
Differential Mitochondrial Genome Expression of Four Skink Species Under High-Temperature Stress and Selection Pressure Analyses in Scincidae



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Temperature Preferences, Distribution Range, and Grouping Strategies of Sampled Species
2.2. Sample Collection, Acclimation, and High-Temperature Stress
2.3. DNA Extraction and Sequencing
2.4. Mitochondrial Genome Assembly and Annotation
2.5. Phylogenetic Analyses
2.6. Selective Pressure Analyses
2.7. RNA Extraction and cDNA Synthesis
2.8. RT-qPCR Primer Design and Reaction
2.9. RT-qPCR Data Analysis
3. Results
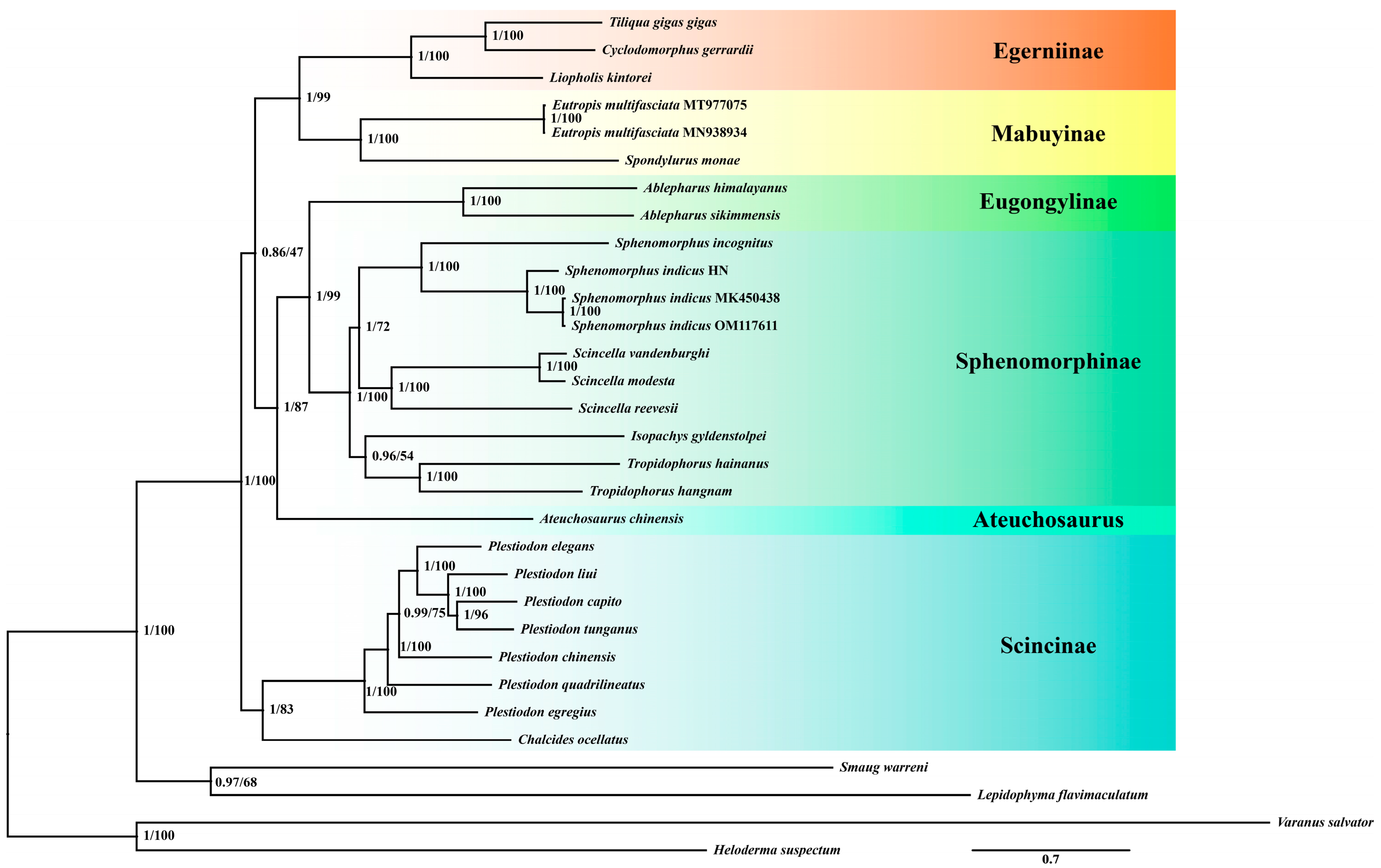
3.1. Phylogenetic Relationships and Selective Pressure Analysis
3.2. Effect of High-Temperature Stress on Transcript Levels of Mitochondrial PCGs
4. Discussion
4.1. Selective Pressure Analysis
4.2. Analysis of Mitochondrial Genome Expression in High-Temperature Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ray, A.; Hughes, L.; Konisky, D.M.; Kaylor, C. Extreme weather exposure and support for climate change adaptation. Glob. Environ. Change 2017, 46, 104–113. [Google Scholar] [CrossRef]
- Johnson, N.C.; Xie, S.P.; Kosaka, Y.; Li, X. Increasing occurrence of cold and warm extremes during the recent global warming slowdown. Nat. Commun. 2018, 9, 1724. [Google Scholar] [CrossRef] [PubMed]
- Sinervo, B.; Méndez-de-la-Cruz, F.; Miles, D.B.; Heulin, B.; Bastiaans, E.; Villagrán-Santa Cruz, M.; Lara-Resendiz, R.; Martínez-Méndez, N.; Calderón-Espinosa, M.L.; Meza-Lázaro, R.N.; et al. Erosion of lizard diversity by climate change and altered thermal niches. Science 2010, 328, 894–899. [Google Scholar] [CrossRef]
- Dubey, S.; Pike, D.A.; Shine, R. Predicting the impacts of climate change on genetic diversity in an endangered lizard species. Clim. Change 2013, 117, 319–327. [Google Scholar] [CrossRef]
- Garcia-Porta, J.; Irisarri, I.; Kirchner, M.; Rodríguez, A.; Kirchhof, S.; Brown, J.L.; MacLeod, A.; Turner, A.P.; Ahmadzadeh, F.; Albaladejo, G.; et al. Environmental temperatures shape thermal physiology as well as diversification and genome-wide substitution rates in lizards. Nat. Commun. 2019, 10, 4077. [Google Scholar] [CrossRef] [PubMed]
- Chapple, D.G.; Roll, U.; Böhm, M.; Aguilar, R.; Amey, A.P.; Austin, C.C.; Baling, M.; Barley, A.J.; Bates, M.F.; Bauer, A.M.; et al. Conservation status of the world’s skinks (scincidae): Taxonomic and geographic patterns in extinction risk. Biol. Conserv. 2021, 257, 109101. [Google Scholar] [CrossRef]
- Meiri, S. Small, rare and trendy: Traits and biogeography of lizards described in the 21st century. J. Zool. 2016, 299, 251–261. [Google Scholar] [CrossRef]
- Blackburn, D.G. Evolution of viviparity in squamate reptiles: Reversibility reconsidered. J. Exp. Zoolog. B Mol. Dev. Evol. 2015, 324, 473–486. [Google Scholar] [CrossRef]
- Greer, A.E. Limb reduction in squamates: Identification of the lineages and discussion of the trends. J. Herpetol. 1991, 25, 166. [Google Scholar] [CrossRef]
