
Genome-Wide Association Study for Belly Traits in Canadian Commercial Crossbred Pigs
 , , , ,
, , , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Phenotypes
2.2. Genotypes
2.3. Statistical Analysis
Sample Size Justification Based on Power Analysis
2.4. Post-GWAS Analyses
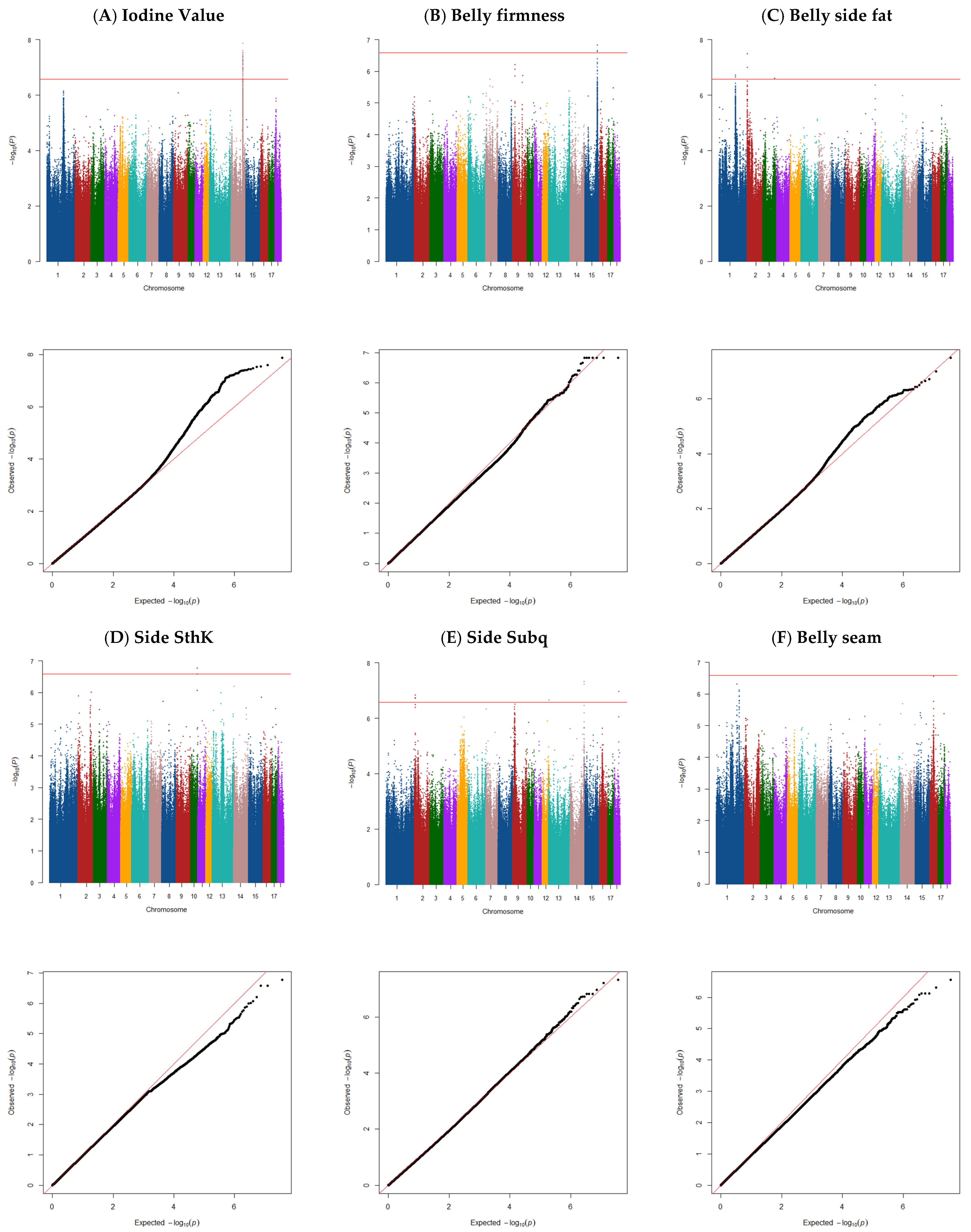
3. Results and Discussion
3.1. Descriptive Statistics and Heritability Estimates
3.2. Iodine Value
3.3. Belly Firmness
3.4. Belly Side Fat Thickness
3.5. Total Side Thickness
3.6. Side Subcutaneous Thickness
3.7. Belly Seam Fat Thickness
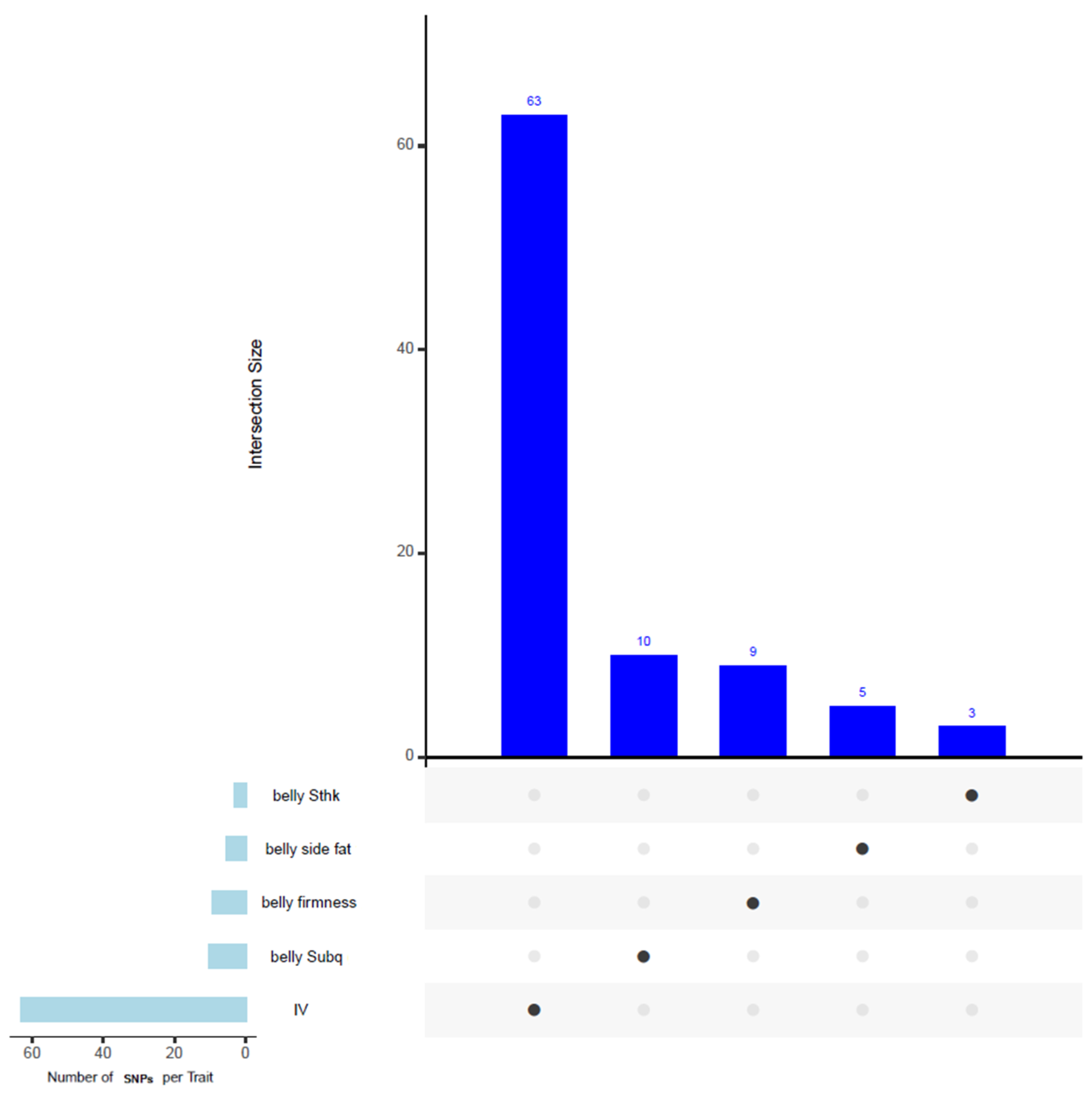
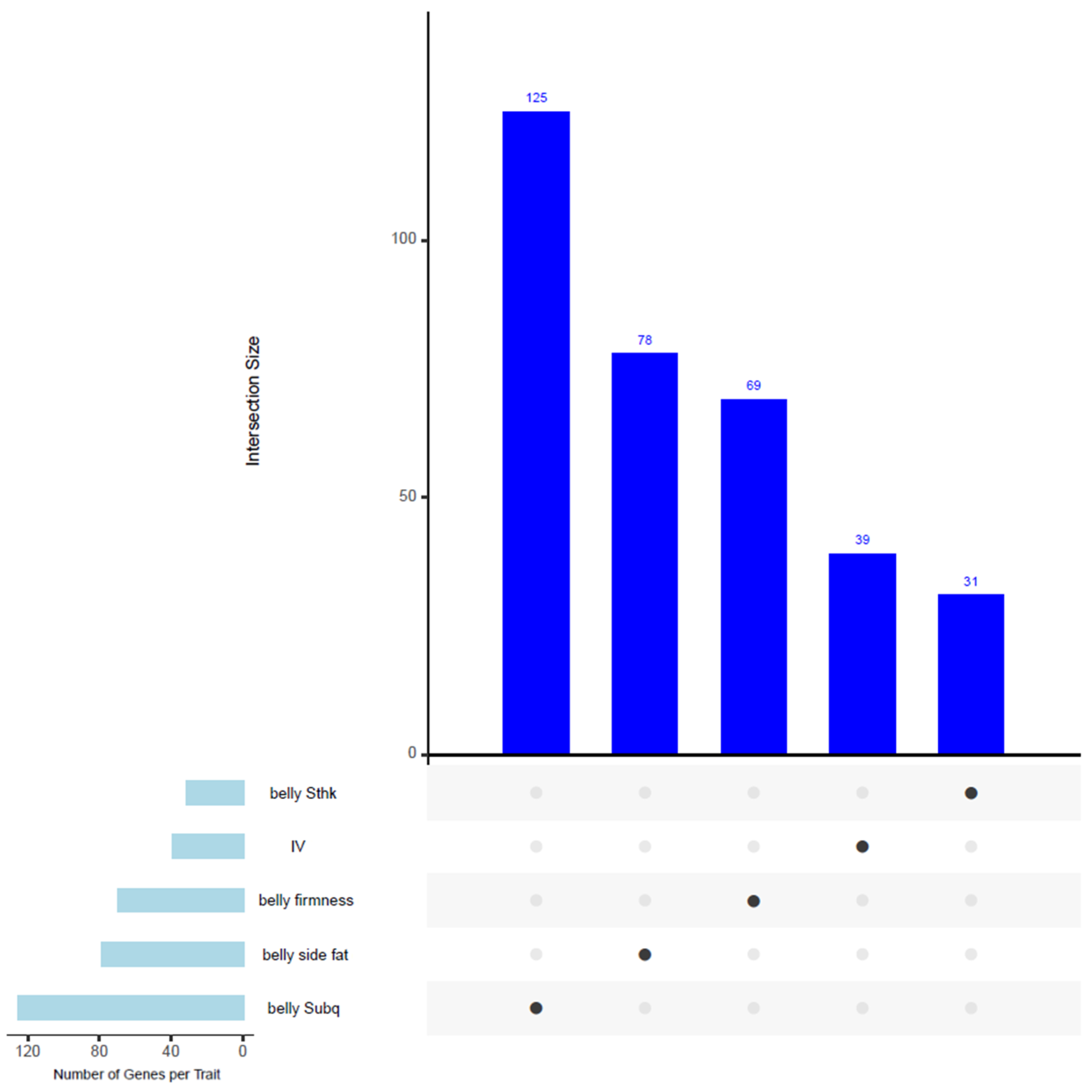
3.8. Common SNPs and Genes in Windows Associated with Multiple Traits
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.H.; Kim, J.M. Breeding Potential for Pork Belly to the Novel Economic Trait. J. Anim. Sci. Technol. 2023, 65, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhou, Q.; Lin, C.; He, L.; Wei, L. Integrative Analyses of Gene Expression and Alternative Splicing to Gain Insights into the Effects of Copper on Hepatic Lipid Metabolism in Swamp Eel (Monopterus albus). Aquaculture 2022, 546, 737367. [Google Scholar] [CrossRef]
- Arkfeld, E.K.; Wilson, K.B.; Overholt, M.F.; Harsh, B.N.; Lowell, J.E.; Hogan, E.K.; Klehm, B.J.; Bohrer, B.M.; Mohrhauser, D.A.; King, D.A.; et al. Pork loin quality is not indicative of fresh belly or fresh and cured ham quality. J. Anim. Sci. 2016, 94, 5155–5167. [Google Scholar] [CrossRef] [PubMed]
- Person, R.C.; McKenna, D.R.; Griffin, D.B.; McKeith, F.K.; Scanga, J.A.; Belk, K.E.; Smith, G.C.; Savell, J.W. Benchmarking Value in the Pork Supply Chain: Processing Characteristics and Consumer Evaluations of Pork Bellies of Different Thicknesses When Manufactured into Bacon. Meat Sci. 2005, 70, 121–131. [Google Scholar] [CrossRef]
- Juárez, M.; Plett, D.; Uttaro, B. Manually Operated Portable Equipment for Measurement and Classification of Fresh Pork Belly Firmness. Invent. Discl. 2023, 3, 100014. [Google Scholar] [CrossRef]
- Knecht, D.; Duziński, K.; Jankowska-Mąkosa, A. Variability of Fresh Pork Belly Quality Evaluation Results Depends on Measurement Locations. Food Anal. Methods 2018, 11, 2195–2205. [Google Scholar] [CrossRef]
- Soladoye, P.O.; Shand, P.J.; Aalhus, J.L.; Gariépy, C.; Juárez, M. Review: Pork Belly Quality, Bacon Properties and Recent Consumer Trends. Can. J. Anim. Sci. 2015, 95, 325–340. [Google Scholar] [CrossRef]
- Hermesch, S.; O’Shea, J.M. Genetic Parameters for Characteristics of Pork Bellies. In Application of New Genetic Technologies to Animal Breeding; CSIRO Publishing: Clayton, Australia, 2005; pp. 137–140. [Google Scholar]
- Oh, S.H.; See, M.T. Pork Preference for Consumers in China, Japan and South Korea. Asian-Australas. J. Anim. Sci. 2012, 25, 143–150. [Google Scholar] [CrossRef]
- Soladoye, O.P.; Uttaro, B.; Zawadski, S.; Dugan, M.E.R.; Gariépy, C.; Aalhus, J.L.; Shand, P.; Juárez, M. Compositional and Dimensional Factors Influencing Pork Belly Firmness. Meat Sci. 2017, 129, 54–61. [Google Scholar] [CrossRef]
- Kang, H.S.; Lopez, B.M.; Kim, T.H.; Kim, H.S.; Kim, S.H.; Nam, K.C.; Seo, K.S. Estimation of Genetic Parameters for Pork Belly Components in Yorkshire Pigs. Asian-Australas. J. Anim. Sci. 2015, 28, 922–925. [Google Scholar] [CrossRef]
