

Meningeal Lymphatic and Glymphatic Structures in a Pelagic Delphinid (Delphinus delphis)
 , , , , , ,
, , , , , ,
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
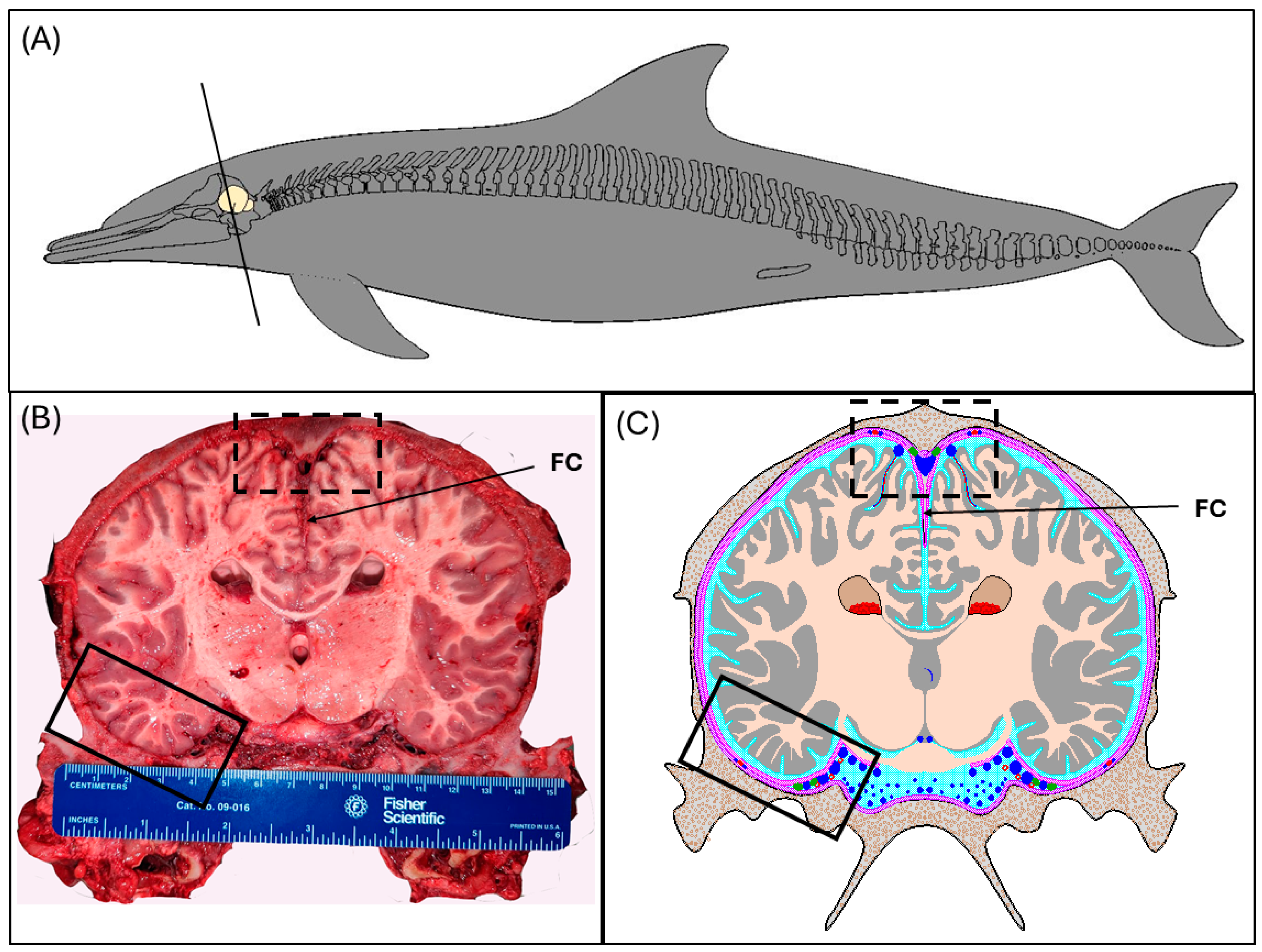
2.1. Specimens
2.2. Gross Dissection
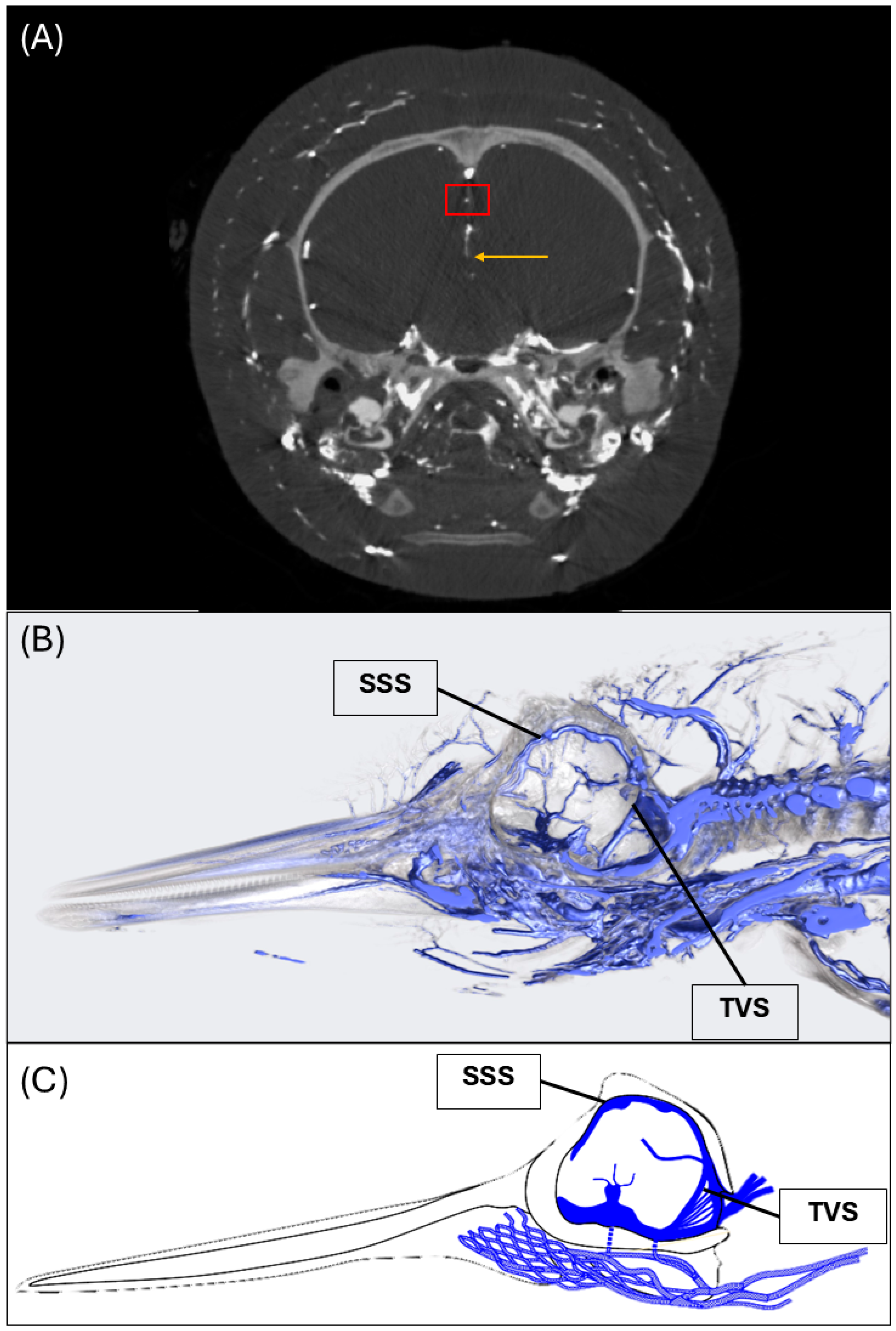
2.3. Computed Tomography
2.4. Histological Preparation
2.5. Histologic and Fluorescent Immunofluorescent Labeling
3. Results
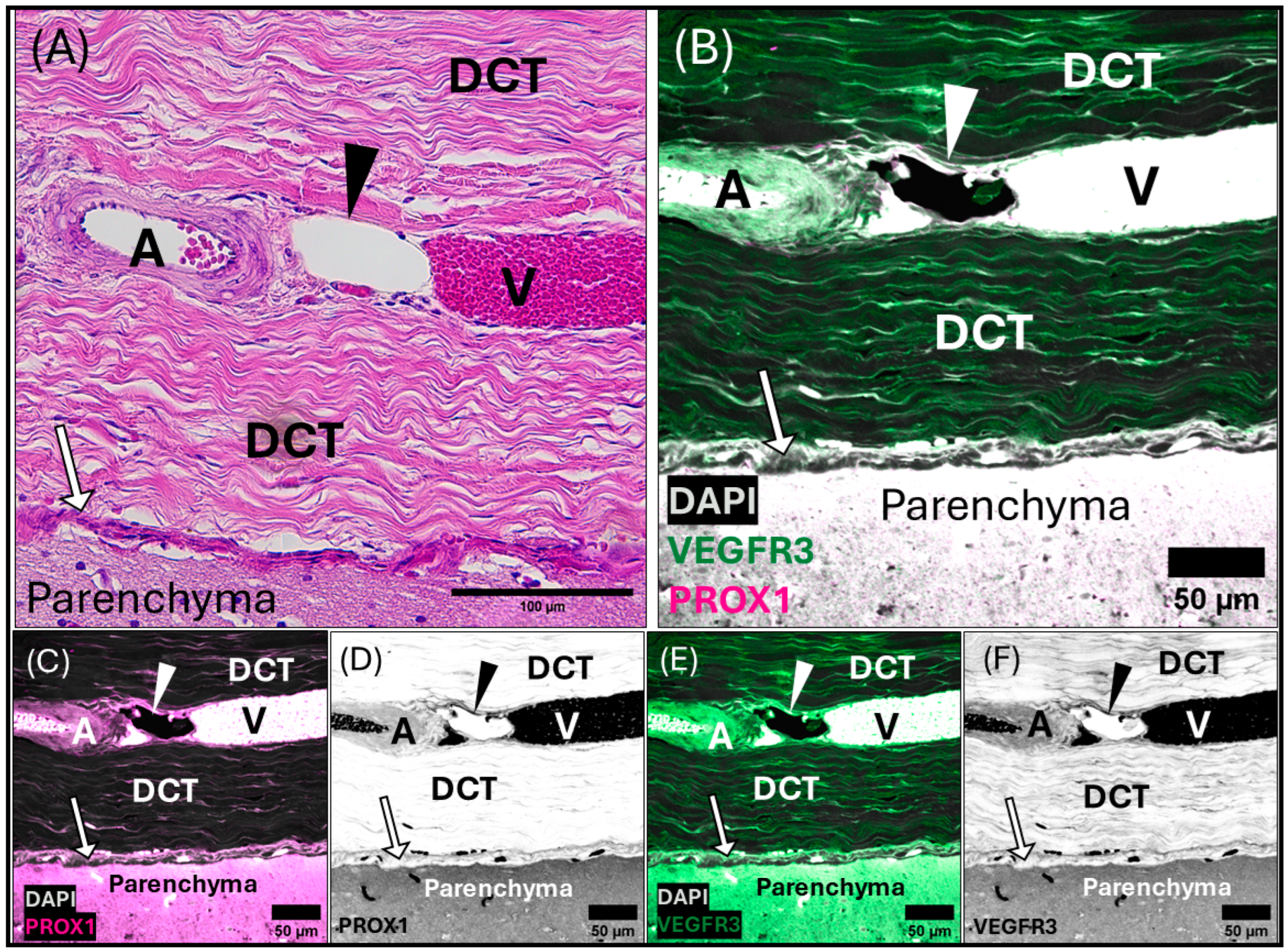
3.1. Dural Venous Sinuses
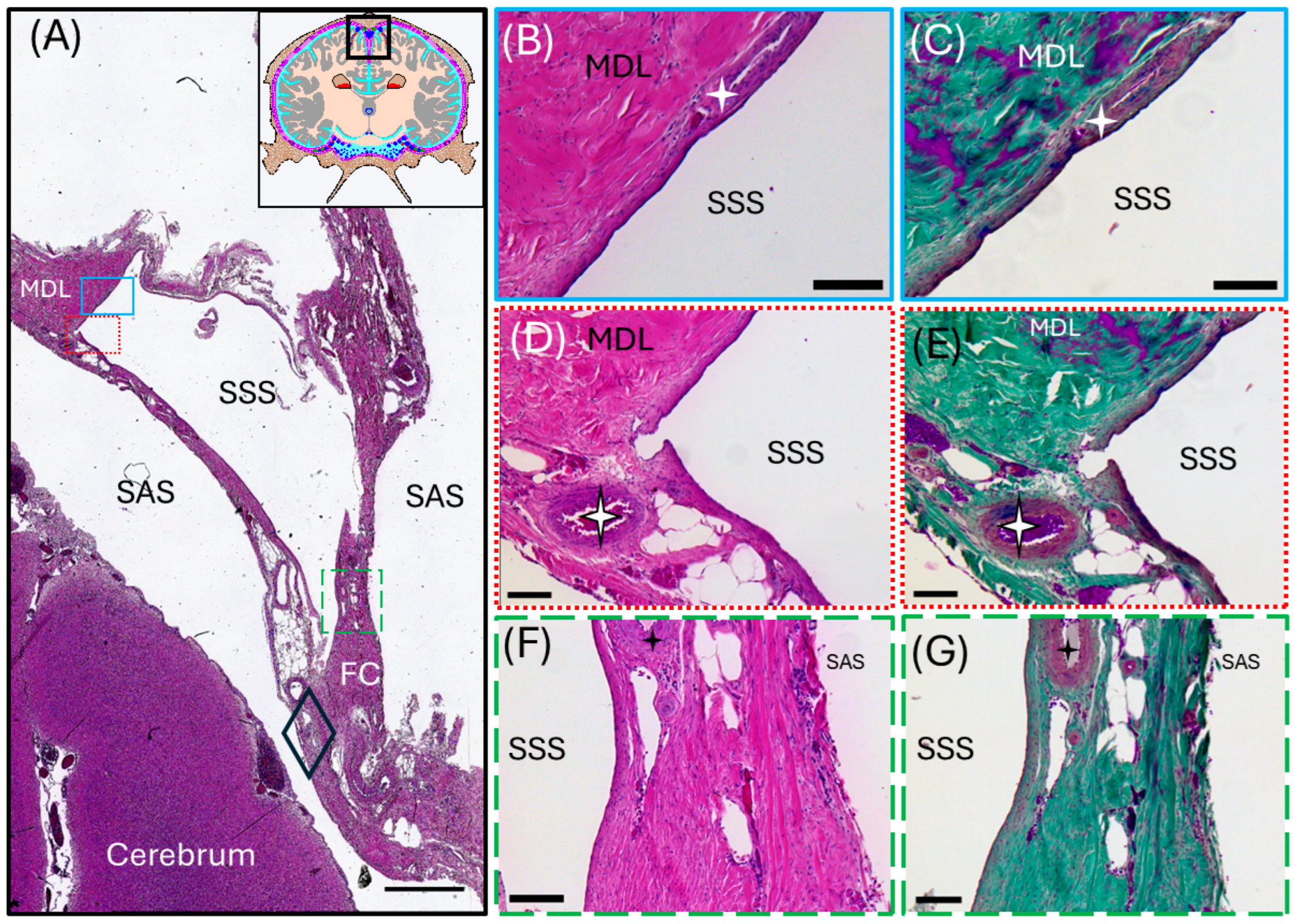
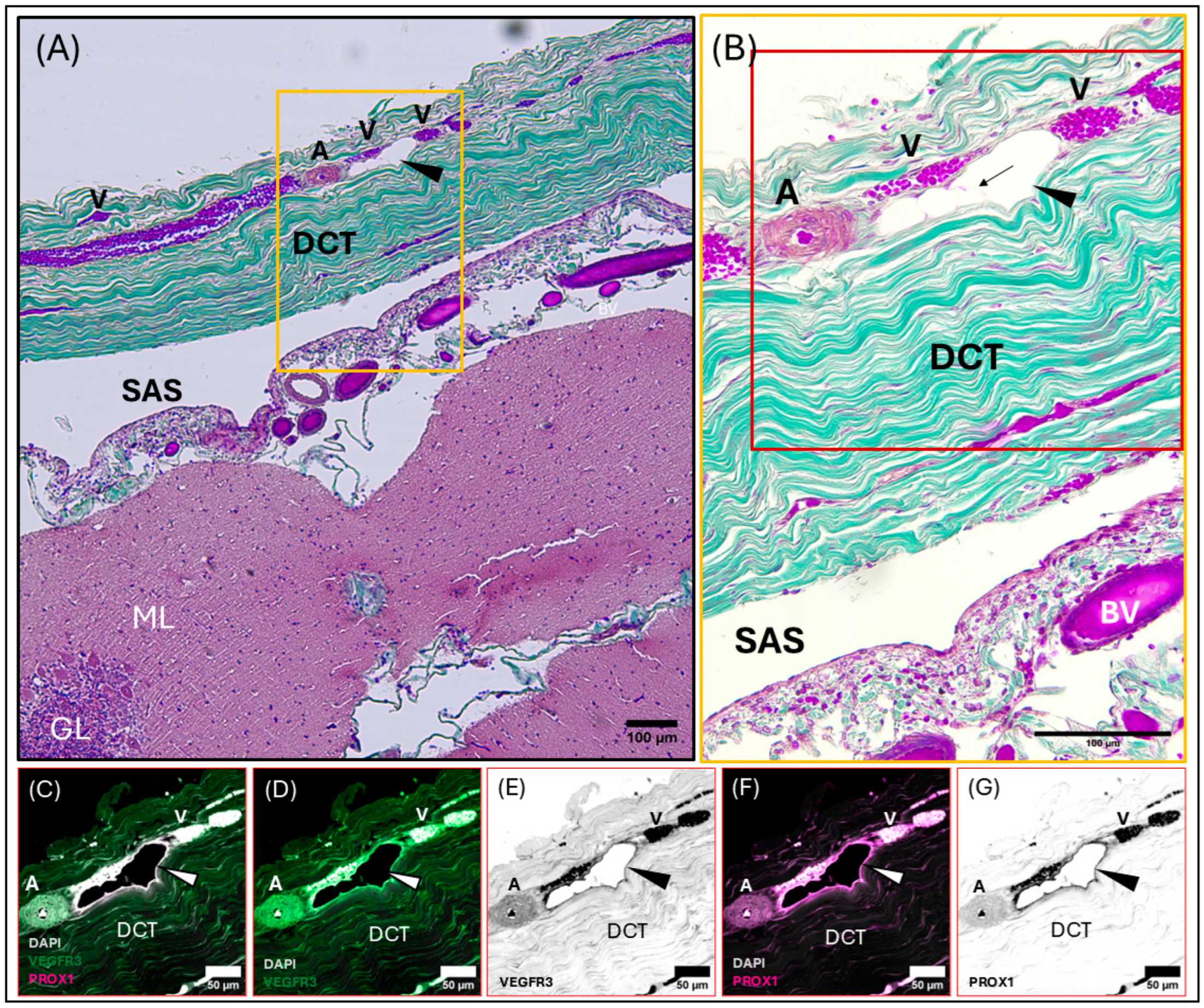
3.2. Meningeal Lymphatic Vessels
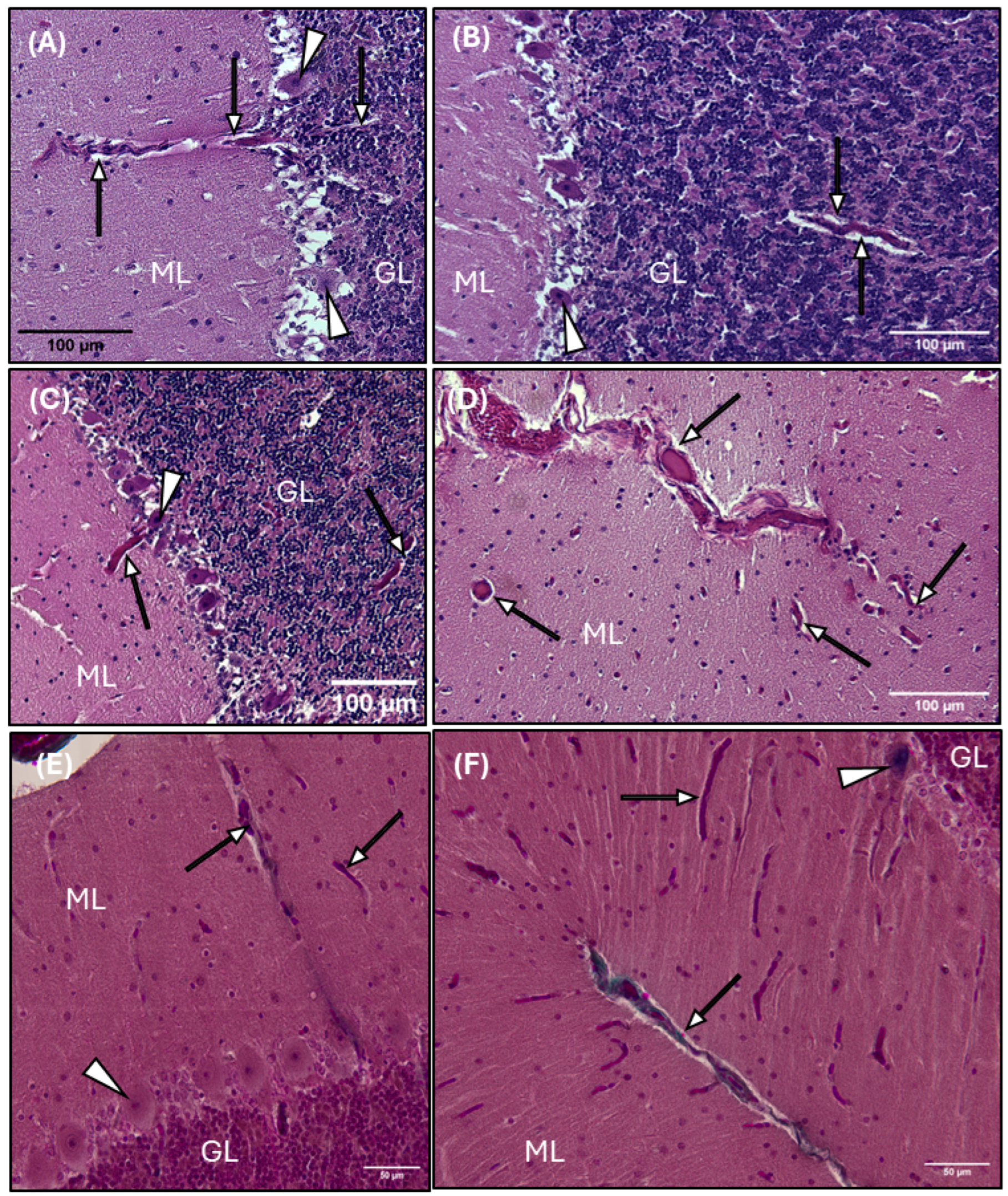
3.3. Perivascular Spaces and AQP4 Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. Fluid transport in the brain. Physiol. Rev. 2022, 102, 1025–1151. [Google Scholar] [CrossRef] [PubMed]
- Da Mesquita, S.; Fu, Z.; Kipnis, J. The meningeal lymphatic system: A new player in neurophysiology. Neuron 2018, 100, 375–388. [Google Scholar] [CrossRef]
- Mestre, H.; Hablitz, L.M.; Xavier, A.L.; Feng, W.; Zou, W.; Pu, T.; Monai, H.; Murlidharan, G.; Castellanos Rivera, R.M.; Simon, M.J. Aquaporin-4-dependent glymphatic solute transport in the rodent brain. eLife 2018, 7, e40070. [Google Scholar] [CrossRef] [PubMed]
- Gomolka, R.S.; Hablitz, L.M.; Mestre, H.; Giannetto, M.; Du, T.; Hauglund, N.L.; Xie, L.; Peng, W.; Martinez, P.M.; Nedergaard, M. Loss of aquaporin-4 results in glymphatic system dysfunction via brain-wide interstitial fluid stagnation. eLife 2023, 12, e82232. [Google Scholar] [CrossRef]
