Harrath, A.H.; Østrup, O.; Rafay, J.; Koničková Florkovičová, I.; Laurincik, J.; Sirotkin, A.V. Metabolic state defines the response of rabbit ovarian cells to leptin. Repro. Biol. 2017, 17, 19–24. [Google Scholar] [CrossRef]
- Blasco, A.; Martínez-Álvaro, M.; García, M.L.; Ibáñez-Escriche, N.; Argente, M.J. Selection for genetic environmental sensitivity of litter size in rabbits. Genet. Sel. Evol. 2017, 49, 48–55. [Google Scholar] [CrossRef]
- Pedersen, T.; Peters, H. Proposal for a classification of oocytes and follicles in the mouse ovary. J. Reprod. Fertil. 1968, 17, 555–557. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Pavlova, S.; Tena-Sempere, M.; Grossmann, R.; Jiménez, M.R.; Rodriguez, J.M.; Valenzuela, F. Food restriction, ghrelin, its antagonist and obestatin control expression of ghrelin and its receptor in chicken hypothalamus and ovary. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 164, 141–153. [Google Scholar] [CrossRef]
- Mescher, A.L. Junqueira’s Basic Histology: Text and Atlas, 16th ed.; McGraw-Hill: New York, NY, USA, 2021. [Google Scholar]
- Perry, S.W.; Epstein, L.G.; Gelbard, H.A. In situ trypan blue staining of monolayer cell cultures for permanent fixation and mounting. BioTechniques 1997, 22, 1020–1024. [Google Scholar] [CrossRef]
- Uzuner, S.Ç. Development of a direct trypan blue exclusion method to detect cell viability of adherent cells into ELISA plates. Celal Bayar Univ. Fen Bilim. Derg. 2018, 14, 99–104. [Google Scholar]
- Fabová, Z.; Loncová, B.; Mlynček, M.; Sirotkin, A.V. Interrelationships between amphiregulin, kisspeptin, FSH and FSH receptor in promotion of human ovarian cell functions. Reprod. Fertil. Dev. 2022, 34, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Jin, F.; Wu, W.; Kumar, S.K. Cell cycle regulation and hematologic malignancies. Blood Sci. 2019, 1, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Ligasová, A.; Frydrych, I.; Koberna, K. Basic Methods of Cell Cycle Analysis. Int. J. Mol. Sci. 2023, 24, 3674. [Google Scholar] [CrossRef]
- Spitz, A.Z.; Gavathiotis, E. Physiological and pharmacological modulation of BAX. Trends Pharmacol. Sci. 2022, 43, 206–220. [Google Scholar] [CrossRef]
- Sirotkin, A.V. Regulators of Ovarian Functions; Nova Science Publishers: New York, NY, USA, 2014; p. 194. [Google Scholar]
- Chou, C.H.; Chen, M.J. The effect of steroid hormones on ovarian follicle development. Vitam. Horm. 2018, 107, 155–175. [Google Scholar] [CrossRef]
- Chen, P.; Li, B.; Ou-Yang, L. Role of estrogen receptors in health and disease. Front. Endocrinol. 2022, 18, 839005. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- García, M.L.; Muelas, R.; Argente, M.J.; Peiró, R. Relationship between prenatal characteristics and body condition and endocrine profile in Rabbits. Animals 2021, 11, 95. [Google Scholar] [CrossRef]
- Rebollar, P.G.; Pérez-Cabal, M.A.; Pereda, N.; Lorenzo, P.L.; Arias-Álvarez, M.; García-Rebollar, P. Effects of parity order and reproductive management on the efficiency of rabbit productive systems. Livest. Scie. 2009, 121, 227–233. [Google Scholar] [CrossRef]