- Caldwell, A.J.; While, G.M.; Wapstra, E. Plasticity of thermoregulatory behaviour in response to the thermal environment by widespread and alpine reptile species. Anim. Behav. 2017, 132, 217–227. [Google Scholar] [CrossRef]
- Artacho, P.; Saravia, J.; Perret, S.; Bartheld, J.L.; Le Galliard, J.F. Geographic variation and acclimation effects on thermoregulation behavior in the widespread lizard Liolaemus pictus. J. Therm. Biol. 2017, 63, 78–87. [Google Scholar] [CrossRef]
- Domínguez Guerrero, S.F.; Muñoz, M.M.; Pasten Téllez, D.D.J.; Arenas Moreno, D.M.; Rodríguez Miranda, L.A.; Manríquez Morán, N.L.; Méndez de La Cruz, F.R. Interactions between thermoregulatory behavior and physiological acclimatization in a wild lizard population. J. Therm. Biol. 2019, 79, 135–143. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Huey, R.B.; Deutsch, C.A. Putting the heat on tropical animals. Science 2008, 320, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Thermal tolerance and the global redistribution of animals. Nat. Clim. Change 2012, 2, 686–690. [Google Scholar] [CrossRef]
- Ghalambor, C.K. Are mountain passes higher in the tropics? Janzen’ s hypothesis revisited. Integr. Comp. Biol. 2006, 46, 5–17. [Google Scholar] [CrossRef]
- Calosi, P.; Bilton, D.T.; Spicer, J.I.; Votier, S.C.; Atfield, A. What determines a species’ geographical range? Thermal biology and latitudinal range size relationships in European diving beetles (Coleoptera: Dytiscidae). J. Anim. Ecol. 2010, 79, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Camus, M.F.; Wolff, J.N.; Sgrò, C.M.; Dowling, D.K. Experimental support that natural selection has shaped the latitudinal distribution of mitochondrial haplotypes in Australian Drosophila melanogaster. Mol. Biol. Evol. 2017, 34, 2600–2612. [Google Scholar] [CrossRef]
- Wang, J.Y.; Zhang, L.H.; Hong, Y.H.; Cai, L.N.; Storey, K.B.; Zhang, J.Y.; Zhang, S.S.; Yu, D.N. How does mitochondrial protein-coding gene expression in Fejervarya kawamurai (Anura: Dicroglossidae) respond to extreme temperatures? Animals 2023, 13, 3015. [Google Scholar] [CrossRef]
- Sun, B.J.; Li, T.; Gao, J.; Ma, L.; Du, W.G. High incubation temperatures enhance mitochondrial energy metabolism in reptile embryos. Sci. Rep. 2015, 5, 8861. [Google Scholar] [CrossRef]
- Chung, D.J.; Morrison, P.R.; Bryant, H.J.; Jung, E.; Brauner, C.J.; Schulte, P.M. Intraspecific variation and plasticity in mitochondrial oxygen binding affinity as a response to environmental temperature. Sci. Rep. 2017, 7, 16238. [Google Scholar] [CrossRef]
- Stöck, M.; Moritz, C.; Hickerson, M.; Frynta, D.; Dujsebayeva, T.; Eremchenko, V.; Macey, J.R.; Papenfuss, T.J.; Wake, D.B. Evolution of mitochondrial relationships and biogeography of Palearctic green toads (Bufo viridis subgroup) with insights in their genomic plasticity. Mol. Phylogenet. Evol. 2006, 41, 663–689. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M. Ectotherm mitochondrial economy and responses to global warming. Acta Physiol. 2023, 237, e13950. [Google Scholar] [CrossRef]
