
The Potential of the Inclusion of Prosopis farcta Extract in the Diet on the Growth Performance, Immunity, Digestive Enzyme Activity, and Oxidative Status of the Common Carp, Cyprinus carpio, in Response to Ammonia Stress

 , , , and
, , , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Extraction of the Plant Material
2.2. Preparation of the Diets and Administration of the Extract
2.3. The Fish and Experimental Design
2.4. Fish Biometry and Sampling
2.5. Ambient Ammonia Exposure
2.6. Analysis
2.6.1. Digestive and Antioxidant Enzymes
2.6.2. Biochemical Markers
2.6.3. Innate Immune Responses
2.7. Statistical Analysis
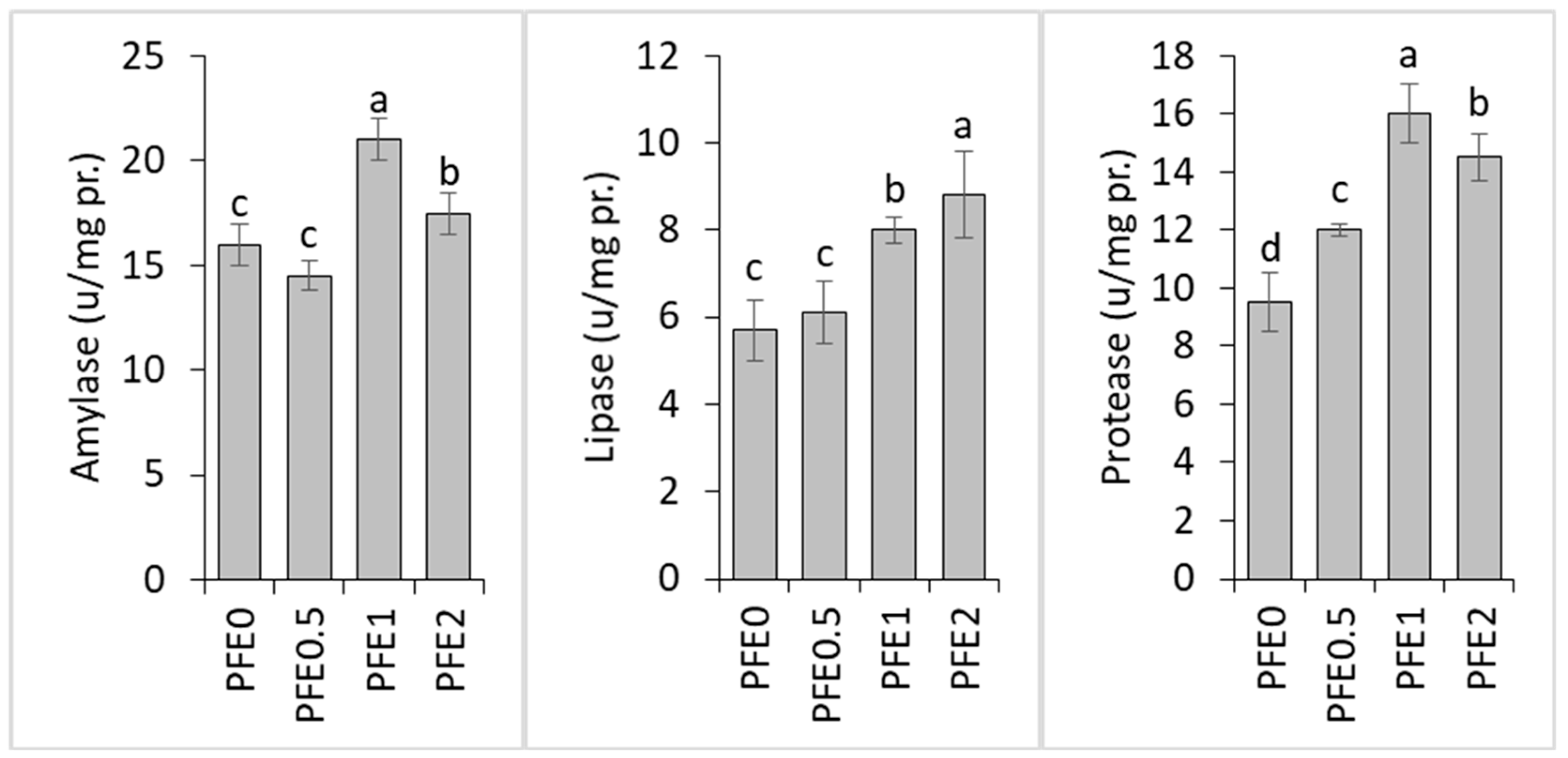
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022: Towards Blue Transformation; FAO: Roma, Italy, 2022. [Google Scholar]
- Li, L.; Shen, Y.; Yang, W.; Xu, X.; Li, J. Effect of different stocking densities on fish growth performance: A meta-analysis. Aquaculture 2021, 544, 737152. [Google Scholar]
- Hoseini, S.M.; Barbieri, E.; Aydın, B.; Yousefi, M. A review of dietary approaches for ammonia toxicity remediation in fish. Aquac. Int. 2024, 32, 5639–5675. [Google Scholar]
- Reverter, M.; Tapissier-Bontemps, N.; Sarter, S.; Sasal, P.; Caruso, D. Moving towards more sustainable aquaculture practices: A meta-analysis on the potential of plant-enriched diets to improve fish growth, immunity and disease resistance. Rev. Aquac. 2020, 13, 537–555. [Google Scholar]
- Ronel, M.; Lev-Yadun, S. Spiny plants in the archaeological record of Israel. J. Arid. Environ. 2009, 73, 754–761. [Google Scholar] [CrossRef]
- Noroozi, R.; Sadeghi, E.; Yousefi, H.; Taheri, M.; Sarabi, P.; Dowati, A.; Ayatallahi, S.A.; Noroozi, R.; Ghafouri-Fard, S. Wound healing features of Prosopis farcta: In vitro evaluation of antibacterial, antioxidant, proliferative and angiogenic properties. Gene Rep. 2019, 17, 100482. [Google Scholar]
- Direkvand-Moghadam, F.; Ghasemi-Seyed, V.; Abdali-Mashhadi, A.-R.; Lotfi, A.; Direkvand-Moghadam, A.; Delpisheh, A. Extraction and measurement of the Quercetin flavonoid of Prosopis farcta in Khouzestan climatic condition. Future Nat. Prod. 2015, 1, 29–35. [Google Scholar]
- Azab, A. Antifungal and anti-termite activities, total phenolic content of Prosopis farcta extracts; attempts to develop weed biocontrol method against it. Eur. Chem. Bull. 2018, 7, 293–302. [Google Scholar] [CrossRef]
