
Genetic Diversity and Conservation Priority of Korean Chicken Breeds Using Single-Nucleotide Polymorphism Markers

 , , , ,
, , , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Genotypes
2.2. Data Filtering
2.3. Population Structure and Genetic Diversity
3. Results
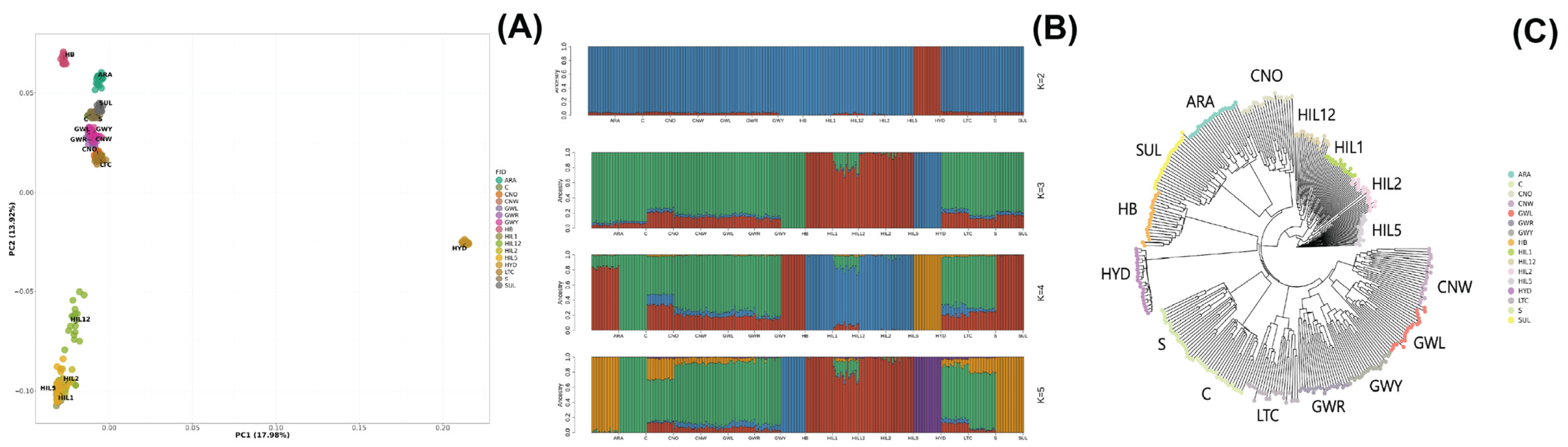
3.1. Population Structure and Genetic Diversity Analysis
3.1.1. Population Structure
3.1.2. Genetic Diversity
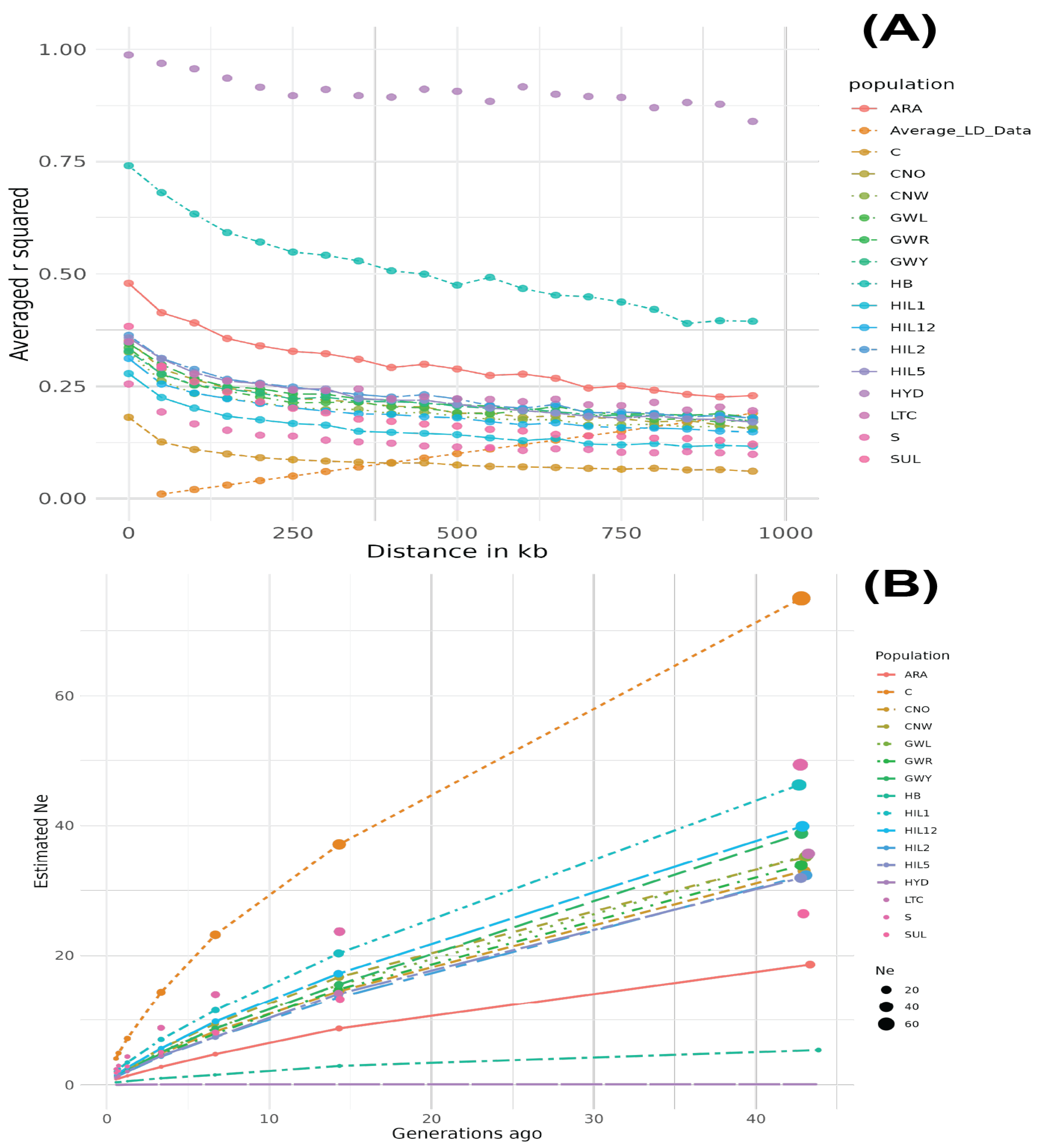
3.1.3. LD and Effective Population Size
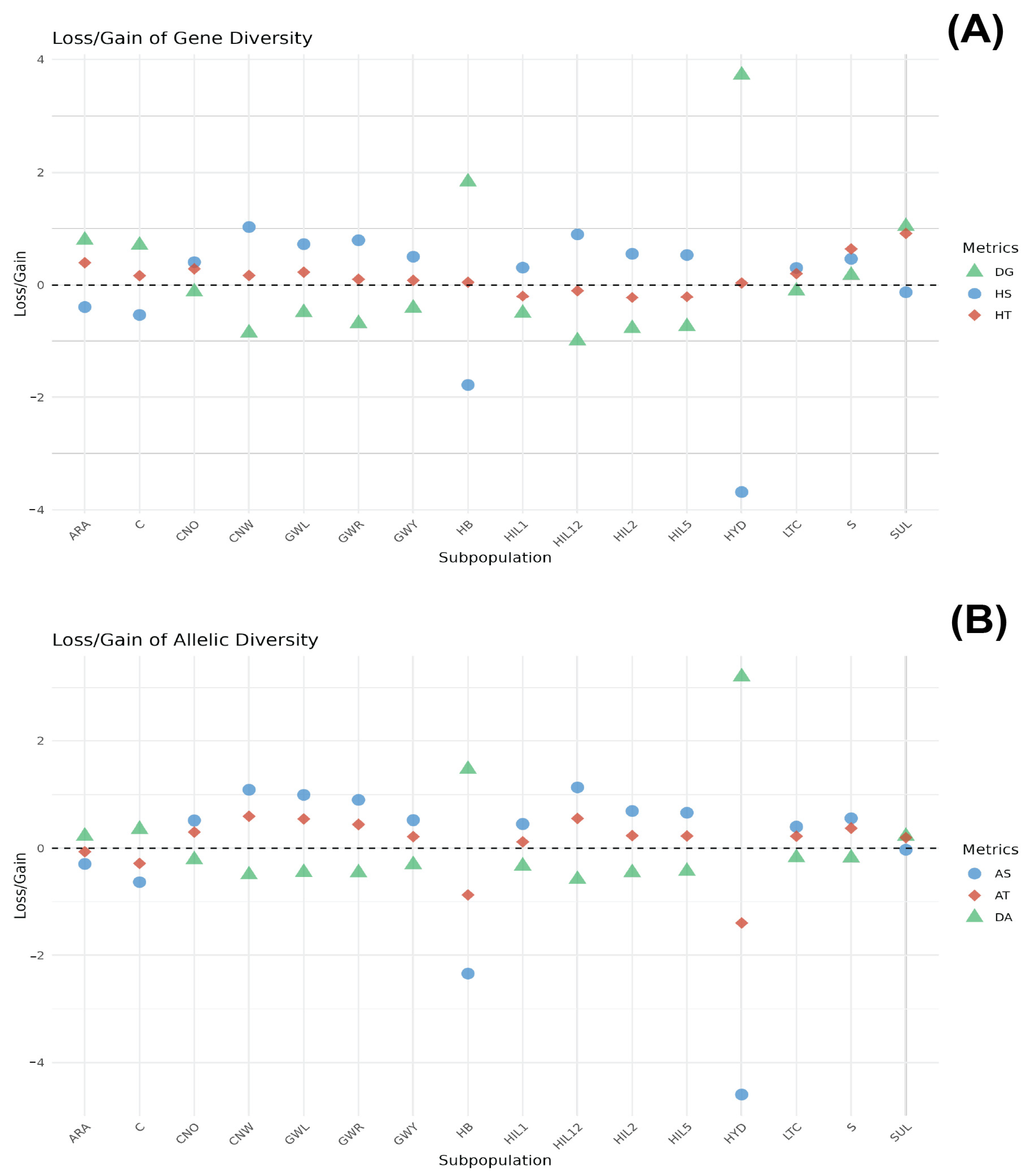
3.2. Contribution of Subpopulation to Total Genetic Diversity
3.2.1. Gene Diversity
3.2.2. Allele Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeWoody, J.A.; Harder, A.M.; Mathur, S.; Willoughby, J.R. The long-standing significance of genetic diversity in conservation. Mol. Ecol. 2021, 30, 4147–4154. [Google Scholar] [CrossRef] [PubMed]
- Catillo, G.; Moioli, B.; Napolitano, F.; Steri, R. Identification of genomic regions harboring diversity between holstein and two local endangered breeds, Modenese and Maremmana. Livest. Sci. 2018, 216, 75–83. [Google Scholar] [CrossRef]
- Makanjuola, B.O.; Miglior, F.; Abdalla, E.A.; Maltecca, C.; Schenkel, F.S.; Baes, C.F. Effect of genomic selection on rate of inbreeding and coancestry and effective population size of holstein and jersey cattle populations. J. Dairy Sci. 2020, 103, 5183–5199. [Google Scholar] [CrossRef] [PubMed]
- Notter, D.R. The importance of genetic diversity in livestock populations of the future. J. Anim. Sci. 1999, 77, 61–69. [Google Scholar] [CrossRef]
- Brown, W.R.A.; Hubbard, S.J.; Tickle, C.; Wilson, S.A. The chicken as a model for large-scale analysis of vertebrate gene function. Nat. Rev. Genet. 2003, 4, 87–98. [Google Scholar] [CrossRef]
- Kralik, G.; Kralik, Z.; Grčević, M.; Hanžek, D. Quality of chicken meat. Anim. Husb. Nutr. 2018, 63. [Google Scholar] [CrossRef]
- Al-Atiyat, R.M.; Al-Rawashdeh, M.; Abu-Alruz, K.; Alasasfa, M.; Salameh, N.; Al-Nawaisah, F.; Al-Khamaiseh, S.; Tabbaa, M.J. Phenotypic characterization and genetic diversity of indigenous chickens of Jordan in comparison with native and commercial breeds for conservation and breeding purposes. Online J. Anim. Feed. Res. 2023, 13, 416–425. [Google Scholar] [CrossRef]