- Friedman, J.R.; Nunnari, J. Mitochondrial form and function. Nature 2014, 505, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Ballard, J.W.O.; Pichaud, N. Mitochondrial DNA: More than an evolutionary bystander. Funct. Ecol. 2014, 28, 218–231. [Google Scholar] [CrossRef]
- Sokolova, I. Mitochondrial adaptations to variable environments and their role in animals’ stress tolerance. Integr. Comp. Biol. 2018, 58, 519–531. [Google Scholar] [CrossRef]
- Monlun, M.; Hyernard, C.; Blanco, P.; Lartigue, L.; Faustin, B. Mitochondria as molecular platforms integrating multiple innate immune signalings. J. Mol. Biol. 2017, 429, 1–13. [Google Scholar] [CrossRef]
- Brown, J.A.; Sammy, M.J.; Ballinger, S.W. An evolutionary, or “mitocentric” perspective on cellular function and disease. Redox Biol. 2020, 36, 101568. [Google Scholar] [CrossRef]
- Pond, S.L.K.; Frost, S.D.W. A genetic algorithm approach to detecting lineage-specific variation in selection pressure. Mol. Biol. Evol. 2005, 22, 478–485. [Google Scholar] [CrossRef]
- Yang, Z.H. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, J.; Luo, H.Y.; Chen, X.; Zhong, J.; Ji, X. Climate-driven mitochondrial selection in lacertid lizards. Ecol. Evol. 2024, 14, e11176. [Google Scholar] [CrossRef]
- Jin, Y.T.; Brandt, D.Y.C.; Li, J.S.; Wo, Y.B.; Tong, H.J.; Shchur, V. Elevation as a selective force on mitochondrial respiratory chain complexes of the Phrynocephalus lizards in the Tibetan plateau. Curr. Zool. 2021, 67, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Georgoulis, I.; Feidantsis, K.; Giantsis, I.A.; Kakale, A.; Bock, C.; Pörtner, H.O.; Sokolova, I.M.; Michaelidis, B. Heat hardening enhances mitochondrial potential for respiration and oxidative defence capacity in the mantle of thermally stressed Mytilus galloprovincialis. Sci. Rep. 2021, 11, 17098. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.T.; Guan, J.Y.; Dai, X.Y.; Wu, G.J.; Zhang, L.P.; Storey, K.B.; Zhang, J.Y.; Zheng, R.Q.; Yu, D.N. Mitochondrial gene expression in different organs of Hoplobatrachus rugulosus from China and Thailand under low-temperature stress. BMC Zool. 2022, 7, 24. [Google Scholar] [CrossRef]
- Hong, Y.H.; Yuan, Y.N.; Li, K.; Storey, K.B.; Zhang, J.Y.; Zhang, S.S.; Yu, D.N. Differential mitochondrial genome expression of four hylid frog species under low-temperature stress and its relationship with Amphibian temperature adaptation. Int. J. Mol. Sci. 2024, 25, 5967. [Google Scholar] [CrossRef]
- Zhan, L.M.; He, J.Y.; Ding, L.Y.; Storey, K.B.; Zhang, J.Y.; Yu, D.N. Comparison of mitochondrial genome expression differences among four skink species distributed at different latitudes under low-temperature stress. Int. J. Mol. Sci. 2024, 25, 10637. [Google Scholar] [CrossRef]
- He, J.Y.; Zhan, L.M.; Meng, S.Q.; Wang, Z.; Gao, L.L.; Wang, W.J.; Storey, K.B.; Zhang, Y.P.; Yu, D.N. Differential mitochondrial genome expression of three sympatric lizards in response to low-temperature stress. Animals 2024, 14, 1158. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.M.; He, J.Y.; Meng, S.Q.; Guo, Z.Q.; Chen, Y.X.; Storey, K.B.; Zhang, J.Y.; Yu, D.N. Mitochondrial protein-coding gene expression in the lizard Sphenomorphus incognitus (Squamata: Scincidae) responding to different temperature stresses. Animals 2024, 14, 1671. [Google Scholar] [CrossRef]