- Hermesch, S. Genetic Relationships between Composition of Pork Bellies and Performance, Carcase and Meat Quality Traits. Animal 2008, 2, 1178–1185. [Google Scholar] [CrossRef] [PubMed]
- Do, C.-H.; Park, C.-H.; Wasana, N.; Choi, J.-G.; Park, S.-B.; Kim, S.-D.; Cho, G.-H.; Lee, D.-H. Genetic and Phenotypic Relationships of Live Body Measurement Traits and Carcass Traits in Crossbred Pigs of Korea. Korean J. Agric. Sci. 2014, 41, 229–236. [Google Scholar] [CrossRef]
- Miar, Y.; Plastow, G.; Bruce, H.; Moore, S.; Manafiazar, G.; Kemp, R.; Charagu, P.; Huisman, A.; Van Haandel, B.; Zhang, C.; et al. Genetic and Phenotypic Correlations between Performance Traits with Meat Quality and Carcass Characteristics in Commercial Crossbred Pigs. PLoS ONE 2014, 9, e110105. [Google Scholar] [CrossRef]
- Willson, H.E.; de Oliveira, H.R.; Schinckel, A.P.; Grossi, D.; Brito, L.F. Estimation of Genetic Parameters for Pork Quality, Novel Carcass, Primal-Cut and Growth Traits in Duroc Pigs. Animals 2020, 10, 779. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Lopes, M.S.; Lopes, P.S.; Gasparino, E. A Genome-Wide Association Study for Feed Efficiency-Related Traits in a Crossbred Pig Population. Animal 2019, 13, 2447–2456. [Google Scholar] [CrossRef]
- Kim, K.S.; Larsen, N.; Short, T.; Plastow, G.; Rothschild, M.F. A Missense Variant of the Porcine Melanocortin-4 Receptor (MC4R) Gene Is Associated with Fatness, Growth, and Feed Intake Traits. Mamm. Genome 2000, 11, 131–135. [Google Scholar] [CrossRef]
- Soladoye, O.P.; Campos, L.; Aalhus, J.L.; Gariépy, C.; Shand, P.; Juárez, M. Accuracy of Dual Energy X-Ray Absorptiometry (DXA) in Assessing Carcass Composition from Different Pig Populations. Meat Sci. 2016, 121, 310–316. [Google Scholar] [CrossRef]
- Turner, T.D.; Mapiye, C.; Aalhus, J.L.; Beaulieu, A.D.; Patience, J.F.; Zijlstra, R.T.; Dugan, M.E.R. Flaxseed Fed Pork: N-3 Fatty Acid Enrichment and Contribution to Dietary Recommendations. Meat Sci. 2014, 96, 541–547. [Google Scholar] [CrossRef]
- Collison, M.W. Official Methods and Recommended Practices of the AOCS, 7th ed.; American Oil Chemists’ Society: Urbana, IL, USA, 2017. [Google Scholar]
- Uttaro, B.; Zawadski, S.; Larsen, I.; Juárez, M. An Image Analysis Approach to Identification and Measurement of Marbling in the Intact Pork Loin. Meat Sci. 2021, 179, 108549. [Google Scholar] [CrossRef]
- Wei, X.; Bohrer, B.; Uttaro, B.; Juárez, M. Evaluating the Effect of Temperature and Multiple Bends on an Automated Pork Belly Firmness Conveyor Belt Classification System. Meat Sci. 2023, 203, 109222. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A Tool for Genome-Wide Complex Trait Analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Starmer, J.; Martin, E.R. A Multiple Testing Correction Method for Genetic Association Studies Using Correlated Single Nucleotide Polymorphisms. Genet. Epidemiol. 2008, 32, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Peñagaricano, F.; Weigel, K.A.; Khatib, H. Genome-Wide Association Study Identifies Candidate Markers for Bull Fertility in Holstein Dairy Cattle. Anim. Genet. 2012, 43, 65–71. [Google Scholar] [CrossRef]
- Turner, S.D. Qqman: An R Package for Visualizing GWAS Results Using Q-Q and Manhattan Plots. J. Open Source Softw. 2018, 3, 731. [Google Scholar] [CrossRef]