- Muir, W.M.; Wong, G.K.-S.; Zhang, Y.; Wang, J.; Groenen, M.A.M.; Crooijmans, R.P.M.A.; Megens, H.-J.; Zhang, H.; Okimoto, R.; Vereijken, A.; et al. Genome-wide assessment of worldwide chicken SNP genetic diversity indicates significant absence of rare alleles in commercial breeds. Proc. Natl Acad. Sci. USA 2008, 105, 17312–17317. [Google Scholar] [CrossRef]
- Palinkas-Bodzsar, N.; Sztan, N.; Molnar, T.; Hidas, A. Gene conservation of six Hungarian local chicken breeds maintained in small populations over time. PLoS ONE 2020, 15, e0238849. [Google Scholar] [CrossRef]
- Gao, C.; Du, W.; Tian, K.; Wang, K.; Wang, C.; Sun, G.; Kang, X.; Li, W. Analysis of conservation priorities and runs of homozygosity patterns for Chinese indigenous chicken breeds. Animals 2023, 13, 599. [Google Scholar] [CrossRef]
- Hoffmann, I.; Boerma, D.; Scherf, B. The global plan of action for animal genetic resources—The road to common understanding and agreement. Livest. Sci. 2011, 136, 7–14. [Google Scholar] [CrossRef]
- Choi, S.-B.; Byun, M.-J.; Kim, Y.-S.; Kim, M.-J.; Choy, Y.-H.; Kim, D.-H.; Jeong, E.-G.; Kang, K.-S.; Kim, K.-H.; Kim, J.-H. National management system for conservation of livestock genetic resources: An overview. Ann. Anim. Resour. Sci. 2012, 23, 142–148. [Google Scholar] [CrossRef]
- Al-Qamashoui, B.; Simianer, H.; Kadim, I.; Weigend, S. Assessment of genetic diversity and conservation priority of Omani local chickens using microsatellite markers. Trop. Anim. Health Prod. 2014, 46, 747–752. [Google Scholar] [CrossRef]
- Cortes, O.; Cañon, J.; Gama, L.T. Applications of microsatellites and single nucleotide polymorphisms for the genetic characterization of cattle and small ruminants: An overview. Ruminants 2022, 2, 456–470. [Google Scholar] [CrossRef]
- Cendron, F.; Perini, F.; Mastrangelo, S.; Tolone, M.; Criscione, A.; Bordonaro, S.; Iaffaldano, N.; Castellini, C.; Marzoni, M.; Buccioni, A.; et al. Genome-Wide SNP Analysis reveals the population structure and the conservation status of 23 Italian chicken breeds. Animals 2020, 10, 1441. [Google Scholar] [CrossRef] [PubMed]
