
Factors Affecting Cryopreservation of Domestic Cat (Felis catus) Epididymal Spermatozoa



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Epididymal Sperm Collection
2.3. Sperm Cryopreservation
2.3.1. Refrigeration
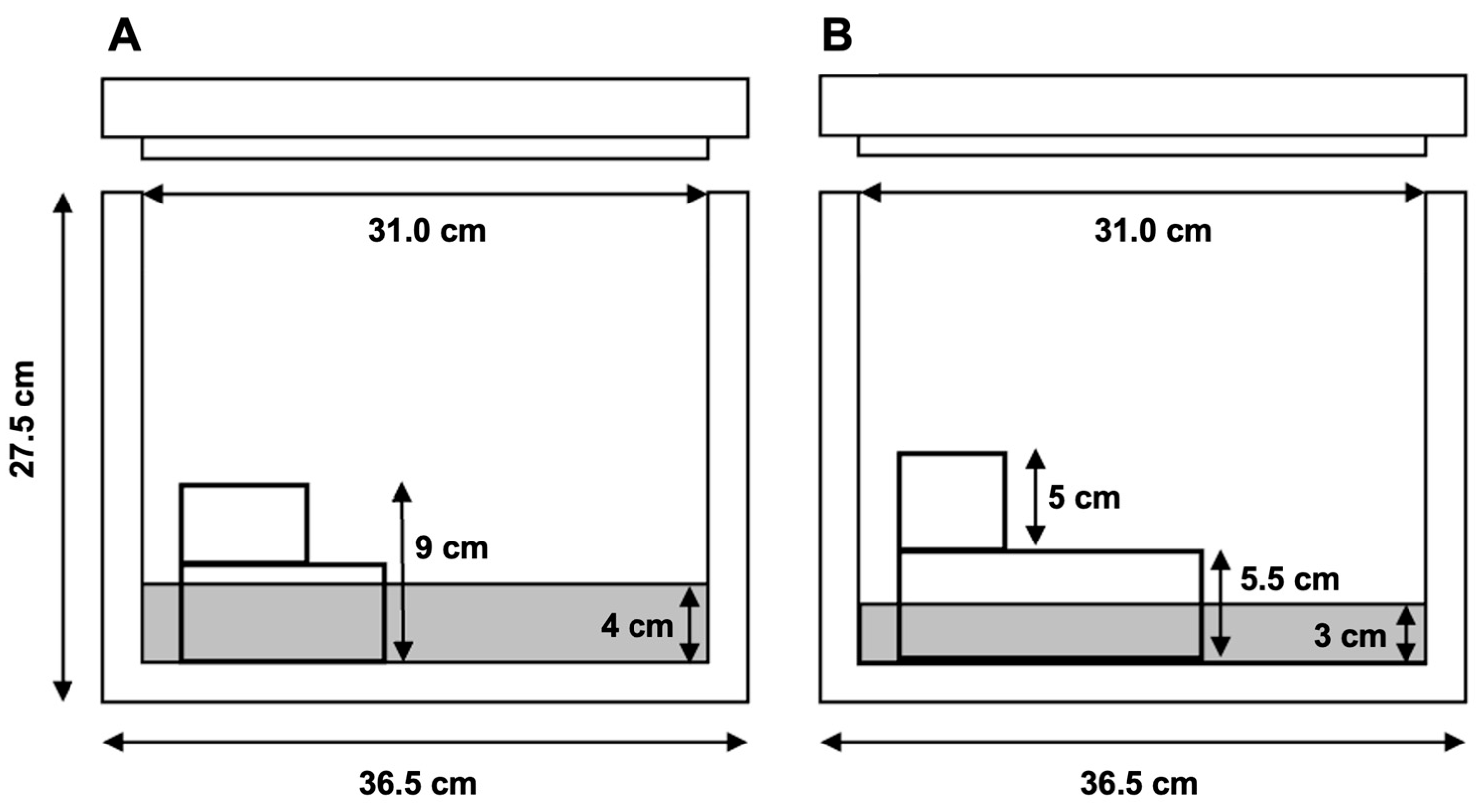
2.3.2. Packaging System
2.3.3. Temperature and Timing of Straw Loading
2.3.4. Freezing
2.4. Sperm Analysis After Thawing
2.5. Sperm Evaluation
2.6. Statistical Analyses
3. Results
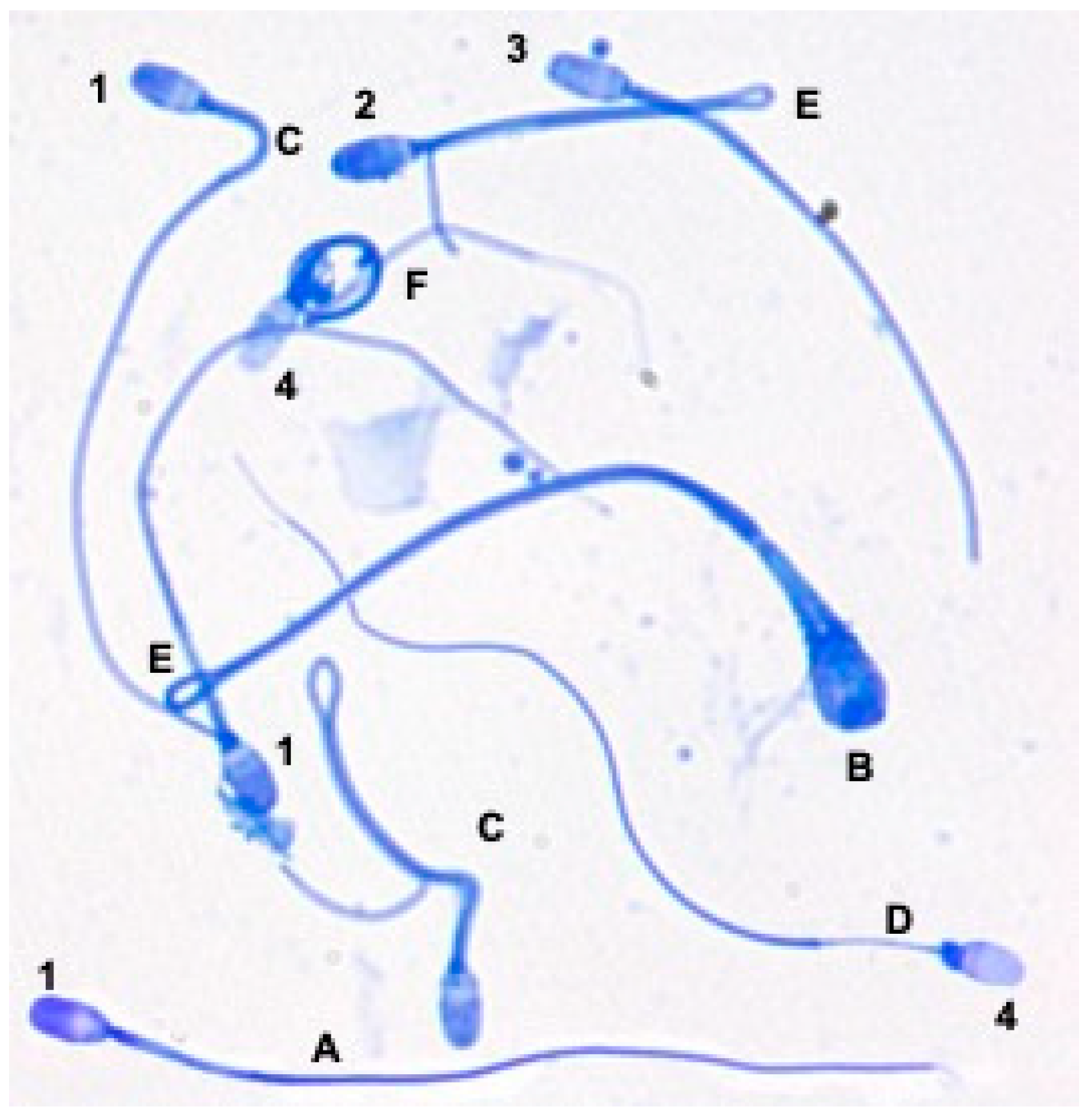
3.1. Sperm Abnormalities in Epididymal Samples of Domestic Cat
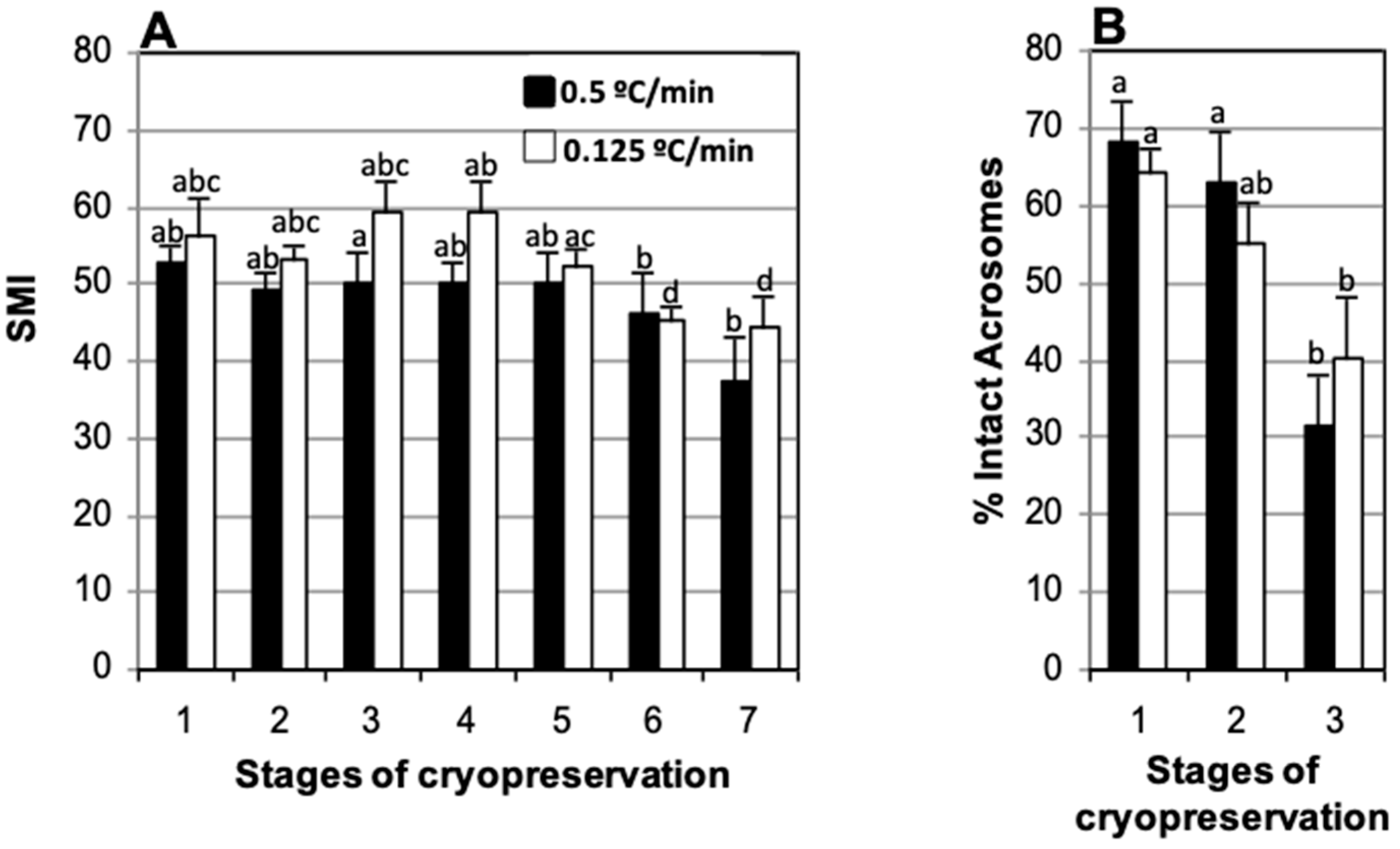
3.2. Effect of Cooling Rate: −0.125 °C/min vs. −0.5 °C/min
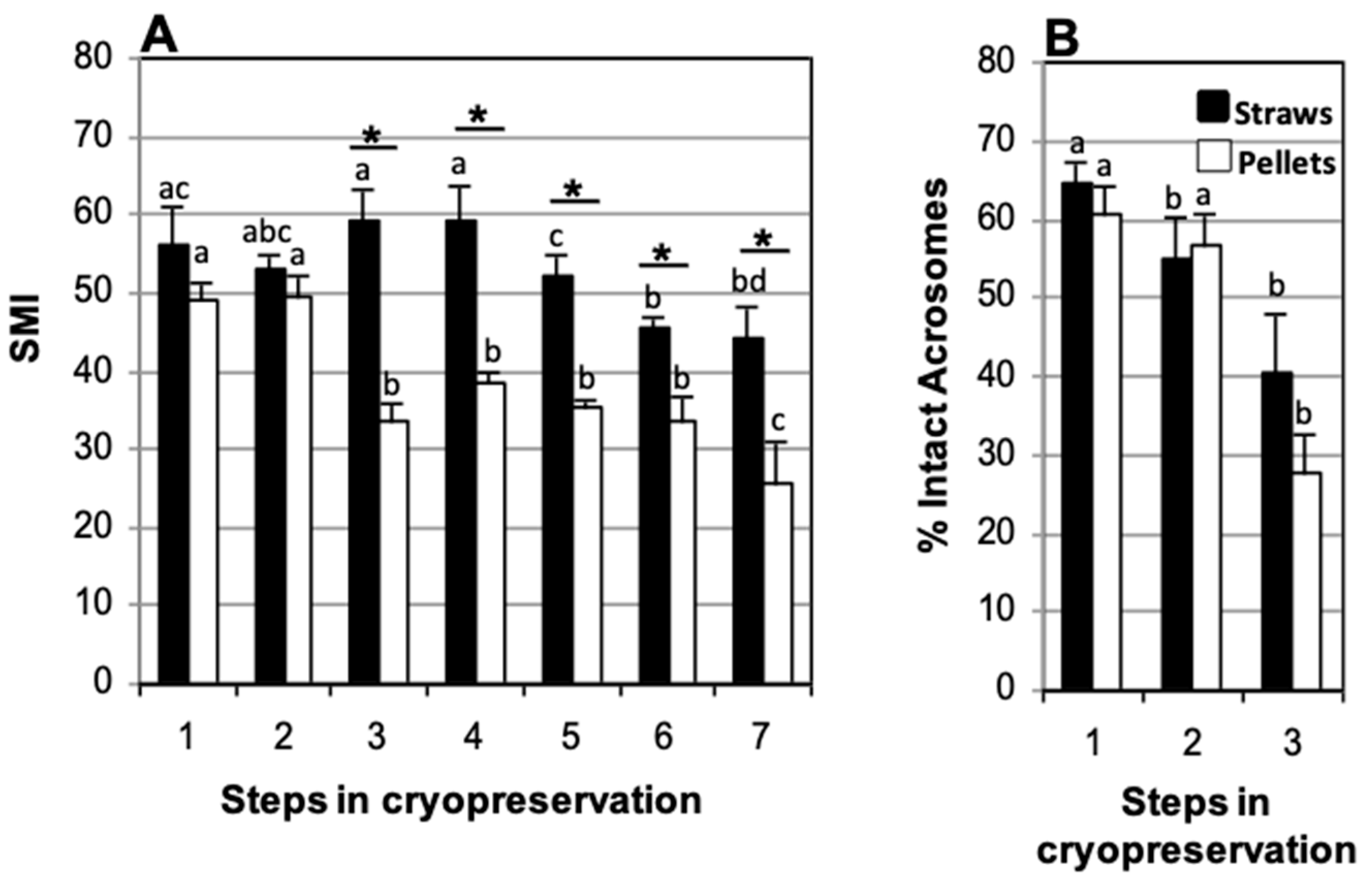
3.3. Effect of Packaging Method: Straws vs. Pellets
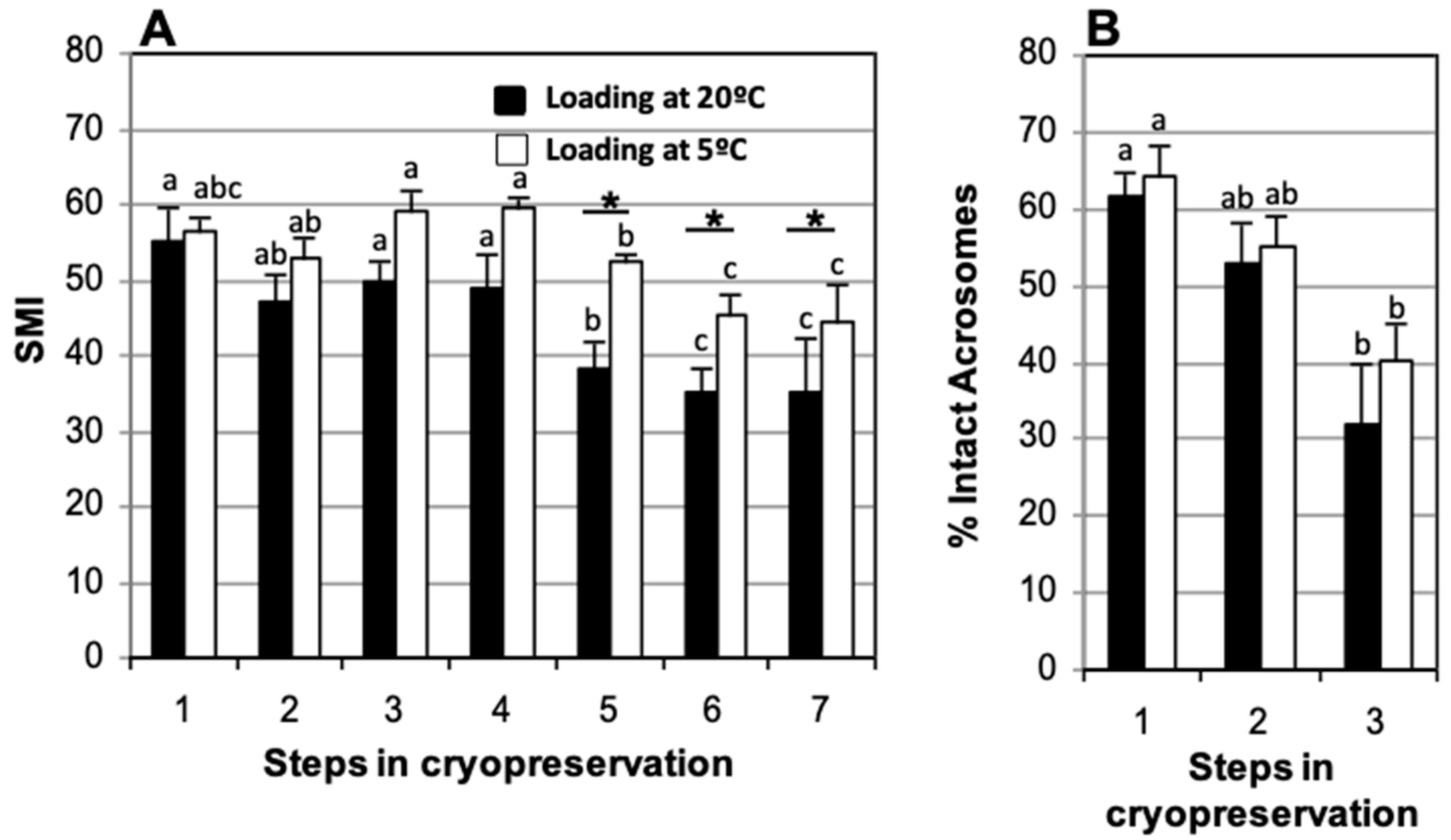
3.4. Straw Loading Temperature: Room Temperature vs. 5 °C
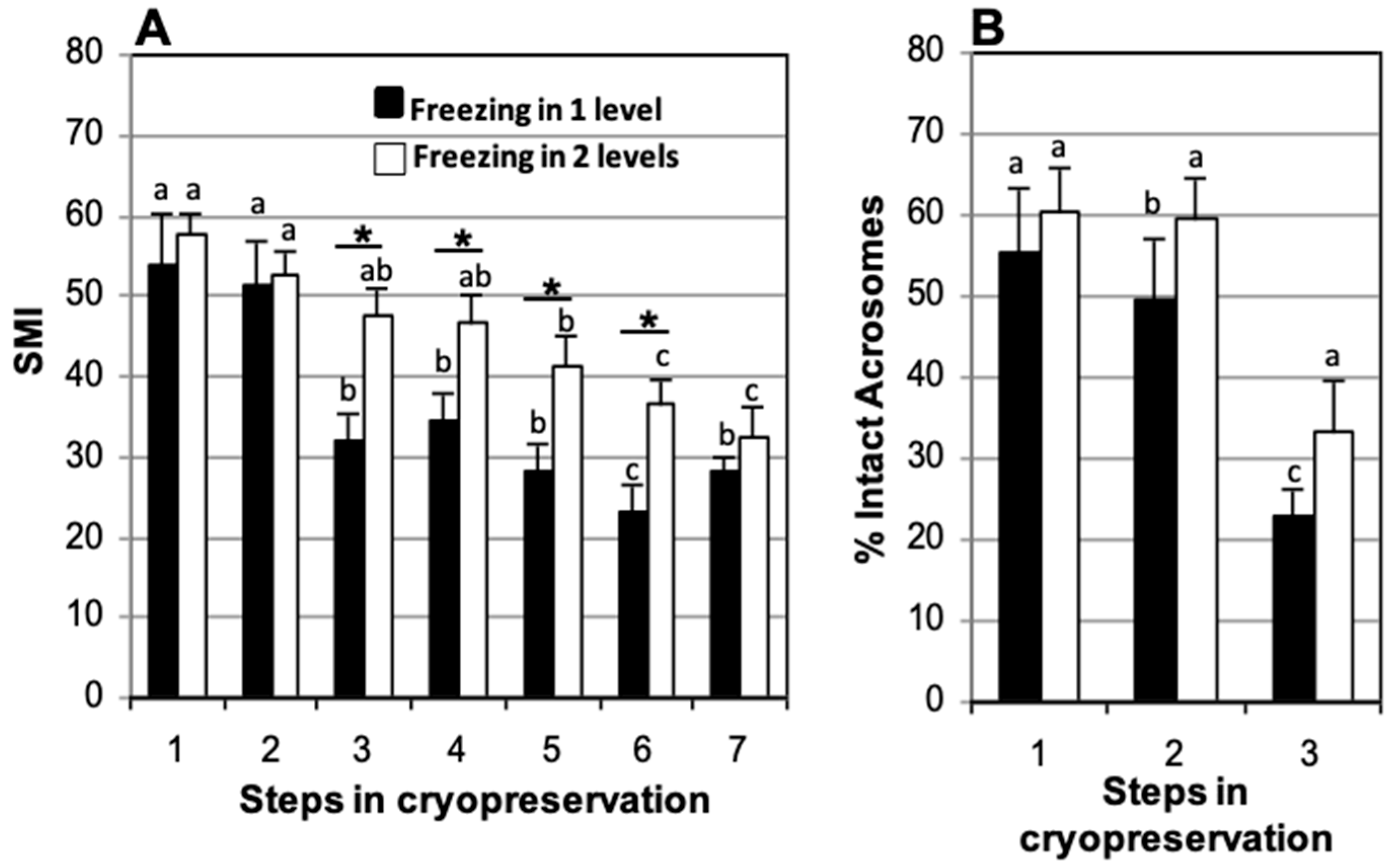
3.5. Effect of Freezing at One or Two Levels over Nitrogen Vapors
3.6. Effect of Interactions Between Factors in the Cryopreservation Process
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wildt, D.E.; Rall, W.F.; Crister, J.K.; Monfort, S.L.; Seal, U.S. Genome Resource Banks Living collections for biodiversity conservation. Bioscience 1997, 47, 689–698. [Google Scholar]
- Roldan, E.R.S.; Garde, J.J. Biotecnología de la reproducción y conservación de especies en peligro de extinción. In Los Retos Medioambientales del Siglo XXI. La Conservación de la Biodiversidad en España; Gomendio, M., Ed.; CSIC-FBBVA: Madrid, Spain, 2004; pp. 283–303. [Google Scholar]
- Pukazhenthi, B.; Santymire, R.; Crosier, A.; Howard, J.G.; Wildt, D.E. Challenges in cryopreserving endangered mammal spermaozoa: Morphology and the value of acrosomal integrity as markers of cryo-survival. In Spermatology; Roldan, E.R.S., Gomendio, M., Eds.; Nottingham University Press: Nottingham, UK, 2007; pp. 433–446. [Google Scholar]