- Hedges, S.B. The high-level classification of skinks (Reptilia, Squamata, Scincomorpha). Zootaxa 2014, 3765, 317–338. [Google Scholar] [CrossRef]
- Jiang, Y. A study on habit of Eumeces capito. Sichuan J. Zool. 2005, 24, 370–372. [Google Scholar]
- Zhao, E.; Adler, K. Herpetology of China; Society for the Study of Amphibians & Reptiles: Oxford, OH, USA, 1993. [Google Scholar]
- Hu, J.R.; Du, J.Z.; Ji, X. Pattern of plasma sex steroid hormone levels during the breeding season of male and female skink: Eumeces chinensis. Shi Yan Sheng Wu Xue Bao 2004, 37, 443–448. [Google Scholar]
- Ji, X. Some aspects of thermal biology of the skink (Eumeces chinensis). Chin. Sci. Abstr. Ser. B 1995, 6, 27. [Google Scholar]
- Ji, X.; Sun, P.Y.; Du, W.G. Selected body temperature, thermal tolerance and food assimilation in a Viviparous skink, Sphenomorphus indicus. Neth. J. Zool. 1996, 47, 103–110. [Google Scholar] [CrossRef]
- Cox, C.L.; Logan, M.L.; Nicholson, D.J.; Chung, A.K.; Rosso, A.A.; McMillan, W.O.; Cox, R.M. Species-specific expression of growth-regulatory genes in 2 anoles with divergent patterns of sexual size dimorphism. Integr. Org. Biol. 2022, 4, obac025. [Google Scholar] [CrossRef]
- Robinson, C.D.; Hale, M.D.; Cox, C.L.; John-Alder, H.B.; Cox, R.M. Effects of testosterone on gene expression are concordant between sexes but divergent across species of Sceloporus lizards. Am. Nat. 2024, 204, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2016, 45, gkw955. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; de Pamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Zhan, L.M.; Chen, Y.X.; He, J.Y.; Guo, Z.Q.; Wu, L.; Storey, K.B.; Zhang, J.Y.; Yu, D.N. The phylogenetic relationships of major lizard families using mitochondrial genomes and selection pressure analyses in Anguimorpha. Int. J. Mol. Sci. 2024, 25, 8464. [Google Scholar] [CrossRef]
- Chen, L.; Lin, Y.F.; Xiao, Q.; Lin, Y.; Du, Y.; Lin, C.X.; Ward-Fear, G.; Hu, C.C.; Qu, Y.F.; Li, H. Characterization of the complete mitochondrial genome of the many-lined sun skink (Eutropis multifasciata) and comparison with other Scincomorpha species. Genomics 2021, 113, 2526–2536. [Google Scholar] [CrossRef]
- Kumazawa, Y.; Nishida, M. Complete mitochondrial DNA sequences of the green turtle and blue-tailed mole skink: Statistical evidence for Archosaurian affinity of turtles. Mol. Biol. Evol. 1999, 16, 784–792. [Google Scholar] [CrossRef]
- Song, T.; Zhang, C.L.; Huang, X.; Zhang, B.W. Complete mitochondrial genome of Eumeces elegans (Squamata: Scincidae). Mitochondrial DNA Part A 2016, 27, 719–720. [Google Scholar] [CrossRef]
- Wu, N.; Cai, B.; Chen, M.L.; Guo, X.G. Next-generation sequencing yields a nearly complete mitochondrial genome of Plestiodon liui (Reptilia, Squamata, Scincidae) endemic to China. Mitochondrial DNA Part B 2020, 5, 3637–3638. [Google Scholar] [CrossRef]