- Khanyile, K.S.; Dzomba, E.F.; Muchadeyi, F.C. Population genetic structure, linkage disequilibrium and effective population size of conserved and extensively raised village chicken populations of Southern Africa. Front. Genet. 2015, 6, 13. [Google Scholar] [CrossRef]
- Tadano, R.; Nagasaka, N.; Goto, N.; Rikimaru, K.; Tsudzuki, M. Genetic characterization and conservation priorities of chicken lines. Poult. Sci. 2013, 92, 2860–2865. [Google Scholar] [CrossRef]
- Gao, C.; Wang, K.; Hu, X.; Lei, Y.; Xu, C.; Tian, Y.; Sun, G.; Tian, Y.; Kang, X.; Li, W. Conservation priority and run of homozygosity pattern assessment of global chicken genetic resources. Poult. Sci. 2023, 102, 103030. [Google Scholar] [CrossRef]
- FAO. Country Report Questionnaire Country Report Supporting the Preparation of the Third Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; FAO: Roma, Italy, 2024. [Google Scholar]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- López-Cortegano, E.; Pérez-Figueroa, A.; Caballero, A. Metapop2: Re-implementation of software for the analysis and management of subdivided populations using gene and allelic diversity. Mol. Ecol. Resour. 2019, 19, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Roychoudhury, A.K. Sampling variances of heterozygosity and genetic distance. Genetics 1974, 76, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef]
- Zhao, Q.B.; López-Cortegano, E.; Oyelami, F.O.; Zhang, Z.; Ma, P.P.; Wang, Q.S.; Pan, Y.C. Conservation priorities analysis of Chinese indigenous pig breeds in the Taihu Lake region. Front. Genet. 2021, 12, 558873. [Google Scholar] [CrossRef]
- Caballero, A.; Toro, M.A. Analysis of genetic diversity for the management of conserved subdivided populations. Conserv. Genet. 2002, 3, 289–299. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- Molotsi, A.; Dube, B.; Oosting, S.; Marandure, T.; Mapiye, C.; Cloete, S.; Dzama, K. Genetic traits of relevance to sustainability of smallholder sheep farming systems in South Africa. Sustainability 2017, 9, 1225. [Google Scholar] [CrossRef]