- Swanson, W.F.; Stoops, M.A.; Magarey, G.M.; Herrick, J.R. Sperm cryopreservation in endangered felids: Developing linkage of in situ-ex situ populations. In Spermatology; Roldan, E.R.S., Gomendio, M., Eds.; Nottingham University Press: Nottingham, UK, 2007; pp. 417–432. [Google Scholar]
- Comizzoli, P.; Amelkina, O.; Chavez, D.R.; Rowlison, T.R.; Lee, P.C. Current knowledge in the biology of gametes and embryos from Carnivora. Theriogenology 2023, 196, 254–263. [Google Scholar] [PubMed]
- Wildt, D.E.; Schiewe, M.C.; Schmidt, P.M.; Goodrowe, K.L.; Howard, J.G.; Phillips, L.G.; O’Brien, S.J.; Bush, M. Developing animals models system for embryo technologies in rare and endangered wildlife. Theriogenology 1986, 25, 33–51. [Google Scholar]
- Swanson, W.F. Research in non-domestic species: Experiences in Reproductive physiology research for conservation of endangered felids. ILAR J. 2003, 4, 307–316. [Google Scholar]
- Pukazhenthi, B.; Comizzoli, P.; Travis, A.J.; Wildt, D.E. Applications of emerging technologies to the study and conservation of threatened and endangered species. Reprod. Fert. Dev. 2006, 18, 77–90. [Google Scholar]
- Farstad, W. Current state of biotechnology in canine and feline reproduction. Anim. Reprod. Sci. 2000, 60–61, 375–387. [Google Scholar]
- Pope, C.E. Embryo technology in conservation efforts for endangered felids. Theriogenology 2000, 53, 163–174. [Google Scholar]
- Luvoni, G.C.; Kalchschmidt, E.; Leoni, S.; Ruggiero, C. Conservation of feline semen. Part I: Cooling and freezing protocols. J. Feline Med. Surg. 2003, 5, 203–208. [Google Scholar]
- Luvoni, G.C.; Kalchschmidt, E.; Marinoni, G. Conservation of feline semen. Part II: Cold-induced damages on spermatozoal dertilizing ability. J. Feline Med. Surg. 2003, 5, 257–263. [Google Scholar]