- Chen, M.L.; Liu, J.L.; Chen, D.L.; Guo, X.G. The complete mitochondrial genome of a blue-tailed skink (Plestiodon tunganus) endemic to Sichuan Basin. Mitochondrial DNA Part B 2019, 4, 1109–1110. [Google Scholar] [CrossRef]
- Wu, L.; Tong, Y.; Ayivi, S.P.G.; Storey, K.B.; Zhang, J.Y.; Yu, D.N. The complete mitochondrial genomes of three Sphenomorphinae species (Squamata: Scincidae) and the selective pressure analysis on mitochondrial genomes of limbless Isopachys gyldenstolpei. Animals 2022, 12, 2015. [Google Scholar] [CrossRef] [PubMed]
- Kumazawa, Y. Mitochondrial genomes from major lizard families suggest their phylogenetic relationships and ancient radiations. Gene 2007, 388, 19–26. [Google Scholar] [CrossRef]
- Castoe, T.A.; Jiang, Z.J.; Gu, W.; Wang, Z.O.; Pollock, D.D. Adaptive evolution and functional redesign of core metabolic proteins in snakes. PLoS ONE 2008, 3, e2201. [Google Scholar] [CrossRef]
- Kumazawa, Y. Mitochondrial genome of the Komodo Dragon: Efficient sequencing method with reptile-oriented primers and novel gene rearrangements. DNA Res. 2004, 11, 115–125. [Google Scholar] [CrossRef]
- Xiang, C.Y.; Gao, F.L.; Jakovlić, I.; Lei, H.P.; Hu, Y.; Zhang, H.; Zou, H.; Wang, G.T.; Zhang, D. Using phylosuite for molecular phylogeny and tree-based analyses. iMeta 2023, 2, e87. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Xia, X.H. DAMBE7: New and improved tools for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-ng: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar]
- Gao, F.L.; Chen, C.J.; Arab, D.A.; Du, Z.G.; He, Y.H.; Ho, S.Y.W. EasyCodeML: A visual tool for analysis of selection using codeml. Ecol. Evol. 2019, 9, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Weadick, C.J.; Chang, B.S.W. An improved likelihood ratio test for detecting site-specific functional divergence among clades of protein-coding genes. Mol. Biol. Evol. 2012, 29, 1297–1300. [Google Scholar] [CrossRef]
- Zang, X.Y.; Guo, J.L.; Geng, X.F.; Li, P.F.; Sun, J.Y.; Wang, Q.W.; Xu, C.S. Proteome analysis of the liver in the chinese fire-bellied newt cynops orientalis. Genet. Mol. Res. 2016, 15, 10–4238. [Google Scholar] [CrossRef]
- Biederman, J.; Yee, J.; Cortes, P. Validation of internal control genes for gene expression analysis in diabetic glomerulosclerosis. Kidney Int. 2004, 66, 2308–2314. [Google Scholar] [CrossRef]
- Cai, L.N.; Zhang, L.H.; Lin, Y.J.; Wang, J.Y.; Storey, K.B.; Zhang, J.Y.; Yu, D.N. Two-fold ND5 genes, three-fold control regions, lncRNA, and the “missing” ATP8 found in the mitogenomes of Polypedates megacephalus (Rhacophridae: Polypedates). Animals 2023, 13, 2857. [Google Scholar] [CrossRef]
- Moeller, A.H.; Ivey, K.; Cornwall, M.B.; Herr, K.; Rede, J.; Taylor, E.N.; Gunderson, A.R. The lizard gut microbiome changes with temperature and is associated with heat tolerance. Appl. Environ. Microbiol. 2020, 86, e01181-20. [Google Scholar] [CrossRef]
- May, R.A.; Stevenson, K.J. Software review of origin 8. J. Am. Chem. Soc. 2009, 131, 872. [Google Scholar] [CrossRef]