- Olschewsky, A.; Hinrichs, D. An overview of the use of genotyping techniques for assessing genetic diversity in local farm animal breeds. Animals 2021, 11, 2016. [Google Scholar] [CrossRef]
- Boettcher, P.J.; Tixier-Boichard, M.; Toro, M.A.; Simianer, H.; Eding, H.; Gandini, G.; Joost, S.; Garcia, D.; Colli, L.; Ajmone-Marsan, P.; et al. Objectives, criteria and methods for using molecular genetic data in priority setting for conservation of animal genetic resources. Anim. Genet. 2010, 41 (Suppl. S1), 64–77. [Google Scholar] [CrossRef]
- Lee, H.-H. Poultry Cultural History; Hyeonchuk Publishing: Gwangmyeong, Republic of Korea, 2019; ISBN 978-89-85841-32-0. [Google Scholar]
- Song, H.M.; Kim, S.C.; Cho, S.R.; Jin, D.H. Investigation of growth and egg production performance of 6 indigenous Korean chicken breeds enrolled in DAD-IS. Korean J. Poult. Sci. 2023, 50, 241–249. [Google Scholar] [CrossRef]
- Jeong, H.; Kim, K.; Caetano-Anollés, K.; Kim, H.; Kim, B.-K.; Yi, J.-K.; Ha, J.-J.; Cho, S.; Oh, D.Y. Whole genome sequencing of Gyeongbuk araucana, a newly developed blue-egg laying chicken breed, reveals its origin and genetic characteristics. Sci. Rep. 2016, 6, 26484. [Google Scholar] [CrossRef] [PubMed]
- Tadano, R.; Kataoka, Y. Genetic diversity in a small chicken population inferred from microsatellite polymorphism. J. Poult. Sci. 2014, 51, 242–247. [Google Scholar] [CrossRef]
- Restoux, G.; Rognon, X.; Vieaud, A.; Guemene, D.; Petitjean, F.; Rouger, R.; Brard-Fudulea, S.; Lubac-Paye, S.; Chiron, G.; Tixier-Boichard, M. Managing genetic diversity in breeding programs of small populations: The case of French local chicken breeds. Genet. Sel. Evol. 2022, 54, 56. [Google Scholar] [CrossRef]
- Kroymann, J.; Mitchell-Olds, T. Epistasis and balanced polymorphism influencing complex trait variation. Nature 2005, 435, 95–98. [Google Scholar] [CrossRef]
- Pettersson, M.; Besnier, F.; Siegel, P.B.; Carlborg, O. Replication and explorations of high-order epistasis using a large advanced intercross line pedigree. PLoS Genet. 2011, 7, e1002180. [Google Scholar] [CrossRef]