- Jahromi, M.A.F.; Etemadfard, H.; Zebarjad, Z. Antimicrobial and antioxidant characteristics of volatile components and ethanolic fruit extract of Prosopis farcta (Bank & Soland.). Trends Pharm. Sci. 2018, 4, 177–186. [Google Scholar]
- Sanchooli, N.; Rigi, M. The effect of plant extracts Prosopis farcta, Datura stramunium and Calotropis procera against three species of fish pathogenic bacteria. J. Vet. Res. 2015, 70, 52–63. [Google Scholar]
- Gholamalipour Alamdari, E.; Taleghani, A. New bioactive compounds characterized by liquid chromatography–mass spectrometry and gas chromatography–mass spectrometry in hydro-methanol and petroleum ether extracts of Prosopis farcta (Banks & Sol.) J. F. Macbr weed. J. Mass Spectrom. 2022, 57, e4884. [Google Scholar]
- Ghaddar, B.; Gence, L.; Veeren, B.; Bringart, M.; Bascands, J.-L.; Meilhac, O.; Diotel, N. Aqueous extract of Psiloxylon mauritianum, rich in gallic acid, prevents obesity and associated deleterious effects in zebrafish. Antioxidants 2022, 11, 1309. [Google Scholar] [CrossRef]
- Lee, P.-T.; Chen, H.-Y.; Liao, Z.-H.; Huang, H.-T.; Chang, T.-C.; Huang, C.-T.; Lee, M.-C.; Nan, F.-H. Effects of three medicinal herbs Bidens pilosa, Lonicera japonica, and Cyathula officinalis on growth and non-specific immune responses of cobia (Rachycentron canadum). Fish Shellfish. Immunol. 2020, 106, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-F.; Chen, C.; Jiang, X.-Y.; Shen, J.-L.; Ling, F.; Li, P.-F.; Wang, G.-X. Luteolin in Lonicera japonica inhibits the proliferation of white spot syndrome virus in the crayfish Procambarus clarkii. Aquaculture 2022, 550, 737852. [Google Scholar] [CrossRef]
- Yang, G.; Yu, R.; Geng, S.; Xiong, L.; Yan, Q.; Kumar, V.; Wen, C.; Peng, M. Apple polyphenols modulates the antioxidant defense response and attenuates inflammatory response concurrent with hepatoprotective effect on grass carp (Ctenopharyngodon idellus) fed low fish meal diet. Aquaculture 2021, 534, 736284. [Google Scholar] [CrossRef]
- Almarri, S.H.; Khalil, A.A.; Mansour, A.T.; El-Houseiny, W. Antioxidant, immunostimulant, and growth-promoting effects of dietary Annona squamosa leaf extract on Nile tilapia, Oreochromis niloticus, and its tolerance to thermal stress and Aeromonas sobria infection. Animals 2023, 13, 746. [Google Scholar] [CrossRef]
- Rajabiesterabadi, H.; Yousefi, M.; Hoseini, S.M. Enhanced haematological and immune responses in common carp Cyprinus carpio fed with olive leaf extract-supplemented diets and subjected to ambient ammonia. Aquac. Nutr. 2020, 26, 763–771. [Google Scholar] [CrossRef]
