
First Bisexually Dimorphic Phytoptid Taxon (Eriophyoidea, Phytoptidae) from Gondwanian Angiosperm Host

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Morphological Measurements
2.2. DNA Extraction and Sequencing
2.3. Sequence Alignment and Molecular Phylogenetic Analyses
3. Results
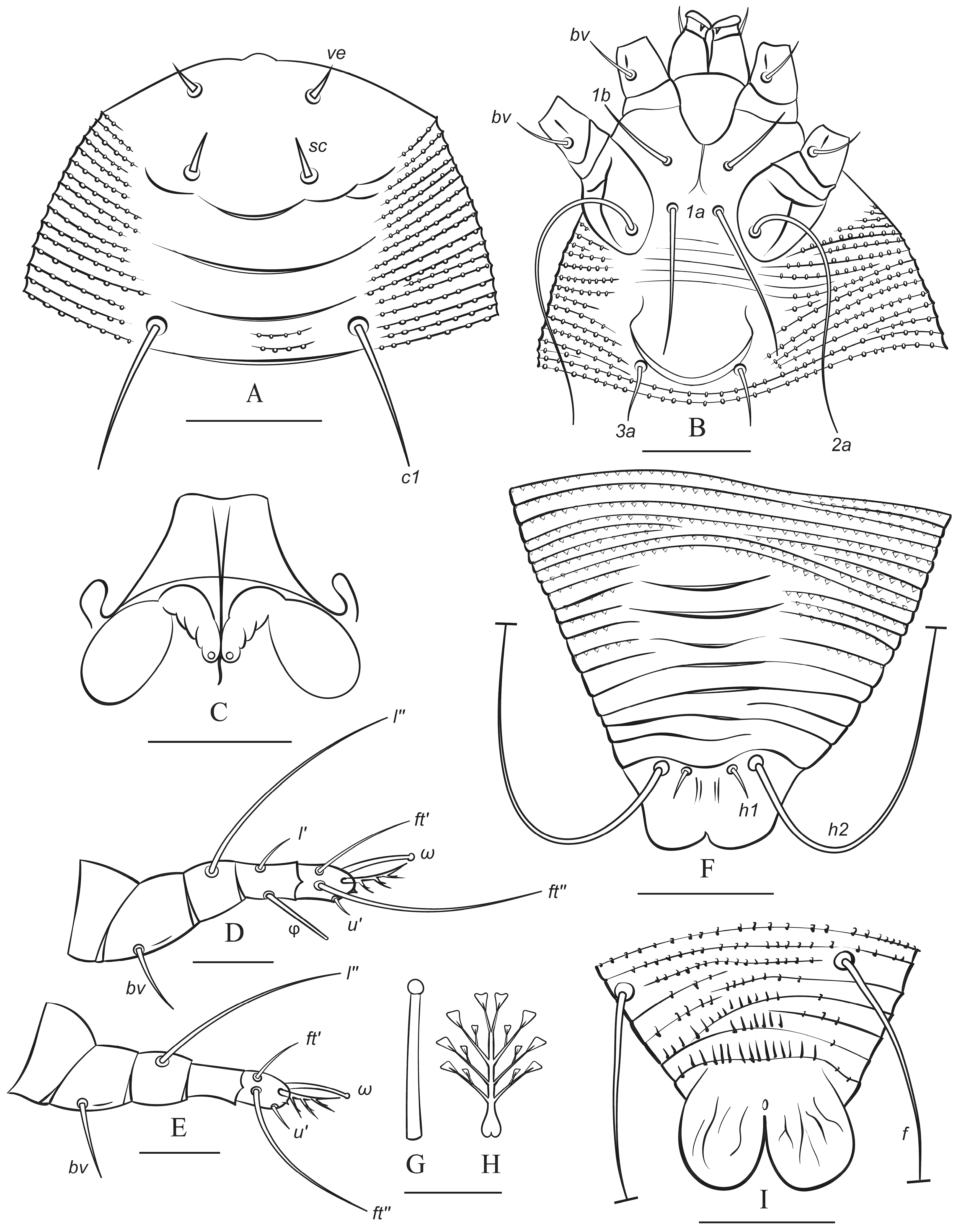
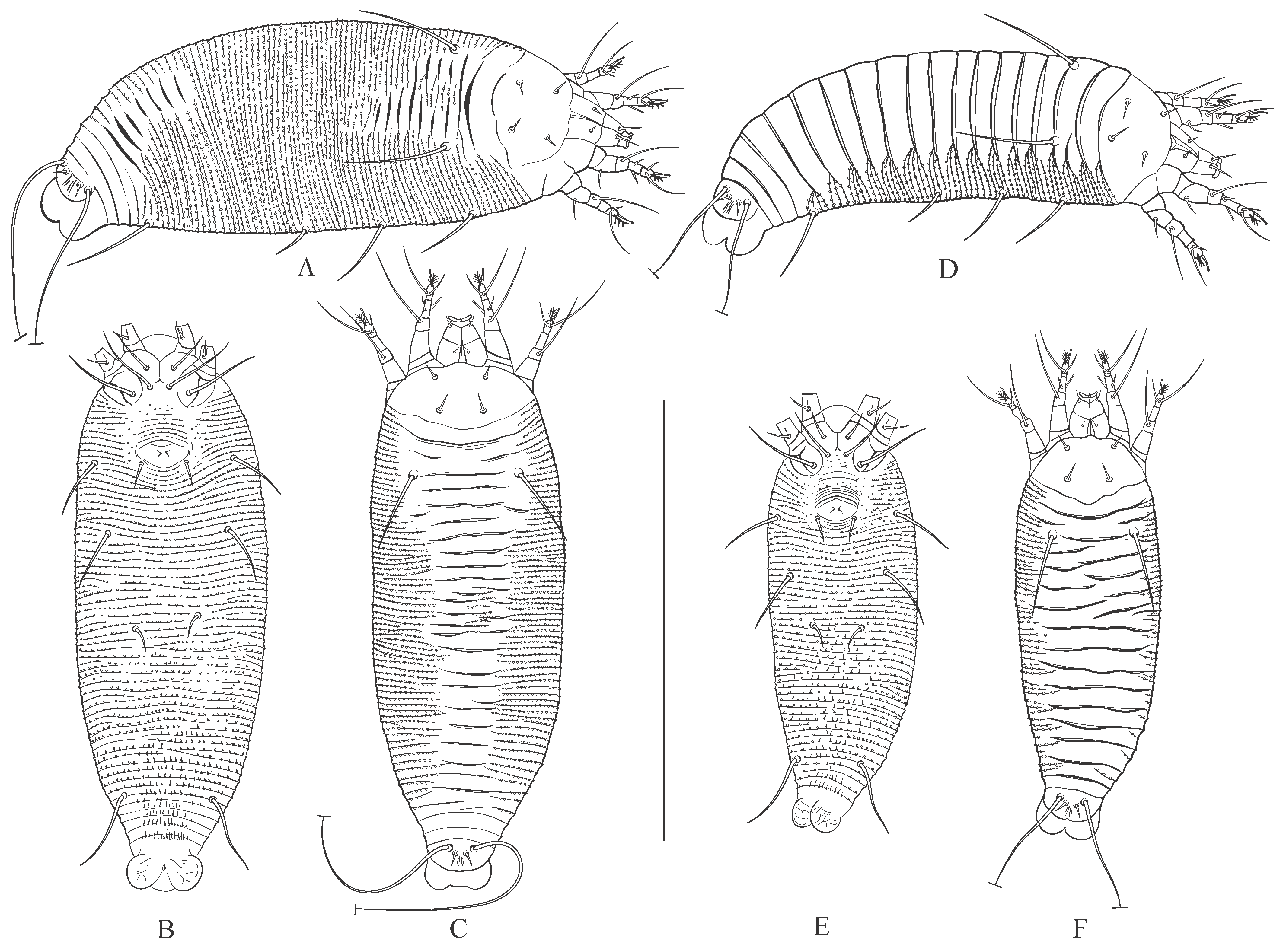
3.1. Microscopic Observations and Morphotypes of Austracus havrylenkonis
3.1.1. Female and Male Morphotype I (MT-I)
3.1.2. Female and Male Morphotype II (MT-II)
3.2. Molecular Phylogenetics
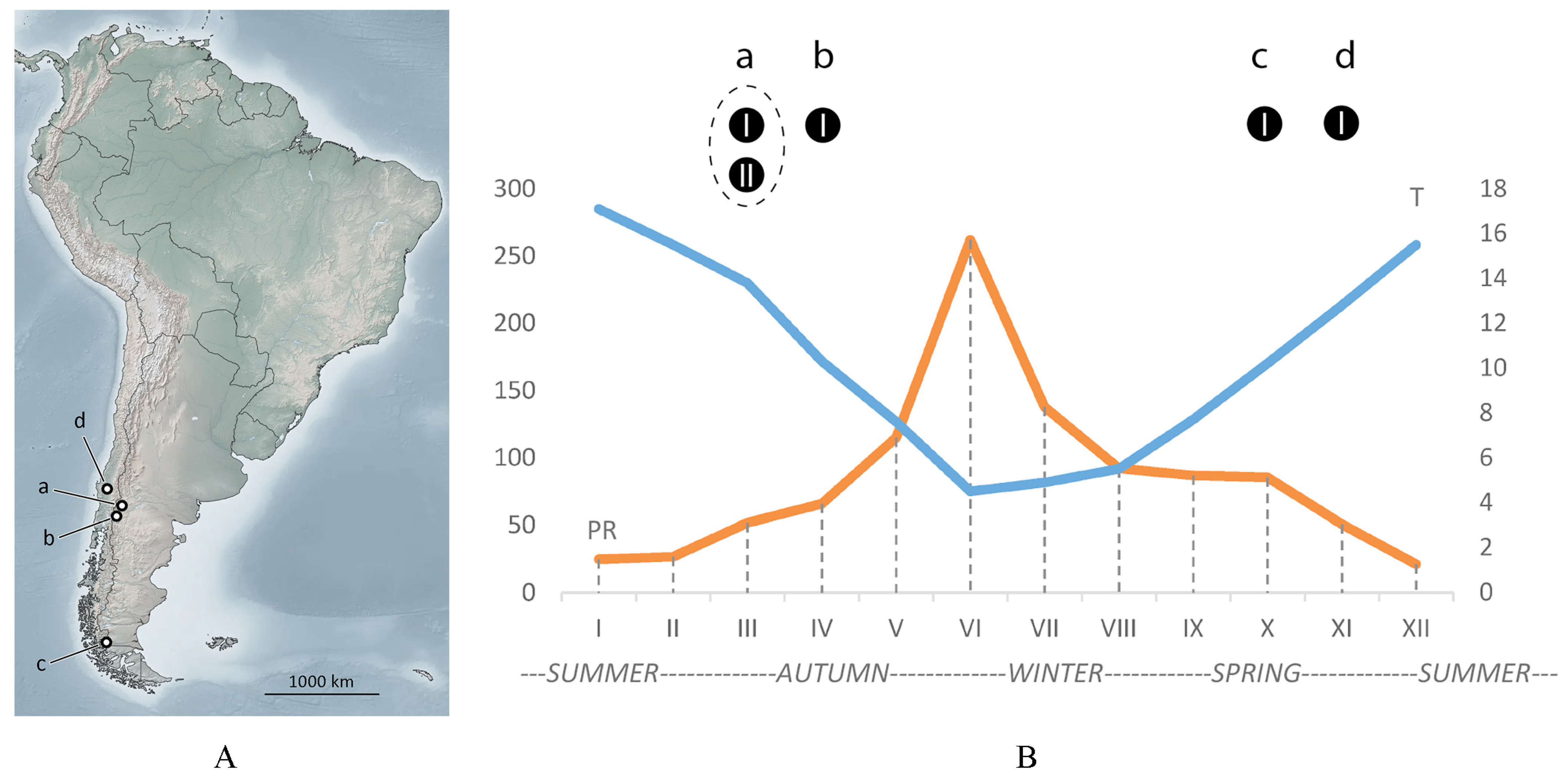
3.3. Seasonal Findings of Austracus and Data on Molting, Sperm Storage, and Putative Dispersal
3.4. Morphological Differences Between Seasonal Morphotypes of Austracus havrylenkonis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walter, D.E.; Lindquist, E.E.; Smith, I.M.; Cook, D.R.; Krantz, G.W. Order Trombidiformes. In A Manual of Acarology; Krantz, G.W., Walter, D.E., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 233–420. [Google Scholar]
- Sukhareva, S.I. Family Phytoptidae Murray 1877 (Acari: Tetrapodili), its consisting, structure and suggested ways of evolution. Acarina 1994, 2, 47–72. [Google Scholar]
- Sidorchuk, E.A.; Schmidt, A.R.; Ragazzi, E.; Roghi, G.; Lindquist, E.E. Plant-feeding mite diversity in Triassic amber (Acari: Tetrapodili). J. Syst. Palaeontol. 2015, 13, 129–151. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Craemer, C.; Gankevich, V.D.; Zhuk, A.S. Integrative Taxonomy of the Gall Mite Nothopoda todeica n. sp. (Eriophyidae) from the Disjunct Afro-Australasian Fern Todea barbara: Morphology, Phylogeny, and Mitogenomics. Insects 2023, 14, 507. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, P.E.; Craemer, C.; Cvrković, T.; Klimov, P.B.; Petanović, R.U.; Romanovich, A.E.; Sukhareva, S.I.; Zukoff, S.; Bolton, S.; Amrine, J. Molecular phylogeny of the phytoparasitic mite family Phytoptidae (Acariformes: Eriophyoidea) identified the female genitalic anatomy as a major macroevolutionary factor and revealed multiple origins of gall induction. Exp. Appl. Acarol. 2021, 83, 31–68. [Google Scholar] [CrossRef] [PubMed]
- Keifer, H.H. Eriophyid studies IX. Bull. Calif. Dept. Agric. 1940, 29, 112–117. [Google Scholar]
- Ozman, S.K. Some biological and morphological differences between gall and vagrant forms of Phytoptus avellanae Nal. (Acari: Phytoptidae). Int. J. Acarol. 2000, 26, 215–219. [Google Scholar] [CrossRef]