- Badke, Y.M.; Bates, R.O.; Ernst, C.W.; Schwab, C.; Steibel, J.P. Estimation of Linkage Disequilibrium in Four US Pig Breeds. BMC Genom. 2012, 13, 24. [Google Scholar] [CrossRef]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R Package for the Visualization of Intersecting Sets and Their Properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, L.; Ma, J.; Chen, C.; Yang, B.; Huang, L. Transcriptome Analyses Reveal Genes and Pathways Associated with Fatty Acid Composition Traits in Pigs. Anim. Genet. 2017, 48, 645–652. [Google Scholar] [CrossRef]
- van Son, M.; Enger, E.G.; Grove, H.; Ros-Freixedes, R.; Kent, M.P.; Lien, S.; Grindflek, E. Genome-Wide Association Study Confirm Major QTL for Backfat Fatty Acid Composition on SSC14 in Duroc Pigs. BMC Genom. 2017, 18, 369. [Google Scholar] [CrossRef]
- Valdés-Hernández, J.; Folch, J.M.; Crespo-Piazuelo, D.; Passols, M.; Sebastià, C.; Criado-Mesas, L.; Castelló, A.; Sánchez, A.; Ramayo-Caldas, Y. Identification of Candidate Regulatory Genes for Intramuscular Fatty Acid Composition in Pigs by Transcriptome Analysis. Genet. Sel. Evol. 2024, 56, 12. [Google Scholar] [CrossRef] [PubMed]
- González-Prendes, R.; Quintanilla, R.; Mármol-Sánchez, E.; Pena, R.N.; Ballester, M.; Cardoso, T.F.; Manunza, A.; Casellas, J.; Cánovas, Á.; Díaz, I.; et al. Comparing the MRNA Expression Profile and the Genetic Determinism of Intramuscular Fat Traits in the Porcine Gluteus Medius and Longissimus Dorsi Muscles. BMC Genom. 2019, 20, 170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, J.; Cui, L.; Ma, J.; Chen, C.; Ai, H.; Xie, X.; Li, L.; Xiao, S.; Huang, L.; et al. Genetic Architecture of Fatty Acid Composition in the Longissimus Dorsi Muscle Revealed by Genome-Wide Association Studies on Diverse Pig Populations. Genet. Sel. Evol. 2016, 48, 5. [Google Scholar] [CrossRef] [PubMed]
- Estany, J.; Ros-Freixedes, R.; Tor, M.; Pena, R.N. A Functional Variant in the Stearoyl-CoA Desaturase Gene Promoter Enhances Fatty Acid Desaturation in Pork. PLoS ONE 2014, 9, e86177. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, W.; Zhang, Z.; Fan, Y.; Xie, X.; Ai, H.; Ma, J.; Xiao, S.; Huang, L.; Ren, J. Genome-Wide Association Analyses for Fatty Acid Composition in Porcine Muscle and Abdominal Fat Tissues. PLoS ONE 2013, 8, e65554. [Google Scholar] [CrossRef]
- Xing, L.; Lu, X.; Zhang, W.; Wang, Q.; Zhang, W. Genetic Structure and Genome-Wide Association Analysis of Growth and Reproductive Traits in Fengjing Pigs. Animals 2024, 14, 2449. [Google Scholar] [CrossRef]
- Catillo, G.; Zappaterra, M.; Zambonelli, P.; Buttazzoni, L.; Steri, R.; Minelli, G.; Davoli, R. Genome-Wide Association Study Identifies Quantitative Trait Loci Regions Involved in Muscle Acidic Profile in Large White Heavy Pigs. Animal 2020, 14, 1342–1350. [Google Scholar] [CrossRef]
- Passols, M.; Llobet-Cabau, F.; Sebastià, C.; Castelló, A.; Valdés-Hernández, J.; Criado-Mesas, L.; Sánchez, A.; Folch, J.M. Identification of Genomic Regions, Genetic Variants and Gene Networks Regulating Candidate Genes for Lipid Metabolism in Pig Muscle. Animal 2023, 17, 101033. [Google Scholar] [CrossRef]