- Sokooti, R.; Dezfoulnejad, M.C.; Javaheri baboli, M. Effects of olive leaf extract (Olea europaea Leecino) on growth, haematological parameters, immune system and carcass composition in common carp (Cyprinus carpio). Aquac. Res. 2021, 52, 2415–2423. [Google Scholar] [CrossRef]
- Taheri Mirghaed, A.; Fayaz, S.; Hoseini, S.M. Dietary 1, 8-cinoele affects serum enzymatic activities and immunological characteristics in common carp (Cyprinus carpio) exposed to ambient ammonia. Aquac. Res. 2018, 50, 146–153. [Google Scholar] [CrossRef]
- Emerson, K.; Russo, R.C.; Lund, R.E.; Thurston, R.V. Aqueous ammonia equilibrium calculations: Effect of pH and temperature. J. Fish. Board Can. 1975, 32, 2379–2383. [Google Scholar] [CrossRef]
- Iijima, N.; Tanaka, S.; Ota, Y. Purification and characterization of bile salt-activated lipase from the hepatopancreas of red sea bream, Pagrus major. Fish Physiol. Biochem. 1998, 18, 59–69. [Google Scholar] [CrossRef]
- Bernfeld, P. [17] Amylases, α and β. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1955; Volume 1, pp. 149–158. [Google Scholar]
- Iversen, S.L.; Jørgensen, M.H. Azocasein assay for alkaline protease in complex fermentation broth. Biotechnol. Tech. 1995, 9, 573–576. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Ghafarifarsani, H.; Hoseinifar, S.H.; Adorian, T.J.; Ferrigolo, F.R.G.; Raissy, M.; Van Doan, H. The effects of combined inclusion of Malvae sylvestris, Origanum vulgare, and Allium hirtifolium boiss for common carp (Cyprinus carpio) diet: Growth performance, antioxidant defense, and immunological parameters. Fish Shellfish. Immunol. 2021, 119, 670–677. [Google Scholar] [CrossRef]
- Yousefi, M.; Adineh, H.; Sedaghat, Z.; Yilmaz, S.; Elgabry, S.E. Effects of dietary costmary (Tanacetum balsamita) essential oil on growth performance, digestive enzymes’ activity, immune responses and subjected to ambient ammonia of common carp Cyprinus carpio. Aquaculture 2023, 569, 739347. [Google Scholar] [CrossRef]
- Ellis, A.E. Lysozyme assays. In Techniques in Fish Immunology; Stolen, J.S., Ed.; SOS Publication: Fair Haven, NJ, USA, 1990; pp. 101–103. [Google Scholar]
- Yano, T. Assays of hemolytic complement activity. In Techniques in Fish Immunology; Stolen, J.S., Ed.; SOS Publication: Fair Haven, NJ, USA, 1992; pp. 131–141. [Google Scholar]