- Bayot, M.L.; Reddy, V.; Zabel, M.K. Neuroanatomy, dural venous sinuses. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Chen, W.W.; Zhang, X.; Huang, W.J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [PubMed]
- Jessen, N.A.; Munk, A.S.; Lundgaard, I.; Nedergaard, M. The Glymphatic System: A Beginner’s Guide. Neurochem. Res. 2015, 40, 2583–2599. [Google Scholar] [CrossRef]
- Plog, B.A.; Nedergaard, M. The Glymphatic System in Central Nervous System Health and Disease: Past, Present, and Future. Annu. Rev. Pathol. 2018, 13, 379–394. [Google Scholar] [CrossRef]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef]
- Jiang, H.; Wei, H.; Zhou, Y.; Xiao, X.; Zhou, C.; Ji, X. Overview of the meningeal lymphatic vessels in aging and central nervous system disorders. Cell Biosci. 2022, 12, 202. [Google Scholar] [CrossRef]
- Licastro, E.; Pignataro, G.; Iliff, J.J.; Xiang, Y.; Lo, E.H.; Hayakawa, K.; Esposito, E. Glymphatic and lymphatic communication with systemic responses during physiological and pathological conditions in the central nervous system. Commun. Biol. 2024, 7, 229. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.; Thrane, A.S.; Nedergaard, M. The Glymphatic System and Brain Interstitial Fluid Homeostasis; Academic Press: London, UK, 2017; pp. 17–25. [Google Scholar]