- Bolet, G.; Garreau, H.; Joly, T.; Theau-Clement, M.; Falieres, J.; Hurtaud, J.; Bodin, L. Genetic homogenisation of birth weight in rabbits: Indirect selection response for uterine horn characteristics. Livest. Scie. 2007, 111, 28–32. [Google Scholar] [CrossRef]
- Argente, M.J.; Santacreu, M.A.; Climent, A.; Blasco, A. Influence of available uterine space per fetus on fetal development and prenatal survival in rabbits selected for uterine capacity. Livest. Scie. 2006, 102, 83–91. [Google Scholar] [CrossRef]
- Žitný, J.; Massányi, P.; Trakovická, A.; Rafaj, J.; Toman, R. Quantification of the ovarian follicular growth in rabbits. Bull. Vet. Inst. Pulawy 2004, 48, 37–40. [Google Scholar]
- Al-Mufti, W.; Bomsel-Helmreich, O.; Chritidès, J.P. Oocyte size and intrafollicular position in polyovular follicles in rabbits. J. Reprod. Fert. 1988, 82, 15–25. [Google Scholar] [CrossRef]
- Naseer, Z.; Ahmad, E.; Epikmen, E.T.; Uçan, U.; Boyacioğlu, M.; İpek, E.; Akosy, M. Quercetin supplemented diet improves follicular development, oocyte quality, and reduces ovarian apoptosis in rabbits during summer heat stress. Theriogenology 2017, 96, 136–141. [Google Scholar] [CrossRef]
- Hutt, K.J.; McLaughlin, E.A.; Holland, M.K. Primordial follicle activation and follicular development in the juvenile rabbit ovary. Cell Tissue Res. 2006, 326, 809–822. [Google Scholar] [CrossRef]
- Alam, M.H.; Miyano, T. Interaction between growing oocytes and granulosa cells in vitro. Reprod. Med. Biol. 2020, 19, 13–23. [Google Scholar] [CrossRef]
- Martí-Clúa, J. Incorporation of 5-Bromo-2′-deoxyuridine into DNA and Proliferative Behavior of Cerebellar Neuroblasts: All That Glitters Is Not Gold. Cells 2021, 10, 1453. [Google Scholar] [CrossRef]
- McCann, J.P.; Hansel, W. Relationship between insulin and glucose metabolism and pituitary-ovarian functions in fasted heifers. Biol. Reprod. 1986, 34, 630–641. [Google Scholar] [CrossRef]
- Dunkley, S.; Scheffler, K.; Mogessie, B. Cytoskeletal form and function in mammalian oocytes and zygotes. Curr. Opin. Cell Biol. 2022, 75, 102073. [Google Scholar] [CrossRef] [PubMed]
- Pathirana, A.; Diao, M.; Huang, S.; Zuo, L.; Liang, Y. Alpha 2 macroglobulin is a maternally-derived immune factor in amphioxus embryos: New evidence for defense roles of maternal immune components in invertebrate chordate. Fish Shellfish Immunol. 2016, 50, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Saadeldin, I.M.; Tukur, H.A.; Aljumaah, R.S.; Sindi, R.A. Rocking the boat: The decisive roles of rho kinases during oocyte, blastocyst, and stem cell development. Front. Cell Dev. Biol. 2021, 8, 616762. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance Assayed | Specificity of Assay (Cross-Reactivity of Antiserum) | Sensitivity of Assay (ng/mL) | Coefficient of Variation (%) | |
|---|---|---|---|---|
| Intra-Assay | Inter-Assay | |||