- Siwicki, A.; Anderson, D. Nonspecific defense mechanisms assay in fish: II. Potential killing activity of neutrophils and macrophages, lysozyme activity in serum and organs and total immunoglobulin level in serum. In Fish Disease Diagnosis and Prevention Methods; Siwicki, A., Anderson, D., Waluga, J., Eds.; Wydawnictwo Instytutu Rybactwa Srodladowego: Olsztyn, Poland, 1993; pp. 105–112. [Google Scholar]
- Citarasu, T. Herbal biomedicines: A new opportunity for aquaculture industry. Aquac. Int. 2010, 18, 403–414. [Google Scholar] [CrossRef]
- Pu, H.; Li, X.; Du, Q.; Cui, H.; Xu, Y. Research progress in the application of Chinese herbal medicines in aquaculture: A review. Engineering 2017, 3, 731–737. [Google Scholar] [CrossRef]
- Ciji, A.; Akhtar, M.S. Stress management in aquaculture: A review of dietary interventions. Rev. Aquac. 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- Mahmoud, H.K.; Reda, F.M.; Alagawany, M.; Farag, M.R. Ameliorating deleterious effects of high stocking density on Oreochromis niloticus using natural and biological feed additives. Aquaculture 2021, 531, 735900. [Google Scholar] [CrossRef]
- Hajirezaee, S.; Rafieepour, A.; Rahimi, R.; Shafiei, S. Effects of gingko, Ginkgo biloba extract on metabolic hormones, liver histology, and growth parameters of rainbow trout, Oncorhynchus mykiss exposed to diazinon. Toxin Rev. 2021, 40, 632–644. [Google Scholar] [CrossRef]
- Awad, E.; Awaad, A. Role of medicinal plants on growth performance and immune status in fish. Fish Shellfish. Immunol. 2017, 67, 40–54. [Google Scholar] [PubMed]
- Sadoul, B.; Geffroy, B. Measuring cortisol, the major stress hormone in fishes. J. Fish Biol. 2019, 94, 540–555. [Google Scholar] [PubMed]
- Dawood, M.A.O.; Sewilam, H. The combined effects of salinity and ammonia on the growth behavior, stress-related markers, and hepato-renal function of common carp (Cyprinus carpio). J. Exp. Zool. Part A Ecol. Integr. Physiol. 2023, 339, 74–82. [Google Scholar]
- Yousefi, M.; Vatnikov, Y.A.; Kulikov, E.V.; Plushikov, V.G.; Drukovsky, S.G.; Hoseinifar, S.H.; Van Doan, H. The protective effects of dietary garlic on common carp (Cyprinus carpio) exposed to ambient ammonia toxicity. Aquaculture 2020, 526, 735400. [Google Scholar]
- Topić Popović, N.; Čižmek, L.; Babić, S.; Strunjak-Perović, I.; Čož-Rakovac, R. Fish liver damage related to the wastewater treatment plant effluents. Environ. Sci. Pollut. Res. 2023, 30, 48739–48768. [Google Scholar]