- Hua, G.; Chen, J.; Wang, J.; Li, J.; Deng, X. Genetic basis of chicken plumage color in artificial population of complex epistasis. Anim. Genet. 2021, 52, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.B.; Chen, S.R.; Zheng, J.X.; Qu, L.J.; Xu, G.Y.; Yang, N. Developmental phenotypic-genotypic associations of tyrosinase and melanocortin 1 receptor genes with changing profiles in chicken plumage pigmentation. Poult. Sci. 2010, 89, 1110–1114. [Google Scholar] [CrossRef]
- Jansson, E.; Ruokonen, M.; Kojola, I.; Aspi, J. Rise and fall of a wolf population: Genetic diversity and structure during recovery, rapid expansion and drastic decline. Mol. Ecol. 2012, 21, 5178–5193. [Google Scholar] [CrossRef]
- Brito, L.F.; McEwan, J.C.; Miller, S.P.; Pickering, N.K.; Bain, W.E.; Dodds, K.G.; Schenkel, F.S.; Clarke, S.M. Genetic diversity of a New Zealand multi-breed sheep population and composite breeds’ history revealed by a high-density SNP chip. BMC Genet. 2017, 18, 25. [Google Scholar] [CrossRef]
- Adem Esmail, B.; Geneletti, D. Multi-criteria decision analysis for nature conservation: A review of 20 years of applications. Methods Ecol. Evol. 2018, 9, 42–53. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Breed Name | Acronym | N | FAO Class 1 | Origin Source |
|---|---|---|---|---|
| Gyeongbuk Araucana | ARA | 20 | Native | Government |
| Rhode Island | C | 20 | Adapted | Government |
| Chungnam Korean Ogye | CNO | 20 | Native | Government |
| Chungnam white Korean native | CNW | 20 | Native | Government |
| Gangwon black Korean native | GWL | 20 | Native | Government |
| Gangwon reddish Korean native | GWR | 19 | Native | Government |
| Gangwon yellowish Korean native | GWY | 19 | Native | Government |
| Hwangbong-dak | HB | 18 | Native | Private |
| Hyuin black | HIL-1 | 20 | Native | Private |
| Hyuin white | HIL-12 | 19 | Native | Private |
| Hyuin yellow | HIL-2 | 20 | Native | Private |
| Hyuin grey | HIL-5 | 20 | Native | Private |
| Hoeungseong-yakdak | HYD | 20 | Native | Private |
| Long-tail chicken | LTC | 20 | Native | Private |
| Cornish | S | 20 | Adapted | Government |
| White Leghorn | SUL | 20 | Adapted | University |
| Breeds | HO | HE | MAF | FHOM | Fii | Si |
|---|---|---|---|---|---|---|
| ARA | 0.2421 | 0.2262 | 0.1685 | 0.3071 | −0.0611 | −0.0231 |