- Cvrković, T.; Chetverikov, P.; Vidović, B.; Petanović, R. Cryptic speciation within Phytoptus avellanae s.l. (Eriophyoidea: Phytoptidae) revealed by molecular data and observations on molting Tegonotus-like nymphs. Exp. Appl. Acarol. 2016, 68, 83–96. [Google Scholar] [CrossRef]
- Sukhareva, S.I.; Chetverikov, P.E. Morphological transformations in the course of transition from protogyne to deutogene form of female in eriophyoid mites (Acari: Eriophyoidea). Vestn. St.-Peterbg. Univ. 2013, 3, 3–15. (In Russian) [Google Scholar]
- Keifer, H.H. The Eriophyoidea Nalepa. In Mites Injurious to Economic Plants; Jeppson, L.R., Keifer, H.H., Baker, E.W., Eds.; University of California Press: Berkeley, CA, USA, 1975; pp. 327–587. [Google Scholar]
- Ozman, S.K.; Toros, S. Life cycles of Phytoptus avellanae Nal. and Cecidophyopsis vermiformis Nal. (Acarina: Eriophyoidea). Acta Hortic. 1997, 445, 493–501. [Google Scholar] [CrossRef]
- Ozman, S.K.; Toros, S. Damage caused by Phytoptus avellanae Nal. and Cecidophyopsis vermiformis Nal. (Eriophyoidea: Acarina) in hazelnut. Acta Hortic. 1997, 445, 537–543. [Google Scholar] [CrossRef]
- Keifer, H.H. Eriophyid studies XIV. Bull. Calif. Dept. Agric. 1944, 28, 18–38. [Google Scholar]
- Chetverikov, P.E.; Craemer, C. Sierraphytoptines (Eriophyoidea: Phytoptidae) from relict eudicots: Reassignment of Sierraphytoptus taiwanensus to a new genus Solenoplatilobus and refinement of generic diagnosis of Austracus. Syst. Appl. Acarol. 2016, 21, 745–758. [Google Scholar] [CrossRef]
- Peralta, L. Informe Sanitario del Servicio Agrícola y Ganadero Ministerio de Agricultura Chile; Report #90601-8; Servicio Agricola y Ganadero: Santiago, Chile, 2019; pp. 1–2. [Google Scholar]
- Peralta, L. Informe Sanitario del Servicio Agrícola y Ganadero Ministerio de Agricultura Chile; Report #79962-8; Servicio Agricola y Ganadero: Santiago, Chile, 2018; pp. 1–2. [Google Scholar]
- Peralta, L. Informe Sanitario del Servicio Agrícola y Ganadero Ministerio de Agricultura Chile; Report #16659-8; Servicio Agricola y Ganadero: Santiago, Chile, 2020; pp. 1–2. [Google Scholar]
- Peralta, L. Informe Sanitario del Servicio Agrícola y Ganadero Ministerio de Agricultura Chile; Report #52694-8; Servicio Agricola y Ganadero: Santiago, Chile, 2020; pp. 1–2. [Google Scholar]
- Peralta, L. Informe Sanitario del Servicio Agrícola y Ganadero Ministerio de Agricultura Chile; Report #54517-8; Servicio Agricola y Ganadero: Santiago, Chile, 2016; pp. 1–2. [Google Scholar]