- Hoseini, S.M.; Yousefi, M.; Hoseinifar, S.H.; Van Doan, H. Antioxidant, enzymatic and hematological responses of common carp (Cyprinus carpio) fed with myrcene- or menthol-supplemented diets and exposed to ambient ammonia. Aquaculture 2019, 506, 246–255. [Google Scholar] [CrossRef]
- Agirman, E.; Celik, I.; Dogan, A. Consumption of the Syrian mesquite plant (Prosopis farcta) fruit and seed lyophilized extracts may have both protective and toxic effects in STZ-induced diabetic rats. Arch. Physiol. Biochem. 2022, 128, 887–896. [Google Scholar] [CrossRef]
- Safari, A.; Sharifzadeh, M.; Hassanzadeh, G. The protective effect of a standardized hydroalcoholic extract of Prosopis farcta (Banks & Sol.) JF Macbr. fruit in a rat model for experimental ulcerative colitis. Tradit Med. Res. 2021, 6, 40–47. [Google Scholar]
- Rashidian, G.; Kajbaf, K.; Prokić, M.D.; Faggio, C. Extract of common mallow (Malvae sylvestris) enhances growth, immunity, and resistance of rainbow trout (Oncorhynchus mykiss) fingerlings against Yersinia ruckeri infection. Fish Shellfish. Immunol. 2020, 96, 254–261. [Google Scholar]
- Yousefi, M.; Vatnikov, Y.A.; Kulikov, E.V.; Ahmadifar, E.; Mirghaed, A.T.; Hoseinifar, S.H.; Van Doan, H. Effects of dietary Hibiscus sabdariffa supplementation on biochemical responses and inflammatory-related genes expression of rainbow trout, Oncorhynchus mykiss, to ammonia toxicity. Aquaculture 2021, 533, 736095. [Google Scholar]
- Hegazi, M.M.; Attia, Z.I.; Ashour, O.A. Oxidative stress and antioxidant enzymes in liver and white muscle of Nile tilapia juveniles in chronic ammonia exposure. Aquat. Toxicol. 2010, 99, 118–125. [Google Scholar] [PubMed]
- Santana, M.S.; de Melo, G.D.; Sandrini-Neto, L.; Di Domenico, M.; Prodocimo, M.M. A meta-analytic review of fish antioxidant defense and biotransformation systems following pesticide exposure. Chemosphere 2022, 291, 132730. [Google Scholar] [PubMed]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [PubMed]
- Islam, M.N.; Rauf, A.; Fahad, F.I.; Emran, T.B.; Mitra, S.; Olatunde, A.; Shariati, M.A.; Rebezov, M.; Rengasamy, K.R.R.; Mubarak, M.S. Superoxide dismutase: An updated review on its health benefits and industrial applications. Crit. Rev. Food Sci. Nutr. 2022, 62, 7282–7300. [Google Scholar]
- Ebrahimzadeh Mousavi, H.; Mirghaed, A.T.; Hoseini, S.M.; Ghelichpour, M.; Moghaddam, A.A.; Gharavi, B.; Aydın, B. Myrcene as water conditioner, stress-reducing and antioxidant agent in transportation of common carp, Cyprinus carpio, with plastic bags. Aquac. Rep. 2023, 28, 101458. [Google Scholar]