- Villaverde, A.I.; Melo, C.M.; Martin, I.; Ferreira, T.H.; Papa, F.O.; Taconeli, C.A.; Lopes, M.D. Comparison of efficiency between two artificial insemination methods using frozen-thawed semen in domestic cat (Felis catus): Artificial insemination in domestic cats. Anim. Reprod. Sci. 2009, 114, 434–442. [Google Scholar] [CrossRef]
- Johnston, L.A.; Donoghue, A.M.; O’Brien, S.J.; Wildt, D.E. Rescue and Maduration in vitro of follicular oocytes collected from nondomestic felid species. Biol. Reprod. 1991, 45, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Goodrowe, K.L.; Hay, M. Characteristics and zona binding ability of fresh and cooled domestic cat epididymal spermatozoa. Theriogenology 1993, 40, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Lengwinat, T.; Blottner, S. In vitro fertilization of follicular oocytes of domestic cat using fresh and cryopreserved epididymal spermatozoa. Anim. Reprod. Sci. 1994, 35, 291–301. [Google Scholar] [CrossRef]
- Kashiwazaki, N.; Yamaguchi, R.; Uesugi, R.; Hishiyama, N.; Kim, M.; Nakatsukasa, E.; Kojima, Y.; Okuda, Y.; Hisamatsu, S.; Inomata, T.; et al. Sperm motility, plasma membrane integrity, and binding capacity to homologous zona pellucida of cryopreserved epididymal spermatozoa in the domestic cat. J. Reprod. Dev. 2005, 51, 735–739. [Google Scholar]
- Zambelli, D.; Prati, F.; Cunto, M.; Iacono, E.; Merlo, B. Quality and in vitro fertilizing ability of cryopreserved cat spermatozoa obtained by urethral catheterization after medetomidine administration. Theriogenology 2008, 69, 485–490. [Google Scholar] [CrossRef]
- Zambelli, D.; Merlo, B.; Iacono, E.; Prati, F.; Belluzzi, S. Fertilizing ability of electro-ejaculated cryopreserved semen in the domestic cat. Reprod. Domest. Anim. 2006, 41, 137–141. [Google Scholar]