- Peralta, L. Informe Sanitario del Servicio Agrícola y Ganadero Ministerio de Agricultura Chile; Report #63839-8; Servicio Agricola y Ganadero: Santiago, Chile, 2022; pp. 1–2. [Google Scholar]
- Peralta, L. Informe Sanitario del Servicio Agrícola y Ganadero Ministerio de Agricultura Chile; Report #37512-8; Servicio Agricola y Ganadero: Santiago, Chile, 2016; pp. 1–2. [Google Scholar]
- Amrine, J.W., Jr.; Manson, D.C.M. Preparation, mounting and descriptive study of eriophyoid mites. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 383–396. [Google Scholar] [CrossRef]
- Lindquist, E.E. External anatomy and notation of structures. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 3–31. [Google Scholar] [CrossRef]
- Amrine, J.W., Jr.; Stasny, T.A.H.; Flechtmann, C.H.W. Revised Keys to the World Genera of the Eriophyoidea (Acari: Prostigmata); Indira Publishing House: West Bloomfield, MI, USA, 2003; pp. 1–244. [Google Scholar]
- Chetverikov, P.E. Video projector: A digital replacement for camera lucida for drawing mites and other microscopic objects. Syst. Appl. Acarol. 2016, 21, 1278–1280. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Bertone, M. First rhyncaphytoptine mite (Eriophyoidea, Diptilomiopidae) parasitizing american hazelnut (Corylus americana): Molecular identification, confocal microscopy, and phylogenetic position. Exp. Appl. Acarol. 2022, 88, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transformation. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Navia, D.; Flechtmann, C.H.; Amrine, J.W., Jr. Supposed ovoviviparity and viviparity in the coconut mite, Aceria guerreronis Keifer (Prostigmata: Eriophyidae), as a result of female senility. Int. J. Acarol. 2005, 31, 63–65. [Google Scholar] [CrossRef]
- Putman, W.L. The plum nursery mite (Phyllocoptes fockeui Nal. & Trt.). In The 70th Annual Report of the Entomological Society of Ontario; Warwick & Sons Publishing: Toronto, ON, Canada, 1939; pp. 33–40. [Google Scholar]
- Keifer, H.H. Eriophyid Studies XII. Bull. Calif. Dept. Agric. 1942, 31, 117–129. [Google Scholar]
- De-Millo, A.P. O dimorfizme samtsov u chetyrekhnogikh kleshchei (Acarina, Eriophyidae). Vestn. Leningr. Univ. 1967, 3, 26–33. (In Russian) [Google Scholar]
- Manson, D.C.M.; Oldfield, G.N. Life forms, deuterogyny, diapause and seasonal development. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 173–183. [Google Scholar] [CrossRef]
- Keifer, H.H. Eriophyid Studies B-7; Bureau of Entomology of the California Department of Agriculture: Sacramento, CA, USA, 1962; pp. 1–20. [Google Scholar]