- Puig-Oliveras, A.; Revilla, M.; Castelló, A.; Fernández, A.I.; Folch, J.M.; Ballester, M. Expression-Based GWAS Identifies Variants, Gene Interactions and Key Regulators Affecting Intramuscular Fatty Acid Content and Composition in Porcine Meat. Sci. Rep. 2016, 6, 31803. [Google Scholar] [CrossRef] [PubMed]
- Ryabova, A.E.; Pozovnikova, M.V.; Dementieva, N.V.; Shcherbakov, Y.S.; Tulinova, O.V.; Romanova, E.A.; Azovtseva, A.I. Analysis of the Genetic Diversity of Ayrshire Cattle in Russia. Message 2. Genome Analysis Based on Data on the Distribution of ROH Patterns in Ayrshire Cows. Ecol. Genet. 2023, 21, 235–248. [Google Scholar] [CrossRef]
- Marín-Garzón, N.A.; Magalhães, A.F.B.; Mota, L.F.M.; Fonseca, L.F.S.; Chardulo, L.A.L.; Albuquerque, L.G. Genome-Wide Association Study Identified Genomic Regions and Putative Candidate Genes Affecting Meat Color Traits in Nellore Cattle. Meat Sci. 2021, 171, 108288. [Google Scholar] [CrossRef] [PubMed]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. Sci. World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, Z.; Bruce, H.; Kemp, R.A.; Charagu, P.; Miar, Y.; Yang, T.; Plastow, G. Genome-Wide Association Studies (GWAS) Identify a QTL Close to PRKAG3 Affecting Meat PH and Colour in Crossbred Commercial Pigs. BMC Genet. 2015, 16, 33. [Google Scholar] [CrossRef] [PubMed]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M.; Gurgul, A. A Comprehensive Transcriptome Analysis of Skeletal Muscles in Two Polish Pig Breeds Differing in Fat and Meat Quality Traits. Genet. Mol. Biol. 2018, 41, 125–136. [Google Scholar] [CrossRef]
- Lee, K.T.; Chung, W.H.; Lee, S.Y.; Choi, J.W.; Kim, J.; Lim, D.; Lee, S.; Jang, G.W.; Kim, B.; Choy, Y.H.; et al. Whole-Genome Resequencing of Hanwoo (Korean Cattle) and Insight into Regions of Homozygosity. BMC Genom. 2013, 14, 519. [Google Scholar] [CrossRef]
- Stuczyńska, A.; Piórkowska, K.; Tyra, M.; Żukowski, K. The Effect of QTL-Rich Region Polymorphisms Identified by Targeted DNA-Seq on Pig Production Traits. Mol. Biol. Rep. 2018, 45, 361–371. [Google Scholar] [CrossRef]
- Ryan, M.T.; Hamill, R.M.; O’Halloran, A.M.; Davey, G.C.; McBryan, J.; Mullen, A.M.; McGee, C.; Gispert, M.; Southwood, O.I.; Sweeney, T. SNP Variation in the Promoter of the PRKAG3 Gene and Association with Meat Quality Traits in Pig. BMC Genet. 2012, 13, 66. [Google Scholar] [CrossRef]
- Verardo, L.L.; Sevón-Aimonen, M.L.; Serenius, T.; Hietakangas, V.; Uimari, P. Whole-Genome Association Analysis of Pork Meat PH Revealed Three Significant Regions and Several Potential Genes in Finnish Yorkshire Pigs. BMC Genet. 2017, 18, 13. [Google Scholar] [CrossRef]
- Chen, X.; Luo, Y.; Jia, G.; Liu, G.; Zhao, H.; Huang, Z. FTO Promotes Adipogenesis through Inhibition of the Wnt/β-Catenin Signaling Pathway in Porcine Intramuscular Preadipocytes. Anim. Biotechnol. 2017, 28, 268–274. [Google Scholar] [CrossRef]
- Gunawan, A.; Basril, S.Y.; Listyarini, K.; Furqon, A.; Bilyaro, W.; Jakaria; Uddin, M.J.; Sumantri, C. Preliminary Study of Solute Carrier Family 23 Member 3 (SLC23A3) Gene as Candidate Marker for Fatty Acid Traits in Kampung-Broiler Crossbred Chickens. J. Indones. Trop. Anim. Agric. 2018, 43, 201–210. [Google Scholar] [CrossRef]