- Kiviniemi, V.; Wang, X.; Korhonen, V.; Keinänen, T.; Tuovinen, T.; Autio, J.; LeVan, P.; Keilholz, S.; Zang, Y.-F.; Hennig, J. Ultra-fast magnetic resonance encephalography of physiological brain activity–glymphatic pulsation mechanisms? J. Cereb. Blood Flow. Metab. 2016, 36, 1033–1045. [Google Scholar] [CrossRef]
- Mestre, H.; Tithof, J.; Du, T.; Song, W.; Peng, W.; Sweeney, A.M.; Olveda, G.; Thomas, J.H.; Nedergaard, M.; Kelley, D.H. Flow of cerebrospinal fluid is driven by arterial pulsations and is reduced in hypertension. Nat. Commun. 2018, 9, 4878. [Google Scholar] [CrossRef]
- van Veluw, S.J.; Hou, S.S.; Calvo-Rodriguez, M.; Arbel-Ornath, M.; Snyder, A.C.; Frosch, M.P.; Greenberg, S.M.; Bacskai, B.J. Vasomotion as a driving force for paravascular clearance in the awake mouse brain. Neuron 2020, 105, 549–561.e545. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Iliff, J.J. The impact of neurovascular, blood-brain barrier, and glymphatic dysfunction in neurodegenerative and metabolic diseases. Int. Rev. Neurobiol. 2020, 154, 413–436. [Google Scholar]
- Absinta, M.; Ha, S.K.; Nair, G.; Sati, P.; Luciano, N.J.; Palisoc, M.; Louveau, A.; Zaghloul, K.A.; Pittaluga, S.; Kipnis, J.; et al. Human and nonhuman primate meninges harbor lymphatic vessels that can be visualized noninvasively by MRI. eLife 2017, 6, e29738. [Google Scholar] [CrossRef]