- Bogliolo, L.; Leoni, G.; Ledda, S.; Naitana, S.; Zedda, M.; Carluccio, A.; Pau, S. Intracytoplasmic sperm injection of in vitro matured oocytes of domestic cats with frozen-thawed epididymal spermatozoa. Theriogenology 2001, 56, 955–967. [Google Scholar]
- Tsutsui, T. Artificial insemination in domestic cats (Felis catus). Theriogenology 2006, 66, 122–125. [Google Scholar]
- Tsutsui, T.; Wada, M.; Aznai, M.; Hori, T. Artificial Insemination with frozen epididymal sperm in cats. J. Vet. Med. Sci. 2002, 65, 397–399. [Google Scholar]
- Tsutsui, T.; Tanaka, A.; Takagi, Y.; Nakagawa, K.; Fujimoto, Y.; Murai, M.; Anzai, M.; Hori, T. Unilateral intrauterine horn insemination of frozen semen in cats. J. Vet. Med. Sci. 2000, 62, 1247–1251. [Google Scholar]
- Chatdarong, K.; Axnér, E.; Manee-In, S.; Thuwanut, P.; Linde-Forsberg, C. Pregnancy in the domestic cat after vaginal or transcervical insemination with fresh and frozen semen. Theriogenology 2007, 68, 1326–1333. [Google Scholar] [PubMed]
- Howard, J.G.; Wildt, D.E. Approaches and efficacy of artificial insemination in felids and mustelids. Theriogenology 2009, 71, 130–148. [Google Scholar] [PubMed]
- Brusentsev, E.; Kizilova, E.; Mokrousova, V.; Kozhevnikova, V.; Rozhkova, I.; Amstislavsky, S. Characteristics and fertility of domestic cat epididymal spermatozoa cryopreserved with two different freezing media. Theriogenology 2018, 110, 148–152. [Google Scholar] [PubMed]
- Macente, B.I.; Gutierrez, R.R.; Apparício, M.; Balieiro, C.C.; Mansano, C.F.M.; Pereira, M.M.; Borges-Silva, J.C.; Pires-Buttler, E.A.; Galvão, A.L.B.; Toniollo, G.H.; et al. Cat epididymal semen cryopreserved with and without vitamin E: Effect on sperm parameters and lipid peroxidation. Anim. Reprod. 2018, 15, 1193–1198. [Google Scholar]
- Barbosa, B.S.; Izzo, R.G.; Silva, H.V.R.; Nunes, T.G.P.; Brito, B.F.; Silva, T.F.P.D.; Silva, L.D.M.D. Recovery and cryopreservation of epididymal sperm from domestic cat using powdered coconut water (ACP-117c) and TRIS extenders. Cryobiology 2020, 92, 103–108. [Google Scholar]