- Ponsuksili, S.; Trakooljul, N.; Basavaraj, S.; Hadlich, F.; Murani, E.; Wimmers, K. Epigenome-Wide Skeletal Muscle DNA Methylation Profiles at the Background of Distinct Metabolic Types and Ryanodine Receptor Variation in Pigs. BMC Genom. 2019, 20, 492. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Izumi-Yamamoto, K.; Iizuka, Y.; Shirota, M.; Nagase, M.; Fujita, T.; Gotoda, T. A Novel Link between Slc22a18 and Fat Accumulation Revealed by a Mutation in the Spontaneously Hypertensive Rat. Biochem. Biophys. Res. Commun. 2013, 440, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Iizuka, Y.; Izumi-Yamamoto, K.; Shirota, M.; Mori, N.; Tahara, Y.; Fujita, T.; Gotoda, T. Overexpression of Slc22a18 Facilitates Fat Accumulation in Mice. Biochem. Biophys. Res. Commun. 2024, 712–713, 149922. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Tan, X.; Yan, Y.; Zhang, T.; Wang, J.; Chen, X.; Xu, J. A Genome-Wide Association Study Identified Candidate Regions and Genes for Commercial Traits in a Landrace Population. Front. Genet. 2024, 15, 1505197. [Google Scholar] [CrossRef]
- Liu, C.; Chen, Z.; Zhang, Z.; Wang, Z.; Guo, X.; Pan, Y.; Wang, Q. Unveiling the Genetic Mechanism of Meat Color in Pigs through GWAS, Multi-Tissue, and Single-Cell Transcriptome Signatures Exploration. Int. J. Mol. Sci. 2024, 25, 3682. [Google Scholar] [CrossRef]
- Desire, S.; Johnsson, M.; Ros-Freixedes, R.; Chen, C.Y.; Holl, J.W.; Herring, W.O.; Gorjanc, G.; Mellanby, R.J.; Hickey, J.M.; Jungnickel, M.K. A Genome-Wide Association Study for Loin Depth and Muscle PH in Pigs from Intensely Selected Purebred Lines. Genet. Sel. Evol. 2023, 55, 42. [Google Scholar] [CrossRef]
- Boshove, A.; Derks, M.F.L.; Sevillano, C.A.; Lopes, M.S.; van Son, M.; Knol, E.F.; Dibbits, B.; Harlizius, B. Large Scale Sequence-Based Screen for Recessive Variants Allows for Identification and Monitoring of Rare Deleterious Variants in Pigs. PLoS Genet. 2024, 20, e1011034. [Google Scholar] [CrossRef]
- Martínez-Montes, Á.M.; Fernández, A.; Muñoz, M.; Noguera, J.L.; Folch, J.M.; Fernández, A.I. Using Genome Wide Association Studies to Identify Common QTL Regions in Three Different Genetic Backgrounds Based on Iberian Pig Breed. PLoS ONE 2018, 13, e0190184. [Google Scholar] [CrossRef]
- Gozalo-Marcilla, M.; Buntjer, J.; Johnsson, M.; Batista, L.; Diez, F.; Werner, C.R.; Chen, C.Y.; Gorjanc, G.; Mellanby, R.J.; Hickey, J.M.; et al. Genetic Architecture and Major Genes for Backfat Thickness in Pig Lines of Diverse Genetic Backgrounds. Genet. Sel. Evol. 2021, 53, 76. [Google Scholar] [CrossRef]
- Torres-Romero, I.; Légeret, B.; Huleux, M.; Sorigue, D.; Damm, A.; Cuiné, S.; Veillet, F.; Blot, C.; Brugière, S.; Couté, Y.; et al. The α/β hydrolase domain-containing protein 1 (ABHD1) acts as a lysolipid lipase and is involved in lipid droplet formation. bioRxiv 2023. Preprint. [Google Scholar] [CrossRef]
- Wang, X.; Song, H.; Liang, J.; Jia, Y.; Zhang, Y. Abnormal Expression of HADH, an Enzyme of Fatty Acid Oxidation, Affects Tumor Development and Prognosis (Review). Mol. Med. Rep. 2022, 26, 355. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, Y.; Sukegawa, S.; Yamashita, M.; Katsuda, N.; Tong, B.; Ohta, T.; Kose, H.; Yamada, T. Identification of Genes Showing Differential Expression Profile Associated with Growth Rate in Skeletal Muscle Tissue of Landrace Weanling Pig. J. Genet. 