- Austin, J.J.; Arnold, E.N. Using ancient and recent DNA to explore relationships of extinct and endangered Leiolopisma skinks (Reptilia: Scincidae) in the Mascarene islands. Mol. Phylogenet. Evol. 2006, 39, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Whiting, A. Phylogenetic relationships and limb loss in sub-Saharan African Scincine lizards (Squamata: Scincidae). Mol. Phylogenet. Evol. 2003, 29, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Linkem, C.W.; Hesed, K.M.; Diesmos, A.C.; Brown, R.M. Species boundaries and cryptic lineage diversity in a Philippine forest skink complex (Reptilia; Squamata; Scincidae: Lygosominae). Mol. Phylogenet. Evol. 2010, 56, 572–585. [Google Scholar] [CrossRef]
- Brandt, U. Energy converting NADH: Quinone oxidoreductase (complex I). Annu. Rev. Biochem. 2006, 75, 69–92. [Google Scholar] [CrossRef]
- Wirth, C.; Brandt, U.; Hunte, C.; Zickermann, V. Structure and function of mitochondrial complex I. Biochim. Biophys. Acta BBA Bioenerg. 2016, 1857, 902–914. [Google Scholar] [CrossRef]
- Bridges, H.R.; Birrell, J.A.; Hirst, J. The mitochondrial-encoded subunits of respiratory complex I (NADH: Ubiquinone oxidoreductase): Identifying residues important in mechanism and disease. Biochem. Soc. Trans. 2011, 39, 799–806. [Google Scholar] [CrossRef]
- Vartak, R.; Deng, J.; Fang, H.Z.; Bai, Y.D. Redefining the roles of mitochondrial DNA-encoded subunits in respiratory complex I assembly. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2015, 1852, 1531–1539. [Google Scholar] [CrossRef]
- Bai, Y.D.; Attardi, G. The mtDNA-encoded ND6 subunit of mitochondrial NADH dehydrogenase is essential for the assembly of the membrane arm and the respiratory function of the enzyme. EMBO J. 1998, 17, 4848–4858. [Google Scholar] [CrossRef]
- Sazanov, L.A.; Hinchliffe, P. Structure of the hydrophilic domain of respiratory complex I from Thermus thermophilus. Science 2006, 311, 1430–1436. [Google Scholar] [CrossRef]
- Basu, U.; Bostwick, A.M.; Das, K.; Dittenhafer-Reed, K.E.; Patel, S.S. Structure, mechanism, and regulation of mitochondrial dna transcription initiation. J. Biol. Chem. 2020, 295, 18406–18425. [Google Scholar] [CrossRef] [PubMed]
- Hillen, H.S.; Morozov, Y.I.; Sarfallah, A.; Temiakov, D.; Cramer, P. Structural basis of mitochondrial transcription initiation. Cell 2017, 171, 1072–1081.e10. [Google Scholar] [CrossRef]
- Seebacher, F.; Murray, S.A.; Else, P.L. Thermal acclimation and regulation of metabolism in a reptile (Crocodylus porosus): The importance of transcriptional mechanisms and membrane composition. Physiol. Biochem. Zool. 2009, 82, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.; Lima, F.P.; Martel, P.; Castilho, R. Thermal adaptation and clinal mitochondrial DNA variation of European anchovy. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141093. [Google Scholar] [CrossRef]
- Lamb, A.M.; Gan, H.M.; Greening, C.; Joseph, L.; Lee, Y.P.; Morán-Ordóñez, A.; Sunnucks, P.; Pavlova, A. Climate-driven mitochondrial selection: A test in Australian songbirds. Mol. Ecol. 2018, 27, 898–918. [Google Scholar] [CrossRef]