- Bower, N.I.; Koltowska, K.; Pichol-Thievend, C.; Virshup, I.; Paterson, S.; Lagendijk, A.K.; Wang, W.; Lindsey, B.W.; Bent, S.J.; Baek, S.; et al. Mural lymphatic endothelial cells regulate meningeal angiogenesis in the zebrafish. Nat. Neurosci. 2017, 20, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.; Gardner, D.; Choi, D.; Park, E.; Jin Seong, Y.; Yang, S.; Castorena-Gonzalez, J.; Louveau, A.; Zhou, Z.; Lee, G.K.; et al. Development and Characterization of A Novel Prox1-EGFP Lymphatic and Schlemm’s Canal Reporter Rat. Sci. Rep. 2017, 7, 5577. [Google Scholar] [CrossRef]
- Keenan, T.F.; Jackson, O.N.; Nelson-Maney, N.P.; Rommel, S.A.; McLellan, W.A.; Pabst, D.A.; Costidis, A.; Caron, K.M.; Kernagis, D.N.; Rotstein, D.S.; et al. Morphology of the glymphatic and meningeal lymphatic structures of the bottlenose dolphin (Tursiops truncatus). Scientific reports, under review.
- Andrews, R.D.; Jones, D.R.; Williams, J.D.; Thorson, P.H.; Oliver, G.W.; Costa, D.P.; Le Boeuf, B.J. Heart rates of northern elephant seals diving at sea and resting on the beach. J. Exp. Biol. 1997, 200, 2083–2095. [Google Scholar] [CrossRef]
- Houser, D.S.; Dankiewicz-Talmadge, L.A.; Stockard, T.K.; Ponganis, P.J. Investigation of the potential for vascular bubble formation in a repetitively diving dolphin. J. Exp. Biol. 2010, 213, 52–62. [Google Scholar] [CrossRef]