- Chetverikov, P.E. Phytoptus atherodes n. sp. (Acari: Eriophyoidea: Phytoptidae) and a supplementary description of Phytoptus hirtae Roivainen 1950 from sedges (Cyperaceae). Zootaxa 2011, 3045, 26–44. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Desnitskiy, A.G.; Klimov, P.B.; Ozman-Sullivan, S.K.; Romanovich, A.E.; Sukhareva, S.I. Deuterogyny and the association of two vagrant eriophyoid mites (Acariformes, Eriophyoidea) with the host-plant generative organs of two broad-leaved trees in North-West Russia. Zool. Stud. 2023, 62, e35. [Google Scholar] [CrossRef] [PubMed]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef]
- Fišer, C.; Robinson, C.T.; Malard, F. Cryptic species as a window into the paradigm shift of the species concept. Mol. Ecol. 2018, 27, 613–635. [Google Scholar] [CrossRef]
- Dabert, M.; Witalinski, W.; Kazmierski, A.; Olszanowski, Z.; Dabert, J. Molecular phylogeny of acariform mites (Acari, Arachnida): Strong conflict between phylogenetic signal and long-branch attraction artifacts. Mol. Phylogenet. Evol. 2010, 56, 222–241. [Google Scholar] [CrossRef]
- Skoracka, A.; Magalhães, S.; Rector, B.G.; Kuczyński, L. Cryptic speciation in the Acari: A function of species lifestyles or our ability to separate species? Exp. Appl. Acarol. 2015, 67, 165–182. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Bruin, J. Evolutionary ecology: Life history patterns, food plant choice and dispersal. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 329–366. [Google Scholar] [CrossRef]
- Skoracka, A.; Kuczyński, L.; de Mendonça, R.S.; Dabert, M.; Szydło, W.; Knihinicki, D.; Truol, G.; Navia, D. Cryptic species within the wheat curl mite Aceria tosichella (Keifer) (Acari: Eriophyoidea), revealed by mitochondrial, nuclear and morphometric data. Invertebr. Syst. 2012, 26, 417–433. [Google Scholar] [CrossRef]
- Skoracka, A.; Kuczyński, L.; Rector, B.; Amrine, J.W., Jr. Wheat curl mite and dry bulb mite: Untangling a taxonomic conundrum through a multidisciplinary approach. Biol. J. Linn. Soc. 2014, 111, 421–436. [Google Scholar] [CrossRef]
- Miller, A.D.; Umina, P.A.; Weeks, A.R.; Hoffmann, A.A. Population genetics of the wheat curl mite (Aceria tosichella Keifer) in Australia: Implications for the management of wheat pathogens. Bull. Entomol. Res. 2012, 102, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Laska, A.; Majer, A.; Szydło, W.; Karpicka-Ignatowska, K.; Hornyák, M.; Labrzycka, A.; Skoracka, A. Cryptic diversity within grass-associated Abacarus species complex (Acariformes: Eriophyidae), with the description of a new species, Abacarus plumiger n. sp. Exp. Appl. Acarol. 