- Ben Slimen, H.; Schaschl, H.; Knauer, F.; Suchentrunk, F. Selection on the mitochondrial ATP synthase 6 and the NADH dehydrogenase 2 genes in hares (Lepus capensis L., 1758) from a steep ecological gradient in North Africa. BMC Evol. Biol. 2017, 17, 46. [Google Scholar] [CrossRef]
- Wu, Z.; Sainz, A.G.; Shadel, G.S. Mitochondrial DNA: Cellular genotoxic stress sentinel. Trends Biochem. Sci. 2021, 46, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, C.M.; Falkenberg, M.; Larsson, N.G. Maintenance and expression of mammalian mitochondrial DNA. Annu. Rev. Biochem. 2016, 85, 133–160. [Google Scholar] [CrossRef]
- Okoye, C.N.; Koren, S.A.; Wojtovich, A.P. Mitochondrial complex I ROS production and redox signaling in hypoxia. Redox Biol. 2023, 67, 102926. [Google Scholar] [CrossRef]
- Pharaoh, G.; Ostrom, E.L.; Stuppard, R.; Campbell, M.; Borghardt, J.M.; Franti, M.; Filareto, A.; Marcinek, D.J. A novel mitochondrial complex I ROS inhibitor partially improves muscle regeneration in adult but not old mice. Redox Biol. 2023, 64, 102770. [Google Scholar] [CrossRef]
- Okoye, C.; Onukwufor, J.; Wojtovich, A. Mechanisms of mitochondrial complex I-ROS mediated behavioral response. Free Radic. Biol. Med. 2022, 192, 89. [Google Scholar] [CrossRef]
- Chai, L.H.; Chen, A.X.; Luo, P.P.; Zhao, H.F.; Wang, H.Y. Histopathological changes and lipid metabolism in the liver of Bufo gargarizans tadpoles exposed to Triclosan. Chemosphere 2017, 182, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhang, Y.H.; Chai, L.H.; Wang, H.Y. Histological changes, lipid metabolism and oxidative stress in the liver of Bufo gargarizans exposed to cadmium concentrations. Chemosphere 2017, 179, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Downs, C.A.; Heckathorn, S.A. The mitochondrial small heat-shock protein protects NADH: Ubiquinone oxidoreductase of the electron transport chain during heat stress in plants. FEBS Lett. 1998, 430, 246–250. [Google Scholar] [CrossRef]
- Michaelsen, J.; Fago, A.; Bundgaard, A. High temperature impairs mitochondrial function in rainbow trout cardiac mitochondria. J. Exp. Biol. 2021, 224, jeb242382. [Google Scholar] [CrossRef]
- Brand, M.D.; Chien, L.F.; Ainscow, E.K.; Rolfe, D.F.S.; Porter, R.K. The causes and functions of mitochondrial proton leak. Biochim. Biophys. Acta BBA Bioenerg. 1994, 1187, 132–139. [Google Scholar] [CrossRef]
- Vaanholt, L.M.; Speakman, J.R.; Garland, T.; Lobley, G.E.; Visser, G.H. Protein synthesis and antioxidant capacity in aging mice: Effects of long-term voluntary exercise. Physiol. Biochem. Zool. 2008, 81, 148–157. [Google Scholar] [CrossRef]
- Okado-Matsumoto, A.; Fridovich, I. Subcellular distribution of superoxide dismutases (SOD) in rat liver. J. Biol. Chem. 2001, 276, 38388–38393. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Guppy, M.; Fuery, C.J.; Flanigan, J.E. Biochemical principles of metabolic depression. Comp. Biochem. Physiol. Part B Comp. Biochem. 1994, 109, 175–189. [Google Scholar] [CrossRef]
- Garvin, M.R.; Bielawski, J.P.; Sazanov, L.A.; Gharrett, A.J. Review and meta-analysis of natural selection in mitochondrial complex I in metazoans. J. Zool. Syst. Evol. Res. 2015, 53, 1–17. [Google Scholar] [CrossRef]