- Buranaamnuay, K. Effect of different permeable cryoprotectants on the quality of cat epididymal spermatozoa. Cryo Lett. 2020, 41, 237–244. [Google Scholar]
- Leibo, S.P.; Bradley, L. Comparative cryobiology of mammalian spermatozoa. In The Male Gamete: From Basic Science to Clinical Applications; Gagnon, C., Ed.; Cache River Press: Vienna, IL, USA, 1999; pp. 501–516. [Google Scholar]
- Holt, W.V. Fundamental aspects of sperm cryobiology: The importance of species and individual differences. Theriogenology 2000, 53, 47–58. [Google Scholar]
- Fuchs, B.; Jakop, U.; Göritz, F.; Hermes, R.; Hildebrandt, T.; Schiller, J.; Müller, K. MALDI-TOF “fingerprint” phospholipid mass spectra allow the differentiation between ruminantia and feloideae spermatozoa. Theriogenology 2009, 71, 568–575. [Google Scholar]
- Pope, C.E.; Turner, J.L.; Quatman, S.P.; Dresser, B.L. Semen storage in the domestic felid: A comparison of cryopreservation methods and storage temperature. Biol. Reprod. 1991, 44, 117–126. [Google Scholar]
- Hermansson, U.; Axnér, E. Epididymal and ejaculated cat spermatozoa are resistant to cold shock but egg yolk promotes sperm longevity during cold storage at 4 °C. Theriogenology 2007, 67, 1239–1248. [Google Scholar]
- Zambelli, D.; Canappele, B.; Castagnetti, C.; Belluzzi, S. Cryopreservation of cat semen in straws: Comparison of five different freezing rates. Reprod. Dom. Anim. 2002, 37, 310–313. [Google Scholar]
- Minter, L.J.; Pinto, C.R.F.; Davis, M.; Kozink, D.M. Influence of extender and packaging on post-thaw survival of epididymal cat spermatozoa. Theriogenology 2007, 68, 501–502. [Google Scholar]
- Buranaamnuay, K. Protocols for sperm cryopreservation in the domestic cat: A review. Anim. Reprod. Sci. 2017, 183, 56–65. [Google Scholar] [PubMed]