2016, 95, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Sudre, K.; Cassar-Malek, I.; Listrat, A.; Ueda, Y.; Leroux, C.; Jurie, C.; Auffray, C.; Renand, G.; Martin, P.; Hocquette, J.F. Biochemical and Transcriptomic Analyses of Two Bovine Skeletal Muscles in Charolais Bulls Divergently Selected for Muscle Growth. Meat Sci. 2005, 70, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Xu, Y.; Liang, X.; Wang, Y.; Jin, F.; Liu, D.; Ma, Y.; Yuan, H.; Song, X.; Zeng, W. Porcine Skeletal Muscle Differentially Expressed Gene ATP5B: Molecular Characterization, Expression Patterns, and Association Analysis with Meat Quality Traits. Mamm. Genome 2013, 24, 142–150. [Google Scholar] [CrossRef]
- Kim, N.K.; Park, H.R.; Lee, H.C.; Yoon, D.; Son, E.S.; Kim, Y.S.; Kim, S.R.; Kim, O.H.; Lee, C.S. Comparative Studies of Skeletal Muscle Proteome and Transcriptome Profilings between Pig Breeds. Mamm. Genome 2010, 21, 307–319. [Google Scholar] [CrossRef]
- Li, A.; Mo, D.; Zhao, X.; Jiang, W.; Cong, P.; He, Z.; Xiao, S.; Liu, X.; Chen, Y. Comparison of the Longissimus Muscle Proteome between Obese and Lean Pigs at 180 Days. Mamm. Genome 2013, 24, 72–79. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Ma, C.; Wang, W.; Wang, H.; Jiang, Y. Comparative Transcriptomic Analysis of MRNAs, MiRNAs and LncRNAs in the Longissimus Dorsi Muscles between Fat-Type and Lean-Type Pigs. Biomolecules 2022, 12, 1294. [Google Scholar] [CrossRef]
- Silva, E.F.P.; Gaia, R.C.; Mulim, H.A.; Pinto, L.F.B.; Iung, L.H.S.; Brito, L.F.; Pedrosa, V.B. Genome-Wide Association Study of Conformation Traits in Brazilian Holstein Cattle. Animals 2024, 14, 2472. [Google Scholar] [CrossRef]
- Bongiorni, S.; Gruber, C.E.M.; Bueno, S.; Chillemi, G.; Ferrè, F.; Failla, S.; Moioli, B.; Valentini, A. Transcriptomic Investigation of Meat Tenderness in Two Italian Cattle Breeds. Anim. Genet. 2016, 47, 273–287. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, Y.; Chen, C.; Zhang, L.; Wang, J.; Yang, C.; Wu, T.; Yang, S.; Tao, C.; Wang, Y. Bone Morphogenetic Protein 2 Enhances Porcine Beige Adipogenesis via AKT/MTOR and MAPK Signaling Pathways. Int. J. Mol. Sci. 2024, 25, 3915. [Google Scholar] [CrossRef]
- Pan, J.; Jin, H.; He, L.; Zhu, J.; Zhu, Y.; Wang, Y.; Jin, G.; Tang, X. The Regulatory Mechanism of Salt-Induced Lipid Metabolism in Porcine Biceps Femoris Through Proteomic Analysis of Lipid Droplets. Food Bioprocess Technol. 2024, 17, 4163–4176. [Google Scholar] [CrossRef]
- Banerjee, D.; Girirajan, S. Cross-Ancestry Analysis Identifies Genes Associated with Obesity Risk and Protection. medRxiv 2024. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Gong, H.; Cui, L.; Ma, J.; Chen, C.; Ai, H.; Xiao, S.; Huang, L.; Yang, B. Landscape of Loci and Candidate Genes for Muscle Fatty Acid Composition in Pigs Revealed by Multiple Population Association Analysis. Front. Genet. 2019, 10, 1067. [Google Scholar] [CrossRef] [PubMed]
- Hurov, J.B.; Huang, M.; White, L.S.; Lennerz, J.; Cheol, S.C.; Cho, Y.R.; Kim, H.J.; Prior, J.L.; Piwnica-Worms, D.; Cantley, L.C.; et al. Loss of the Par-1b/MARK2 Polarity Kinase Leads to Increased Metabolic Rate, Decreased Adiposity, and Insulin Hypersensitivity in Vivo. Proc. Natl. Acad. Sci. USA 2007, 104, 5680–5685. [Google Scholar] [CrossRef]