- Morales, H.E.; Pavlova, A.; Joseph, L.; Sunnucks, P. Positive and purifying selection in mitochondrial genomes of a bird with mitonuclear discordance. Mol. Ecol. 2015, 24, 2820–2837. [Google Scholar] [CrossRef] [PubMed]
- Nabholz, B.; Ellegren, H.; Wolf, J.B.W. High levels of gene expression explain the strong evolutionary constraint of mitochondrial protein-coding genes. Mol. Biol. Evol. 2013, 30, 272–284. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genes | Models | Number of Parameters | Log-Likelihood Values | Parameter Estimates | LRT p Values |
|---|---|---|---|---|---|---|
| Plestiodon capito | ND4 | CmC M2a-rel | 58 57 | −17,582.82 −17,587.35 | ω0 = 0.01, p0 = 0.66; ω1 = 1.00, p1 = 0.03; ω2 = 0.53, p2 = 0.31 ω0 = 0.01, p0 = 0.65; ω1 = 1.00, p1 = 0.04; ω2 = 0.15, p2 = 0.31 | 0.00 ** |
| Plestiodon chinensis | ND2 | CmC M2a-rel | 58 57 | −13,841.47 −13,845.21 | ω0 = 0.01, p0 = 0.54; ω1 = 1.00, p1 = 0.01; ω2 = 0.22, p2 = 0.45 ω0 = 0.01, p0 = 0.54; ω1 = 1.00, p1 = 0.01; ω2 = 0.16, p2 = 0.45 | 0.00 ** |
| Plestiodon chinensis | ND5 | CmC M2a-rel | 58 57 | −23,658.81 −23,660.96 | ω0 = 0.01, p0 = 0.62; ω1 = 1.00, p1 = 0.04; ω2 = 0.30, p2 = 0.34 ω0 = 0.01, p0 = 0.62; ω1 = 1.00, p1 = 0.03; ω2 = 0.19, p2 = 0.34 | 0.04 * |
| Sphenomorphus indicus | ND6 | CmC M2a-rel | 58 57 | −6020.09 −6024.34 | ω0 = 0.01, p0 = 0.70; ω1 = 1.00, p1 = 0.03; ω2 = 9.82, p2 = 0.27 ω0 = 0.01, p0 = 0.70; ω1 = 1.00, p1 = 0.03; ω2 = 0.12, p2 = 0.27 | 0.00 ** |
| Scincella modesta | COI | CmC M2a-rel | 58 57 | −15,138.68 −15,141.70 | ω0 = 0.00, p0 = 0.88; ω1 = 1.00, p1 = 0.00; ω2 = 0.47, p2 = 0.12 ω0 = 0.00, p0 = 0.88; ω1 = 1.00, p1 = 0.00; ω2 = 0.11, p2 = 0.12 | 0.01 * |
| Scincella modesta | ND4 | CmC M2a-rel | 58 57 | −17,585.31 −17,587.35 | ω0 = 0.01, p0 = 0.65; ω1 = 1.00, p1 = 0.04; ω2 = 0.65, p2 = 0.31 ω0 = 0.01, p0 = 0.65; ω1 = 1.00, p1 = 0.04; ω2 = 0.15, p2 = 0.31 | 0.04 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Zhan, L.; Storey, K.B.; Zhang, J.; Yu, D. Differential Mitochondrial Genome Expression of Four Skink Species Under High-Temperature Stress and Selection Pressure Analyses in Scincidae. Animals 2025, 15, 999. https://doi.org/10.3390/ani15070999
Wu X, Zhan L, Storey KB, Zhang J, Yu D. Differential Mitochondrial Genome Expression of Four Skink Species Under High-Temperature Stress and Selection Pressure Analyses in Scincidae. Animals. 2025; 15(7):999. https://doi.org/10.3390/ani15070999
Chicago/Turabian StyleWu, Xuxiang, Lemei Zhan, Kenneth B. Storey, Jiayong Zhang, and Danna Yu. 2025. "Differential Mitochondrial Genome Expression of Four Skink Species Under High-Temperature Stress and Selection Pressure Analyses in Scincidae" Animals 15, no. 7: 999. https://doi.org/10.3390/ani15070999
APA StyleWu, X., Zhan, L., Storey, K. B., Zhang, J., & Yu, D. (2025). Differential Mitochondrial Genome Expression of Four Skink Species Under High-Temperature Stress and Selection Pressure Analyses in Scincidae. Animals, 15(7), 999. https://doi.org/10.3390/ani15070999