| C | 0.2231 | 0.2181 | 0.1640 | 0.2670 | −0.0217 | −0.0077 |
| CNO | 0.2834 | 0.2723 | 0.2049 | 0.2072 | −0.0402 | −0.0140 |
| CNW | 0.3293 | 0.3078 | 0.2334 | 0.1270 | −0.0646 | −0.0247 |
| GWL | 0.3056 | 0.2904 | 0.2184 | 0.1936 | −0.0398 | −0.0177 |
| GWR | 0.3155 | 0.2970 | 0.2247 | 0.1677 | −0.0590 | −0.0219 |
| GWY | 0.3013 | 0.2794 | 0.2117 | 0.1925 | −0.0730 | −0.0276 |
| HB | 0.1387 | 0.1353 | 0.1012 | 0.5872 | −0.0245 | −0.0084 |
| HIL-1 | 0.2867 | 0.2668 | 0.2010 | 0.2145 | −0.0700 | −0.0254 |
| HIL-12 | 0.3167 | 0.3033 | 0.2288 | 0.1683 | −0.0396 | −0.0153 |
| HIL-2 | 0.2982 | 0.2808 | 0.2116 | 0.1975 | −0.0584 | −0.0213 |
| HIL-5 | 0.3005 | 0.2796 | 0.2102 | 0.1909 | −0.0706 | −0.0258 |
| HYD | 0.0440 | 0.0388 | 0.0306 | 0.8524 | −0.1264 | −0.0526 |
| LTC | 0.2575 | 0.2665 | 0.2002 | 0.2910 | 0.0230 | 0.0115 |
| S | 0.2842 | 0.2755 | 0.2078 | 0.2125 | −0.0304 | −0.0109 |
| SUL | 0.2551 | 0.2420 | 0.1813 | 0.1223 | −0.0489 | −0.0182 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, H.; Kim, Y.; Lee, S.; Choi, B.; Kim, S.; Jin, D.; Kim, G.; Ha, S.; Mun, S.; Cho, Y.; et al. Genetic Diversity and Conservation Priority of Korean Chicken Breeds Using Single-Nucleotide Polymorphism Markers. Animals 2025, 15, 1084. https://doi.org/10.3390/ani15081084
Song H, Kim Y, Lee S, Choi B, Kim S, Jin D, Kim G, Ha S, Mun S, Cho Y, et al. Genetic Diversity and Conservation Priority of Korean Chicken Breeds Using Single-Nucleotide Polymorphism Markers. Animals. 2025; 15(8):1084. https://doi.org/10.3390/ani15081084
Chicago/Turabian StyleSong, Huimang, Yoonsik Kim, Seunghwan Lee, Bonghwan Choi, Seungchang Kim, Daehyeok Jin, Gaeun Kim, Seungmin Ha, Seongsil Mun, Youngchul Cho, and et al. 2025. "Genetic Diversity and Conservation Priority of Korean Chicken Breeds Using Single-Nucleotide Polymorphism Markers" Animals 15, no. 8: 1084. https://doi.org/10.3390/ani15081084
APA StyleSong, H., Kim, Y., Lee, S., Choi, B., Kim, S., Jin, D., Kim, G., Ha, S., Mun, S., Cho, Y., & Chung, Y. (2025). Genetic Diversity and Conservation Priority of Korean Chicken Breeds Using Single-Nucleotide Polymorphism Markers. Animals, 15(8), 1084. https://doi.org/10.3390/ani15081084