- Jelinkova, K.; Vitasek, R.; Novotny, R.; Bartoskova, A. A comparison of quality parameters of fresh feline ejaculates collected by three different collection techniques. Reprod. Domest. Anim. 2018, 53, 1068–1074. [Google Scholar]
- Tebet, J.M.; Martins, M.I.; Chirinea, V.H.; Souza, F.F.; Campagnol, D.; Lopes, M.D. Cryopreservation effects on domestic cat epididymal versus electroejaculated spermatozoa. Theriogenology 2006, 66, 1629–1632. [Google Scholar]
- Klaus, C.; Eder, S.; Franz, C.; Müller, K. Successful cryopreservation of domestic cat (Felis catus) epididymal sperm after slow equilibration to 15 or 10 °C. Reprod. Domest. Anim. 2016, 51, 195–203. [Google Scholar]
- Chatdarong, K. Retained fertilizing capability in cryopreserved feline spermatozoa. Reprod. Domest. Anim. 2017, 52, 261–264. [Google Scholar]
- Müller, K.; Eder, S.; Jakop, U.; Schiller, J.; Müller, P.; Bashawat, M. Assisted reproduction for felid species conservation-Sperm competences at risk. Reprod. Domest. Anim. 2020, 55, 55–60. [Google Scholar]
- Eder, S.; Bailey, L.D.; Müller, K. Equilibration in freezing extender alters in vitro sperm-oviduct binding in the domestic cat (Felis catus). Theriogenology 2020, 149, 79–87. [Google Scholar]
- Howard, J.G.; Bush, M.; Wildt, D.E. Semen collection, analysis and cryopreservation in nondomestic mammals. In Current Therapy in Theriogenology II; Morrow, D., Ed.; WB Saunders: Philadelphia, PA, USA, 1986; pp. 1047–1053. [Google Scholar]
- Howard, J.G. Feline semen analysis and artifitial insemination. In Kirk’s Current Veterinary Therapy. XI Small Animal Practice; Kirk, R.W., Bonagura, J.D., Eds.; WB Saunders: Philadelphia, PA, USA, 1992; pp. 929–934. [Google Scholar]
- Larson, J.L.; Miller, D.J. Simple histochemical stain for acrosomes on sperm from several species. Mol. Reprod. Dev. 1999, 52, 445–449. [Google Scholar]