- Fan, B.; Onteru, S.K.; Du, Z.Q.; Garrick, D.J.; Stalder, K.J.; Rothschild, M.F. Genome-Wide Association Study Identifies Loci for Body Composition and Structural Soundness Traits in Pigs. PLoS ONE 2011, 6, e14726. [Google Scholar] [CrossRef]
- Wang, Z.; Chai, J.; Wang, Y.; Gu, Y.; Long, K.; Li, M.; Jin, L. LncPLAAT3-AS Regulates PLAAT3-Mediated Adipocyte Differentiation and Lipogenesis in Pigs through MiR-503-5p. Genes 2023, 14, 161. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, D.; Yin, Y.; Ji, M.; Xu, K.; Huang, X.; Peng, Y.; Zhang, J. Comprehensive Transcriptomic View of the Role of the LGALS12 Gene in Porcine Subcutaneous and Intramuscular Adipocytes. BMC Genom. 2019, 20, 509. [Google Scholar] [CrossRef]
- Piórkowska, K.; Sroka, J.; Żukowski, K.; Zygmunt, K.; Ropka-Molik, K.; Tyra, M. The Effect of BSCL2 Gene on Fat Deposition Traits in Pigs. Animals 2023, 13, 641. [Google Scholar] [CrossRef]
- Kociucka, B.; Jackowiak, H.; Kamyczek, M.; Szydlowski, M.; Szczerbal, I. The Relationship between Adipocyte Size and the Transcript Levels of SNAP23, BSCL2 and COPA Genes in Pigs. Meat Sci. 2016, 121, 12–18. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Z.; Chen, L.; Zhang, H. ITRAQ-Based Proteomic Analysis Reveals Alterations in the Liver Induced by Restricted Meal Frequency in a Pig Model. Nutrition 2016, 32, 871–876. [Google Scholar] [CrossRef]
- Leal-Gutiérrez, J.D.; Elzo, M.A.; Johnson, D.D.; Hamblen, H.; Mateescu, R.G. Genome Wide Association and Gene Enrichment Analysis Reveal Membrane Anchoring and Structural Proteins Associated with Meat Quality in Beef. BMC Genom. 2019, 20, 151. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Zhang, Q.; Wang, H.; Zhang, X.; Tian, M.; Liu, D.; Yang, X. Effects of HOXC8 on the Proliferation and Differentiation of Porcine Preadipocytes. Animals 2023, 13, 2615. [Google Scholar] [CrossRef] [PubMed]
- Parrillo, L.; Spinelli, R.; Costanzo, M.; Florese, P.; Cabaro, S.; Desiderio, A.; Prevenzano, I.; Raciti, G.A.; Smith, U.; Miele, C.; et al. Epigenetic Dysregulation of the Homeobox A5 (HOXA5) Gene Associates with Subcutaneous Adipocyte Hypertrophy in Human Obesity. Cells 2022, 11, 728. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Trait | n | min | max | mean | SD |
|---|---|---|---|---|---|
| IV | 1117 | 50.9 | 75.7 | 60.3 | 3.34 |
| Belly firmness (°) | 494 | 69.8 | 177 | 132 | 26.4 |
| Belly side fat (mm) | 1083 | 0.95 | 4.00 | 2.50 | 0.45 |
| Belly SThK (mm) | 964 | 2.00 | 5.49 | 3.71 | 0.58 |
| Belly Subq (mm) | 1083 | 0.82 | 2.50 | 1.36 | 0.23 |
| Belly Seam (mm) | 964 | 0.40 | 4.07 | 2.12 | 0.58 |
| Trait | Chr | Start, BP | End, BP | SNP (Lowest p-Value) | MAF | p-Value | Var 1 | Candidate Genes 2 |
|---|---|---|---|---|---|---|---|---|
| Iodine value | 14 | 110,834,310 | 112,238,899 | 14:111437307C:T | 0.329 | 1.33 × 10−8 | 0.06 | ENTPD7, COX15, CUTC, ABCC2, Y_RNA, DNMBP, CPN1, CHUK, CWF19L1, SNORA12, BLOC1S2, PKD2L1, SCD, WNT8B, SEC31B, NDUFB8, HIF1AN, ENSSSCG00000044499, ENSSSCG00000049992, ENSSSCG00000058770, PAX2, SLF2, SEMA4G, ssc-mir-10382, MRPL43, TWNK, LZTS2, ENSSSCG00000053081, SFXN3, KAZALD1, and TLX1 |