2018, 76, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Knapp, M.; Stöckler, K.; Havell, D.; Delsuc, F.; Sebastiani, F.; Lockhart, P.J. Relaxed molecular clock provides evidence for long-distance dispersal of Nothofagus (southern beech). PLoS Biol. 2005, 3, e14. [Google Scholar] [CrossRef]
- Cook, L.G.; Crisp, M.D. Not so ancient: The extant crown group of Nothofagus represents a post-Gondwanan radiation. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2005, 272, 2535–2544. [Google Scholar] [CrossRef]
- Amigo, J.; Rodríguez-Guitián, M.A. Bioclimatic and phytosociological diagnosis of the species of the Nothofagus genus (Nothofagaceae) in South America. Int. J. Geobot. Res. 2011, 1, 1–20. [Google Scholar] [CrossRef]
- Marchelli, P.; Gallo, L.A. The combined role of glaciation and hybridization in shaping the distribution of genetic variation in a Patagonian southern beech. J. Biogeogr. 2004, 31, 451–460. [Google Scholar] [CrossRef]
- Guo, J.F.; Li, H.S.; Wang, B.; Xue, X.F.; Hong, X.Y. DNA barcoding reveals the protogyne and deutogyne of Tegolophus celtis sp. nov. (Acari: Eriophyidae). Exp. Appl. Acarol. 2015, 67, 393–410. [Google Scholar] [CrossRef]
- Mattera, M.G.; Pastorino, M.J.; Lantschner, M.V.; Marchelli, P.; Soliani, C. Genetic diversity and population structure in Nothofagus pumilio, a foundation species of Patagonian forests: Defining priority conservation areas and management. Sci. Rep. 2020, 10, 19231. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mite Species | Host | Collecting Data | Reference | Designation in Figure 1 |
|---|---|---|---|---|
| A. havrylenkonis Keifer 1944 | N. dombeyi (inside fruits) | April 1943, Nahuel Huapi National Park (Argentina) | [13] | (a) |
| A. havrylenkonis Keifer 1944 | N. antarctica (inside fruits) | 23 March 2017, San Martín de los Andes (Argentina) | [5] | (b) |
| A. havrylenkonis Keifer 1944 | Accidental females on a swamp sedge | 15 November 2015, Laguna Parrillar National Reserve, Magallanes (Chile) | [14] | (c) |
| A. havrylenkonis Keifer 1944 | N. antarctica (vagrant females on young leaves) | 15 November 2019, Puerto Natales, Ultima Esperanza, Magallanes (Chile) | [15] | |
| A. havrylenkonis Keifer 1944 | N. dombeyi | 4 October 2018, La Araucania, comuna Curarrehue, Villarica (Chile) | [16] | (d) |
| Austracus sp. | N. dombeyi | 3 February 2020, La Araucania, comuna Curarrehue, Villarica (Chile) | [17] | * |