- Ponganis, P.J. Diving Physiology of Marine Mammals and Seabirds, 1st ed.; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Scholander, P.F. Experimental Investigations on the Respiratory Function in Diving Mammals and Birds; I kommisjon hos Jacob Dybwad: Oslo, Norway, 1940; p. 131. [Google Scholar]
- Zapol, W.M.; Liggins, G.; Schneider, R.C.; Qvist, J.; Snider, M.T.; Creasy, R.K.; Hochachka, P.W. Regional blood flow during simulated diving in the conscious Weddell seal. J. Appl. Physiol. 1979, 47, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Irving, L.; Scholander, P.; Grinnell, S. The regulation of arterial blood pressure in the seal during diving. Am. J. Physiol. -Leg. Content 1942, 135, 557–566. [Google Scholar] [CrossRef]
- Dreha-Kulaczewski, S.; Joseph, A.A.; Merboldt, K.-D.; Ludwig, H.-C.; Gärtner, J.; Frahm, J. Inspiration is the major regulator of human CSF flow. J. Neurosci. 2015, 35, 2485–2491. [Google Scholar] [CrossRef]
- Dreha-Kulaczewski, S.; Joseph, A.A.; Merboldt, K.-D.; Ludwig, H.-C.; Gärtner, J.; Frahm, J. Identification of the upward movement of human CSF in vivo and its relation to the brain venous system. J. Neurosci. 2017, 37, 2395–2402. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, S.H.; Hanson, A.C. Sperm whales and killer whales with the largest brains of all toothed whales show extreme differences in cerebellum. Brain Behav. Evol. 2014, 83, 266–274. [Google Scholar] [CrossRef]
- Kesarev, V.; Malofeeva, L.; Trykova, O. Structural organization of the cetacean neocortex. Arkhiv Anat. Gistol. I Embriol. 1977, 73, 23–30. [Google Scholar]