| Progesterone | ≤1.1% with 11-desoxycorticosterone, ≤0.35% with pregnenolone, ≤0.30% 17α-OH with progesterone, ≤0.20% with corticosterone, ˂0.10% with estriol, 17β-estradiol, testosterone, cortisone, and 11-desoxycortisol, ˂0.02% with DHEA-S and cortisol | 0.045 | 5.4 | 5.6 |
| 17β-estradiol | ≤9.5% with fulvestrant, ≤4.2% with estrone, ≤3.8% with E2-3-glucuronide, ≤3.6% with E2-3-sulphate, ≤0.4% with estriol, ˂0.1% with androstenedione, 17-hydroxyprogesterone, corticosterone, pregnenolone, E2-17-glucuronide, progesterone, and testosterone | 0.0062 | 6.4 | 4.5 |
| Traits | Initial (n = 8) | Final (n = 8) | p | |
|---|---|---|---|---|
| Female weight | NF (g) | 3532 ± 171 | 3766 ± 175 | ns |
| RF (g) | 3349 ± 125 | 3190 ± 104 | ns |
| Traits | NF (n = 8) | RF (n =8) | p | |
|---|---|---|---|---|
| Female weight | Initial (g) | 3532 ± 171 | 3349 ± 125 | ns |
| Final (g) | 3766 ± 175 | 3190 ± 104 | ** | |
| Ovary weight | Left (g) | 0.36 ± 0.05 | 0.25 ± 0.03 | * |
| Right (g) | 0.40 ± 0.05 | 0.27 ± 0.03 | * | |
| Uterine horn length | Left (cm) | 6.56 ± 0.30 | 8.40 ± 0.66 | * |
| Right (cm) | 6.56 ± 0.43 | 8.24 ± 0.52 | * | |
| Uterine horn weight | Left (g) | 5.09 ± 0.35 | 2.90 ± 0.37 | *** |
| Right (g) | 4.92 ± 0.44 | 3.08 ± 0.40 | ** |
| Ovarian Structures and Their Parameters | Feeding | |
|---|---|---|
| NF | RF | |
| Dormant primordial follicles: | ||
| Number | 126 | 176 |
| Proportion concerning all detected follicles (%) | 45 | 48.2 |
| Diameter of follicles (µm) | 30.59 ± 0.45 | 32.01 ± 0.36 * |
| Diameter of oocytes (µm) | 23.21 ± 0.36 | 24.54 ± 0.35 |
| % of degenerated follicles | 0 | 0 |
| Growing primordial follicles: | ||
| Number | 64 | 70 |
| Proportion concerning all detected follicles (%) | 22.9 | 19.18 |
| Diameter of follicles (µm) | 55.62 ± 1.72 | 58.69 ± 1.89 |
| Diameter of oocytes (µm) | 35.44 ± 1.28 | 36.24 ± 1.27 |
| % of degenerated follicles | 18.8 | 20.0 |
| Primary (preantral) follicles: | ||
| Number | 52 | 74 |
| Proportion concerning all detected follicles (%) | 18.57 | 20.27 |
| Diameter of follicles (µm) | 182.27 ± 7.54 | 170.72 ± 5.57 |
| Thickness of theca (µm) | 17.62 ± 0.54 | 16.51 ± 1.06 |
| Thickness of granulosa (µm) | 31.65 ± 1.06 | 34.56 ± 1.82 * |
| Diameter of oocytes (µm) | 84.90 ± 2.70 | 103.14 ± 4.91 * |
| % of degenerated follicles | 7.7 | 25.7 * |
| Secondary (antral) follicles: | ||
| Number | 36 | 39 |
| Proportion concerning all detected follicles (%) | 12.86 | 10.68 |
| Diameter of follicles (µm) | 403.95 ± 15.36 | 460.75 ± 31.09 * |
| Thickness of theca (µm) | 41.76 ± 1.56 | 45.61 ± 1.11 * |
| Thickness of granulosa (µm) | 110.80 ± 1.71 | 127.65 ± 0.84 * |
| Diameter of oocytes (µm) | 103.02 ± 3.81 | 114.25 ± 2.49 * |
| % of degenerated follicles | 27.8 | 30.8 |
| Preovulatory follicles: | ||
| Number | 2 | 6 |
| Proportion concerning all detected follicles (%) | 0.71 | 1.64 * |
| Diameter of follicles (µm) | 429.53 ± 63.47 | 824.78 ± 101.56 * |
| Thickness of theca (µm) | 33.27 ± 1.60 | 58.15 ± 3.10 * |
| Thickness of granulosa (µm) | 124.52 ± 6.12 | 294.79 ± 35.94 * |