- Ghanbari, E.; Khazaei, M.; Yousefzaei, F. The restorative effect of Prosopis farcta on fertility parameters and antioxidant status in diabetic rats. J. Babol Univ. Med. Sci. 2017, 19, 53–60. [Google Scholar]

{kind=link}
| Ingredients (g/kg Diet) | PFE0 | PFE0.5 | PFE1 | PFE2 |
|---|---|---|---|---|
| Kilka fishmeal (72.72% CP) | 100 | 100 | 100 | 100 |
| Poultry slaughterhouse by-product (58.5% CP) | 200 | 200 | 200 | 200 |
| Soybean meal (46.78% CP) | 230 | 230 | 230 | 230 |
| Wheat meal (14.52% CP) | 389 | 384 | 379 | 369 |
| Corn flour (7.77% CP) | 50.0 | 50.0 | 50.0 | 50.0 |
| Fish oil | 7.00 | 7.00 | 7.00 | 7.00 |
| Soybean oil | 7.00 | 7.00 | 7.00 | 7.00 |
| Lysine | 7.00 | 7.00 | 7.00 | 7.00 |
| Methionine | 5.00 | 5.00 | 5.00 | 5.00 |
| Vitamin premix 1 | 2.50 | 2.50 | 2.50 | 2.50 |
| Mineral premix 1 | 2.50 | 2.50 | 2.50 | 2.50 |
| PFE | - | 5.00 | 10.0 | 20.0 |
| Total | 1000 | 1000 | 1000 | 1000 |
| Dry matter | 89.22 | 89.17 | 89.13 | 89.04 |
| Crude protein (%) | 35.30 | 35.30 | 35.29 | 35.27 |
| Crude fat (%) | 5.78 | 5.78 | 5.78 | 5.78 |
| Crude ash (%) | 5.88 | 5.88 | 5.88 | 5.87 |
| Energy (kcal/kg) | 4049.19 | 4047.23 | 4045.27 | 4041.35 |
| PFE0 (Control) | PFE0.5 | PFE1 | PFE2 | p-Value | |
|---|---|---|---|---|---|
| IW (g) | 14.9 ± 0.41 | 14.9 ± 0.20 | 15.4 ± 0.44 | 15.2 ± 0.50 | 0.379 |
| FW (g) | 36.7 ± 1.79 b | 38.3 ± 0.75 b | 44.4 ± 1.15 a | 44.7 ± 2.23 a | <0.001 |
| WG (%) | 146 ± 3.59 c | 157 ± 1.75 b | 188 ± 1.77 a | 194 ± 5.15 a | <0.001 |
| SGR (%/d) | 1.50 ± 0.02 c | 1.58 ± 0.01 b | 1.76 ± 0.01 a | 1.80 ± 0.03 a | <0.001 |
| FI (g/fish) | 39.4 ± 0.24 a | 41.8 ± 0.34 b | 43.6 ± 0.15 c | 43.8 ± 0.22 c | <0.001 |
| FCR | 1.81 ± 0.10 a | 1.79 ± 0.03 a | 1.51 ± 0.05 b | 1.49 ± 0.09 b | 0.001 |
| Survival (%) | 100 | 100 | 100 | 100 |
| Cortisol (ng/mL) | Glucose (mg/dL) | AST (U/L) | ALT (U/L) | |
|---|---|---|---|---|
| Basal | ||||
| PFE0 | 57.87 ± 4.99 a | 80.06 ± 2.43 a | 278.47 ± 3.75 a | 14.28 ± 0.68 a |
| PFE0.5 | 49.34 ± 2.78 b | 75.40 ± 1.50 b | 254.58 ± 3.34 b | 13.36 ± 0.66 a |
| PFE1 | 47.42 ± 2.95 b | 71.21 ± 1.98 c | 241.69 ± 5.97 c | 10.23 ± 0.38 b |
| PFE2 | 47.71 ± 3.26 b | 69.62 ± 2.53 c | 238.80 ± 2.11 c | 11.24 ± 0.60 b |
| Stressed | ||||
| PFE0 | 135.97 ± 3.44 a* | 165.02 ± 2.83 a* | 399.95 ± 5.33 a* | 23.76 ± 0.73 a* |
| PFE0.5 | 91.25 ± 2.88 b* | 112.59 ± 1.73 b* | 300.01 ± 4.32 c* | 18.60 ± 0.69 b* |