- Glezer, I.; Jacobs, M.; Morgane, P. The so-called “initial” type of neocortex in relation to cetacean brain organization. In Proceedings of the Abstracts of the 15th Annual Meeting of the Society for Neuroscience, Dallas, TX, USA, 20–25 October 1985; p. 1308. [Google Scholar]
- Glezer, I.I.; Jacobs, M.S.; Morgane, P.J. Ultrastructure of the blood-brain barrier in the dolphin (Stenella coeruleoalba). Brain Res. 1987, 414, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, C.E.; McLellan, W.A.; Rommel, S.A.; Costidis, A.M.; Yopak, K.E.; Koopman, H.N.; Glandon, H.L.; Ann Pabst, D. Comparative morphology of the spinal cord and associated vasculature in shallow versus deep diving cetaceans. J. Morphol. 2021, 282, 1415–1431. [Google Scholar] [CrossRef]
- Costidis, A. The Morphology of the Venous System in the Head and Neck of the Bottlenose Dolphin (Tursiops truncatus) and the Florida Manatee (Trichechus manatus latirostris). Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2012. [Google Scholar]
- Viamonte, M.; Morgane, P.; Galliano, R.; Nagel, E.; McFarland, W. Angiography in the living dolphin and observations on blood supply to the brain. Am. J. Physiol.-Leg. Content 1968, 214, 1225–1249. [Google Scholar] [CrossRef]