| Austracus sp. | N. dombeyi | 2 September 2020, La Araucania, Vilcun (Chile) | [18] | * |
| Austracus sp. | N. dombeyi | 8 June 2016, La Araucania, Vilcun (Chile) | [19] | * |
| Austracus sp. | N. dombeyi | 7 September 2020, La Araucania, Angol (Chile) | [20] | * |
| Austracus sp. | N. pumilio | 4 April 2016, Alto Bio Bio (Chile) | [21] | * |
| DNA Isolate | Morphotype (MT) and Number of Mites Used for DNA Extraction | Mite Color | Cox1 | ITS1-5.8S-ITS2 | D1D2 28S | Reference |
|---|---|---|---|---|---|---|
| d488 | MT-I (n = 1) | orange | PQ406503 (510 bp) | PQ421013 (652 bp) | PQ421009 (971 bp) | this study |
| d489 | MT-I (n = 1) | orange | PQ406504 (492 bp) | PQ421014 (703 bp) | – | this study |
| d490 | MT-I (n = 2) | orange | PQ406505 (502 bp) | PQ421015 (703 bp) | PQ421010 (976 bp) | this study |
| d491 | MT-II (n = 1) | white | PQ406506 (502 bp) | PQ421016 (643 bp) | – | this study |
| d492 | MT-II (n = 1) | white | PQ406507 (399 bp) | PQ421017 (660 bp) | PQ421011 (975 bp) | this study |
| d493 | MT-II (n = 2) | white | PQ406508 (445 bp) | – | PQ421012 (970 bp) | this study |
| d496 | MT-I (n = 1) | orange | – | PQ421018 (432 bp) | – | this study |
| d497 | MT-II (n = 1) | white | – | PQ421019 (614 bp) | – | this study |
| d499 | MT-II (n = 1) | white | – | PQ421020 (674 bp) | – | this study |
| F234 | MT-I (n = 1) | white | MT712721 | – | MT712438 | [5] |
| d96 | MT-I (n = 1) | white | – | – | MT712437 | [5] |
| Characters | Summer Females (MT-II), n = 11 | Winter Females (MT-I), n = 12 | Summer Males (MT-II), n = 7 | Winter Males (MT-I), n = 8 |
|---|---|---|---|---|
| Length of body | 340 (295–388) | 308 (283–328) | 309 (282–350) | 226 (210–250) |
| Width of body | 107 (98–119) | 91 (86–96) | 106 (99–114) | 90 (84–100) |
| Length of prodorsal shield | 44 (41–53) | 42 (39–46) | 47 (44–50) | 45 (41–50) |
| Width of prodorsal shield | 75 (70–82) | 60 (58–74) | 84 (80–91) | 72 (67–78) |
| Length of ve | 5 (5–6) | 7 (5–8) | 6 (5–7) | 7 (6–7) |
| Distance between ve | 29 (26–32) | 23 (20–27) | 35 (34–35) | 29 (27–31) |
| Length of sc | 7 (6–9) | 10 (8–11) | 8 (6.5–9) | 9 (8–9) |
| Distance between sc | 27 (25–30) | 23 (20–24) | 31 (30–31) | 27 (25–30) |
| Distance between ve and sc | 20 (18–22) | 20 (18–21) | 21 (20–21) | 19 (17–21) |
| Length of gnathosoma | 26 (24–29) | 27 (24–30) | 28 (27–28) | 26 (23–30) |
| Length of suboral plate | 16 (15–17) | 17 (14–18) | 17 (16–17) | 16 (15–17) |
| Width of suboral plate | 16 (15–17) | 17 (14–20) | 19 (18–20) | 16 (15–18) |