- Pukazhenthi, B.; Pelican, K.; Wildt, D.E.; Howard, J.G. Sensitivity of the domestic cat (Felis catus) sperm from normospermic versus teratospermic donors to cold-induced acrosomal damage. Biol. Reprod. 1999, 61, 135–141. [Google Scholar] [PubMed]
- Villaverde, A.I.; Martins, M.I.; Castro, V.B.; Lopes, M.D. Morphological and functional characteristics of chilled semen obtained from domestic feline epididymides (Felis catus). Theriogenology 2006, 66, 1641–1644. [Google Scholar] [PubMed]
- Watson, P.F. The effects of cold shock on sperm cell membranes. In Effects of Low Temperatures on Biological Membranes; Morris, G.J., Clarke, A., Eds.; Academic Press: Londres, UK, 1981; pp. 189–218. [Google Scholar]
- Baudi, D.L.K.; Jewgenow, K.; Pukazhenthi, B.; Spercoski, K.M.; Santos, A.S.; Reghelin, A.L.S.; Candido, M.V.; Javorouski, M.L.; Müller, G.; Morais, R.N. Influence of cooling rate on the ability of frozen-thawed sperm to bind to heterologous zona pellucida, as assessed by competitive in vitro binding assay in the ocelot (Leopardus pardalis) and tigrina (Leopardus tigrinus). Theriogenology 2008, 69, 204–211. [Google Scholar] [PubMed]
- Gañán, N.; Gomendio, M.; Roldan, E.R.S. Effect of storage of domestic cat (Felis catus) epididymides at 5 °C on sperm quality and cryopreservation. Theriogenology 2009, 72, 1268–1277. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Status of Sample |
|---|---|
| 1 | Fresh from the epididymis |
| 2 | After refrigeration, before freezing |
| 3 | Soon after thawing |
| 4 | 90 min incubation post-thawing |
| 5 | 150 min incubation post-thawing |
| 6 | 210 min incubation post-thawing |
| 7 | 270 min incubation post-thawing |
| Parameter | Mean ± SEM |
|---|---|
| Total number of spermatozoa (×106) | 28.15 ± 3.23 |
| Normal spermatozoa (%) | 30.95 ± 1.98 |
| Head abnormalities (%) | 1.41 ± 0.21 |
| Medium piece abnormalities (%) | 27.72 ± 1.63 |