- McFarland, W. Blood supply to the brain of the dolphin, Tursiops truncatus, with comparative observations on special aspects of the cerebrovascular supply of other vertebrates. Neurosci. Behav. Rev. 1979, 3, 1–93. [Google Scholar]
- Kooyman, G.L.; Ponganis, P. The physiological basis of diving to depth: Birds and mammals. Annu. Rev. Physiol. 1998, 60, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Nagel, E.; Morgane, P.; McFarland, W.; Galllano, R. Rete mirabile of dolphin: Its pressure-damping effect on cerebral circulation. Science 1968, 161, 898–900. [Google Scholar] [CrossRef] [PubMed]
- Vogl, A.; Fisher, H. The internal carotid artery does not directly supply the brain in the Monodontidae (Order Cetacea). J. Morphol. 1981, 170, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Berta, A.; Ekdale, E.G.; Cranford, T.W. Review of the cetacean nose: Form, function, and evolution. Anat. Rec. 2014, 297, 2205–2215. [Google Scholar] [CrossRef]
- Miller, G.S. The Telescoping of the Cetacean Skull (with Eight Plates); Smithsonian Institution: Washington, DC, USA, 1923. [Google Scholar]
- Ridgway, S.; Carder, D.; Green, R.; Gaunt, A.; Gaunt, S.; Evans, W. Electromyographic and pressure events in the nasolaryngeal system of dolphins during sound production. In Animal Sonar Systems; Springer: Berlin/Heidelberg, Germany, 1980; pp. 239–249. [Google Scholar]
- Ridgway, S.H.; Carder, D.A. Nasal pressure and sound production in an echolocating white whale, Delphinapterus leucas. In Animal sonar: Processes and Performance; Springer: Berlin/Heidelberg, Germany, 1988; pp. 53–60. [Google Scholar]