| Length of chelicera | 22 (19–24) | 23 (21–24) | 21 (19–23) | 21 (18–22) |
| Length of d (s. antapic) | 3 (2–4) | 4 (2–5) | 4 (3–5) | 3 (2–4) |
| Length of v | 2 (1–3) | 2 (1–2) | 2 (1–2) | 1 (0.5–2) |
| Length of ep | 2 (2–4) | 3 (2.5–4) | 3 (2–3) | 3 (2–4) |
| Length of leg I | 39 (37–42) | 40 (39–43) | 38 (37–38) | 38 (37–39) |
| Length of tarsus I | 8 (7–9) | 9 (8–10) | 8 (7–8) | 8 (6–10) |
| Length of ω I | 10 (9–13) | 11 (10–12) | 8 (6–9) | 9 (8–10) |
| Length of empodium I | 7 (6–8) | 7 (6–8) | 8 (7–8) | 7 (6.5–8) |
| Number of rays of empodium I | 4 (4–4) | 4 (4–4) | 4 (4–4) | 4 (4–4) |
| Length of ft’ I | 9 (7–14) | 15 (10–17) | 12 (10–13) | 11 (8–14) |
| Length of ft” I | 28 (23–32) | 33 (30–37) | 30 (29–30) | 24 (19–32) |
| Length of u’ I | 4 (2–5) | 4 (3.5–5) | 5 (4–5) | 4 (2.5–5) |
| Length of tibia I | 8 (7–9) | 9 (8–11) | 8 (7–8) | 7 (5–8) |
| Length of ϕ | 11 (10–12) | 11 (10–12) | 11 (10–12) | 11 (9.5–12) |
| Length of l’ I | 4 (3–5) | 6 (5–7) | 5 (4–6) | 4 (3–5) |
| Length of genu I | 7 (7–8) | 9 (8–10) | 7 (6–7) | 7 (6–8) |
| Length of l’’ I | 31 (26–34) | 30 (27–33) | 33 (32–34) | 29 (27–32.5) |
| Length of femur I | 12 (11–13) | 12 (11–13) | 12 (11–12) | 11 (9.5–12) |
| Length of bv II | 9 (7–10) | 9 (7–13) | 9 (8–10) | 10 (7–13) |
| Length of leg II | 37 (35–39) | 39 (36–41) | 37 (36–37) | 35 (32–36) |
| Length of tarsus II | 8 (7–9) | 9 (8–10) | 7 (6–7) | 8 (6–9) |
| Length of ω II | 9 (7–11) | 12 (10–16) | 10 (9–10) | 9 (9–10) |
| Length of empodium II | 6 (5–7) | 6 (5–7.5) | 8 (7–8) | 7 (7–8) |
| Number of rays of empodium II | 4 (4–4) | 4 (4–4) | 4 (4–4) | 4 (4–4) |
| Length of ft’ II | 6 (5–7) | 7 (6–8) | 4 (3–5) | 5 (4–6) |
| Length of ft” II | 25 (21–29) | 30 (26–33) | 26 (25–27) | 27 (23–29) |
| Length of u’ II | 4 (3–5) | 5 (4–6) | 5 (4–5) | 4 (3–4) |
| Length of tibia II | 6 (5–7) | 8 (7–9) | 7 (6–7) | 5 (4.5–6) |
| Length of genu II | 7 (6–8) | 8 (7–9) | 7 (6–7) | 6 (5.5–7) |
| Length of l” II | 26 (20–29) | 25 (21–29) | 29 (27–30) | 27 (24–30) |
| Length of femur II | 10 (7–12) | 13 (11–13.5) | 10 (9–11) | 11 (10–12) |
| Length of bv II | 12 (9–17) | 12 (10–16) | 14 (12–16) | 11 (7–13) |
| Length of prosternal apodeme | 7 (6–8) | 8 (7–11) | 9 (8–10) | 8 (6–9) |
| Length of 1a | 26 (21–38) | 27 (22–37) | 27 (26–27) | 27 (21–38) |
| Length of 1b | 14 (10–19) | 13 (7–16) | 14 (13–15) | 12 (11–13) |
| Length of 2a | 46 (37–62) | 50 (32–63) | 40 (31–48) | 39 (30–48) |
| Distance between 1a | 11 (10–13) | 12 (10–14) | 11 (10–12) | 11 (10–11) |