| Principal and terminal piece abnormalities (%) | 36.70 ± 2.15 |
| Coiled tail (%) | 4.62 ± 0.77 |
| Cytoplasmic droplet (%) | 27.89 ± 2.38 |
| Intact acrosomes (%) | 57.69 ± 1.40 |
| Double Interactions | Triple Interaction | |||||
|---|---|---|---|---|---|---|
| SMI | F | p | F | p | ||
| 1. Time | 2. Refrigeration | 3. Storage system | (1–2) 0.44 | 0.81 | (1.2.4) 2.58 | 0.07 |
| (1–4) 1.24 | 0.34 | |||||
| (2–4) 0.09 | 0.77 | |||||
| 4. Straw’s loading | (1–2) 1.83 | 0.15 | (1.2.5) 0.57 | 0.72 | ||
| (1–5) 0.67 | 0.65 | |||||
| (2–5) 1.81 | 0.19 | |||||
| Double Interactions | Triple Interaction | |||||
|---|---|---|---|---|---|---|
| % Intact Acrosomes | F | p | F | p | ||
| 1. Time | 2. Refrigeration | 3. Storage system | (1–2) 0.70 | 0.51 | (1.2.4) 0.18 | 0.84 |
| (1–4) 0.43 | 0.06 | |||||
| (2–4) 0.26 | 0.62 | |||||
| 4. Straw’s loading | (1–2) 0.32 | 0.63 | (1.2.5) 0.49 | 0.62 | ||
| (1–5) 0.53 | 0.60 | |||||
| (2–5) 0.02 | 0.88 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gañán, N.; Sanchez-Rodriguez, A.; Roldan, E.R.S. Factors Affecting Cryopreservation of Domestic Cat (Felis catus) Epididymal Spermatozoa. Animals 2025, 15, 949. https://doi.org/10.3390/ani15070949
Gañán N, Sanchez-Rodriguez A, Roldan ERS. Factors Affecting Cryopreservation of Domestic Cat (Felis catus) Epididymal Spermatozoa. Animals. 2025; 15(7):949. https://doi.org/10.3390/ani15070949
Chicago/Turabian StyleGañán, Natalia, Ana Sanchez-Rodriguez, and Eduardo R. S. Roldan. 2025. "Factors Affecting Cryopreservation of Domestic Cat (Felis catus) Epididymal Spermatozoa" Animals 15, no. 7: 949. https://doi.org/10.3390/ani15070949
APA StyleGañán, N., Sanchez-Rodriguez, A., & Roldan, E. R. S. (2025). Factors Affecting Cryopreservation of Domestic Cat (Felis catus) Epididymal Spermatozoa. Animals, 15(7), 949. https://doi.org/10.3390/ani15070949