| Belly firmness | 15 | 120,747,942 | 121,885,708 | 15:121247942C:T | 0.105 | 1.49 × 10−7 | 0.01 | GPBAR1, AAMP, PNKD, CATIP, SLC11A1, CTDSP1, MIR26B, VIL1, USP37, CNOT9, PLCD4, BCS1L, RNF25, STK36, TTLL4, CYP27A1, PRKAG3, WNT6, WNT10A, CDK5R2, FEV, CRYBA2, ssc-mir-375, CFAP65, IHH, NHEJ1, SLC23A3, CNPPD1, RETREG2, ZFAND2B, ABCB6, ATG9A, ANKZF1, GLB1L, STK16, TUBA4A, DNAJB2, PTPRN, MIR153-1, DNPEP, ssc-mir-4334, DES, GMPPA, ASIC4, CHPF, TMEM198, OBSL1, INHA, STK11IP, SLC4A3, and U6 |
| Belly side fat | 1 | 159,498,384 | 160,730,075 | 1:160230075A:C | 0.182 | 1.87 × 10−7 | 0.14 | RNF152 and CDH20 |
| Belly side fat | 2 | 1,451,972 | 2,455,805 | 2:1951972T:C | 0.034 | 3.19 × 10−8 | 0.14 | INS, TH, ASCL2, TSPAN32, CD81, TSSC4, TRPM5, KCNQ1, CDKN1C, SLC22A18, PHLDA2, NAP1L4, CARS1, OSBPL5, NADSYN1, and DHCR7 |
| Belly side fat | 3 | 111,960,719 | 112,960,719 | 3:112460719T:C | 0.019 | 2.45 × 10−7 | 0.08 | TCF23, PREB, ABHD1, KHK, EMILIN1, OST4, AGBL5, TMEM214, MAPRE3, DPYSL5, CENPA, SLC35F6, KCNK3, CIB4, CIMIP2C, OTOF, DRC1, SELENOI, ADGRF3, HADHB, HADHA, GAREM2, RAB10, and U6 |
| Side SthK | 10 | 61,089,623 | 62,089,643 | 10:61589642T:C | 0.205 | 1.68 × 10−7 | 0.1 | ENSSSCG00000061765, ENSSSCG00000045932, ENSSSCG00000055980, andENSSSCG00000052436 |
| side Subq | 2 | 8,082,914 | 9,247,816 | 2:8587647A:T | 0.017 | 1.47 × 10−7 | 0.08 | MACROD1, OTUB1, COX8A, NAA40, RCOR2, MARK2, SPINDOC, ZFTA, ATL3, PLAAT3, LGALS12, PLAAT5, SLC22A8, SLC22A6, SLC3A2, U2, SNORD26, SNORD27, SNORD28, SNORD22, SNORD29, SNORD30, SNORD31, WDR74, TEX54, STX5, NXF1, TMEM223, TMEM179B, TAF6L, POLR2G, TTC9C, HNRNPUL2, BSCL2, UBXN1, UQCC3, CSKMT, SNORA57, C11orf98, INTS5, GANAB, B3GAT3, ROM1, EML3, MTA2, and TUT1 |
| side Subq | 13 | 4,535,728 | 5,535,728 | 13:5035728A:G | 0.012 | 2.24 × 10−7 | 0.11 | TBC1D5 and U6, SATB1 |
| side Subq | 14 | 132,507,485 | 133,507,498 | 14:133007485G:C | 0.014 | 4.65 × 10−8 | 0.13 | PSTK, IKZF5, ACADSB, ssc-mir-4331-2, HMX3, HMX2, BUB3, GPR26, CPXM2, and CHST15 |
| side Subq | 18 | 44,588,816 | 45,588,816 | 18:45088816A:G | 0.014 | 1.06 × 10−7 | 0.1 | JAZF1, TAX1BP1, HIBADH, U6, EVX1, HOXA13, HOXA11, HOXA10, ssc-mir-196b-1, HOXA9, HOXA7, HOXA5, HOXA3, HOXA2, and HOXA1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 His Majesty the King in Right of Canada, as represented by the Minister of Agriculture and Agri-Food. This is an open access article distributed under the CC BY license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mozduri, Z.; Plastow, G.; Dekkers, J.; Houlahan, K.; Kemp, R.; Juárez, M. Genome-Wide Association Study for Belly Traits in Canadian Commercial Crossbred Pigs. Animals 2025, 15, 1254. https://doi.org/10.3390/ani15091254
Mozduri Z, Plastow G, Dekkers J, Houlahan K, Kemp R, Juárez M. Genome-Wide Association Study for Belly Traits in Canadian Commercial Crossbred Pigs. Animals. 2025; 15(9):1254. https://doi.org/10.3390/ani15091254
Chicago/Turabian StyleMozduri, Zohre, Graham Plastow, Jack Dekkers, Kerry Houlahan, Robert Kemp, and Manuel Juárez. 2025. "Genome-Wide Association Study for Belly Traits in Canadian Commercial Crossbred Pigs" Animals 15, no. 9: 1254. https://doi.org/10.3390/ani15091254
APA StyleMozduri, Z., Plastow, G., Dekkers, J., Houlahan, K., Kemp, R., & Juárez, M. (2025). Genome-Wide Association Study for Belly Traits in Canadian Commercial Crossbred Pigs. Animals, 15(9), 1254. https://doi.org/10.3390/ani15091254