| Diameter of oocytes (µm) | 113.93 ± 11.12 | 118.89 ± 3.26 |
| % of degenerated follicles | 0 | 0 |
| Protein Function | Accession Number * | Protein Name | Feeding | |
|---|---|---|---|---|
| NF | RF | |||
| Cell differentiation | G1SDQ4 | Rho guanine nucleotide exchange factor 28 | - | + |
| Cell growth | G1SD01 G1TV13 A0A5F9CH25 G1T8R3 U3KPK3 A0A5F9C3V6 A0A5F9CT68 G1SU35 A0A5F9CHT0 B7NZQ3 A0A5F9C3J9 | Spectrin beta chain Dynein axonemal heavy chain 2 Dynein axonemal heavy chain 17 Utrophin Diaphanous related formin 2 Myosin XVB Cytoskeleton associated protein 5 Dynein axonemal heavy chain 1 Ubiquitinyl hydrolase 1 Deoxyribonuclease Cell migration inducing hyaluronidase 1 | - - - - - + + + - - - | + + + + + - - -+ + + + |
| Transporter activity | G1SLC9 | Sodium/hydrogen exchanger | + | - |
| Extracellular matrix organization | A0A5F9C8H0 | ADAM metallopeptidase with thrombospondin type 1 motif 9 | + | - |
| Programmed cell death | Q27Q52 | Protein Wnt-5a | + | - |
| Mitotic cell cycle | A0A5F9C8V0 | Tetratricopeptide repeat domain 28 | - | + |
| GTP-ase activity | A0A5F9CV79 G1STZ5 G1U5G0 | Neurofibromin 1 Nonspecific serine/threonine protein kinase Rho guanine nucleotide exchange factor 33 | - - + | + + - |
| Antioxidant activity | A0A5F9DGD9 | Peroxidasin like | - | + |
| Regulation of DNA-templated transcription | A0A5F9CNT4 A0A5F9C9Q6 | GLI family zinc finger 3 Zinc finger protein 7 | - + | + - |
| Carbohydrate metabolic processes | A0A5F9CRS6 | Otogelin like | - | + |
| Protease inhibition | A0A5F9C8I9 | Alpha-2-macroglobulin | - | + |
| Chromatin organization | A0A5F9DIU5 | Jumonji domain containing 1C | + | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadjadj, I.; Fabova, Z.; García, M.-L.; Agea, I.; Loncová, B.; Morovic, M.; Makovicky, P.; Argente, M.-J.; Sirotkin, A.V. Food Restriction Induces Changes in Ovarian Folliculogenesis, Cell Proliferation, Apoptosis, and Production of Regulatory Peptides in Rabbits. Animals 2025, 15, 1282. https://doi.org/10.3390/ani15091282
Hadjadj I, Fabova Z, García M-L, Agea I, Loncová B, Morovic M, Makovicky P, Argente M-J, Sirotkin AV. Food Restriction Induces Changes in Ovarian Folliculogenesis, Cell Proliferation, Apoptosis, and Production of Regulatory Peptides in Rabbits. Animals. 2025; 15(9):1282. https://doi.org/10.3390/ani15091282
Chicago/Turabian StyleHadjadj, Imane, Zuzana Fabova, María-Luz García, Iván Agea, Barbora Loncová, Martin Morovic, Peter Makovicky, María-José Argente, and Alexander V. Sirotkin. 2025. "Food Restriction Induces Changes in Ovarian Folliculogenesis, Cell Proliferation, Apoptosis, and Production of Regulatory Peptides in Rabbits" Animals 15, no. 9: 1282. https://doi.org/10.3390/ani15091282
APA StyleHadjadj, I., Fabova, Z., García, M.-L., Agea, I., Loncová, B., Morovic, M., Makovicky, P., Argente, M.-J., & Sirotkin, A. V. (2025). Food Restriction Induces Changes in Ovarian Folliculogenesis, Cell Proliferation, Apoptosis, and Production of Regulatory Peptides in Rabbits. Animals, 15(9), 1282. https://doi.org/10.3390/ani15091282