| PFE1 | 80.21 ± 5.18 c* | 98.99 ± 3.63 c* | 327.19 ± 4.22 b* | 12.12 ± 0.54 d* |
| PFE2 | 76.07 ± 3.45 c* | 90.50 ± 2.95 d* | 322.37 ± 4.41 b* | 14.28 ± 0.71 c* |
| Two-way ANOVA | ||||
| PFE | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Stress | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Interaction | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| SOD (U/mL) | GPX (U/mL) | Catalase (U/mL) | MDA (µmol/L) | |
|---|---|---|---|---|
| Basal | ||||
| PFE0 | 48.52 ± 1.36 c | 194.18 ± 8.69 b | 75.99 ± 2.11 | 67.65 ± 2.31 a |
| PFE0.5 | 49.55 ± 0.53 c | 217.47 ± 5.84 a | 68.23 ± 1.55 | 63.53 ± 1.65 a |
| PFE1 | 57.30 ± 1.05 a | 222.60 ± 7.98 a | 64.72 ± 1.62 | 57.71 ± 1.38 b |
| PFE2 | 55.24 ± 0.93 b | 223.78 ± 6.58 a | 61.12 ± 1.20 | 53.11 ± 3.98 b |
| Stressed | ||||
| PFE0 | 45.58 ± 1.00 c* | 240.07 ± 4.27 b* | 81.66 ± 1.63 | 87.91 ± 2.30 a* |
| PFE0.5 | 55.57 ± 0.64 c* | 255.27 ± 6.13 a* | 73.36 ± 0.94 | 76.44 ± 1.68 a* |
| PFE1 | 60.11 ± 1.04 a* | 258.49 ± 5.12 a* | 66.21 ± 3.83 | 59.32 ± 3.31 b |
| PFE2 | 59.80 ± 0.90 b* | 299.71 ± 3.94 a* | 65.40 ± 2.02 | 62.32 ± 1.71 b* |
| Two-way ANOVA | ||||
| PFE | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Stress | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Interaction | p < 0.001 | p < 0.001 | p = 0.327 | p < 0.001 |
| Stress effects | ||||
| Basal | NA | NA | 67.52 ± 5.92 | NA |
| Stressed | NA | NA | 71.66 ± 7.14 | NA |
| Diet effects | ||||
| PFE0 | NA | NA | 78.83 ± 3.54 | NA |
| PFE0.5 | NA | NA | 70.80 ± 3.03 | NA |
| PFE1 | NA | NA | 65.47 ± 2.76 | NA |
| PFE2 | NA | NA | 63.26 ± 2.78 | NA |
| Lysozyme (U/mL) | Total Ig (mg/mL) | ACH50 (U/mL) | Total Protein (g/dL) | Albumin (g/dL) | |
|---|---|---|---|---|---|
| Basal | |||||
| PFE0 | 34.96 ± 0.72 c | 15.70 ± 0.42 | 130.00 ± 4.08 | 2.58 ± 0.13 | 1.02 ± 0.03 |
| PFE0.5 | 37.31 ± 0.69 b | 19.58 ± 0.45 | 136.03 ± 3.90 | 3.24 ± 0.07 | 1.33 ± 0.06 |
| PFE1 | 39.05 ± 0.89 a | 20.66 ± 1.01 | 137.24 ± 2.12 | 3.71 ± 0.16 | 1.39 ± 0.04 |
| PFE2 | 38.14 ± 0.56 ab | 19.78 ± 0.87 | 141.62 ± 4.05 | 3.46 ± 0.12 | 1.38 ± 0.05 |
| Stressed | |||||
| PFE0 | 28.73 ± 0.83 c* | 11.94 ± 0.83 | 125.51 ± 3.74 | 2.27 ± 0.14 | 0.99 ± 0.11 |
| PFE0.5 | 33.00 ± 1.04 b* | 16.48 ± 0.54 | 131.75 ± 3.27 | 2.94 ± 0.08 | 1.07 ± 0.06 |
| PFE1 | 35.16 ± 0.96 a* | 18.67 ± 0.37 | 135.02 ± 4.65 | 3.15 ± 0.03 | 1.17 ± 0.05 |
| PFE2 | 35.94 ± 0.92 ab* | 18.28 ± 0.80 | 137.88 ± 3.24 | 2.91 ± 0.09 | 1.24 ± 0.12 |