| Distance between 1b | 17 (14–18) | 15 (14–19) | 16 (15–17) | 15 (14.5–16) |
| Distance between 2a | 32 (29–35) | 29 (26–34) | 34 (32–35) | 27 (24–29) |
| Length of 3a | 10 (8–12) | 11 (10–13) | 10 (9–11) | 12 (10–13) |
| Distance between 3a | 26 (22–29) | 23 (21–31) | 26 (25–27) | 23 (22–24) |
| Number of coxigenital annuli | 5 (3–6) | 5 (5–6) | 7 (7–7) | 5 (5–5) |
| Length of epigynium/male genital area | 20 (19–23) | 21 (19–24) | 10 (9–13) | 11 (9–14) |
| Width of epigyium/male genital area | 28 (27–31) | 29 (27–30) | 27 (25–29) | 25 (24–28) |
| Number of ventral annuli | 72 (65–82) | 63 (60–66) | 60 (59–61) | 50 (45–52) |
| Number of dorsal annuli | 22 (12–33) | 17 (16–18) | 22 (18–29) | 18 (16–19) |
| Length of c1 | 42 (36–54) | 47 (36–59) | 41 (34–48) | 46 (41–50) |
| Length of c2 | 21 (14–26) | 20 (17–24) | 23 (19–27) | 21 (17–25) |
| Length of d | 24 (20–28) | 20 (15–26) | 27 (25–30) | 20 (15–26) |
| Length of e | 11 (8–13) | 13 (10–17) | 13 (12–14) | 13 (11–15) |
| Length of f | 30 (20–37) | 37 (29–42) | 38 (37–40) | 31 (30–34) |
| Length of h1 | 7 (5–9) | 7 (6–10) | 6 (5–7.5) | 6 (5–7) |
| Length of h2 | 73 (54–85) | 101 (75–116) | 78 (72–84) | 68 (60–79) |
| Number of annuli before c2 | 12 (10–14) | 10 (9–10) | 11 (10–12) | 9 (8–10) |
| Number of annuli between c2 and d | 15 (11–18) | 13 (12–16) | 10 (9–10) | 10 (9–10) |
| Number of annuli between d and e | 12 (11–14) | 13 (10–15) | 11 (10–12) | 8 (6–9) |
| Number of annuli between e and f | 28 (23–33) | 23 (20–25) | 24 (23–24) | 19 (18–20) |
| Number of annuli between f and h | 5 (4–6) | 5 (4–6) | 5 (4–6) | 5 (4–6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chetverikov, P.E.; Peralta Alba, L.E. First Bisexually Dimorphic Phytoptid Taxon (Eriophyoidea, Phytoptidae) from Gondwanian Angiosperm Host. Animals 2025, 15, 1236. https://doi.org/10.3390/ani15091236
Chetverikov PE, Peralta Alba LE. First Bisexually Dimorphic Phytoptid Taxon (Eriophyoidea, Phytoptidae) from Gondwanian Angiosperm Host. Animals. 2025; 15(9):1236. https://doi.org/10.3390/ani15091236
Chicago/Turabian StyleChetverikov, Philipp E., and Lourdes E. Peralta Alba. 2025. "First Bisexually Dimorphic Phytoptid Taxon (Eriophyoidea, Phytoptidae) from Gondwanian Angiosperm Host" Animals 15, no. 9: 1236. https://doi.org/10.3390/ani15091236
APA StyleChetverikov, P. E., & Peralta Alba, L. E. (2025). First Bisexually Dimorphic Phytoptid Taxon (Eriophyoidea, Phytoptidae) from Gondwanian Angiosperm Host. Animals, 15(9), 1236. https://doi.org/10.3390/ani15091236