| Two-way ANOVA | |||||
| PFE | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Stress | p < 0.001 | p < 0.001 | p = 0.027 | p < 0.001 | p < 0.001 |
| Interaction | p = 0.007 | p = 0.051 | p = 0.950 | p = 0.101 | p = 0.082 |
| Stress effects | |||||
| Basal | NA | 18.93 ± 2.09 | 136.22 ± 5.33 | 3.24 ± 0.45 | 1.28 ± 0.16 |
| Stressed | NA | 16.34 ± 2.84 * | 132.54 ± 5.78 * | 2.82 ± 0.35 * | 1.12 ± 0.13 * |
| Diet effects | |||||
| PFE0 | NA | 13.82 ± 2.14 a | 127.75 ± 4.28 a | 2.43 ± 0.21 a | 1.01 ± 0.08 a |
| PFE0.5 | NA | 18.03 ± 1.76 b | 133.89 ± 3.99 b | 3.09 ± 0.18 b | 1.20 ± 0.15 b |
| PFE1 | NA | 19.66 ± 1.29 c | 136.13 ± 3.46 bc | 3.43 ± 0.32 c | 1.28 ± 0.13 bc |
| PFE2 | NA | 19.03 ± 1.11 c | 139.75 ± 3.87 c | 3.19 ± 0.32 b | 1.31 ± 0.12 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousefi, M.; Adineh, H.; Al Sulivany, B.S.A.; Gholamalipour Alamdari, E.; Yilmaz, S.; Mahboub, H.H.; Hoseini, S.M. The Potential of the Inclusion of Prosopis farcta Extract in the Diet on the Growth Performance, Immunity, Digestive Enzyme Activity, and Oxidative Status of the Common Carp, Cyprinus carpio, in Response to Ammonia Stress. Animals 2025, 15, 895. https://doi.org/10.3390/ani15060895
Yousefi M, Adineh H, Al Sulivany BSA, Gholamalipour Alamdari E, Yilmaz S, Mahboub HH, Hoseini SM. The Potential of the Inclusion of Prosopis farcta Extract in the Diet on the Growth Performance, Immunity, Digestive Enzyme Activity, and Oxidative Status of the Common Carp, Cyprinus carpio, in Response to Ammonia Stress. Animals. 2025; 15(6):895. https://doi.org/10.3390/ani15060895
Chicago/Turabian StyleYousefi, Morteza, Hossein Adineh, Basim S. A. Al Sulivany, Ebrahim Gholamalipour Alamdari, Sevdan Yilmaz, Heba H. Mahboub, and Seyyed Morteza Hoseini. 2025. "The Potential of the Inclusion of Prosopis farcta Extract in the Diet on the Growth Performance, Immunity, Digestive Enzyme Activity, and Oxidative Status of the Common Carp, Cyprinus carpio, in Response to Ammonia Stress" Animals 15, no. 6: 895. https://doi.org/10.3390/ani15060895
APA StyleYousefi, M., Adineh, H., Al Sulivany, B. S. A., Gholamalipour Alamdari, E., Yilmaz, S., Mahboub, H. H., & Hoseini, S. M. (2025). The Potential of the Inclusion of Prosopis farcta Extract in the Diet on the Growth Performance, Immunity, Digestive Enzyme Activity, and Oxidative Status of the Common Carp, Cyprinus carpio, in Response to Ammonia Stress. Animals, 15(6), 895. https://doi.org/10.3390/ani15060895