- Roston, R.A.; Boessenecker, R.W.; Geisler, J.H. Evolution and development of the cetacean skull roof: A case study in novelty and homology. Philos. Trans. R. Soc. B 2023, 378, 20220086. [Google Scholar] [CrossRef]
- Roston, R.A.; Roth, V.L. Cetacean skull telescoping brings evolution of cranial sutures into focus. Anat. Rec. 2019, 302, 1055–1073. [Google Scholar] [CrossRef]
- Cozzi, B.; Huggenberger, S.; Oelschläger, H.A. Anatomy of Dolphins: Insights into Body Structure and Function; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Lillie, M.; Vogl, A.; Gerard, S.; Raverty, S.; Shadwick, R. Retia mirabilia: Protecting the cetacean brain from locomotion-generated blood pressure pulses. Science 2022, 377, 1452–1456. [Google Scholar] [CrossRef]
- Ridgway, S.H.; Brownson, R. Relative brain sizes and cortical surface areas in odontocetes. Acta Zool. Fenn. 1984, 172, 149–152. [Google Scholar]
- Gerussi, T.; Graïc, J.-M.; Peruffo, A.; Behroozi, M.; Schlaffke, L.; Huggenberger, S.; Güntürkün, O.; Cozzi, B. The prefrontal cortex of the bottlenose dolphin (Tursiops truncatus Montagu, 1821): A tractography study and comparison with the human. Brain Struct. Funct. 2023, 228, 1963–1976. [Google Scholar] [CrossRef]
- Di Guardo, G. Cetaceans, models for human disease? Res. Vet. Sci. 2018, 119, 43–44. [Google Scholar] [CrossRef]
- Di Guardo, G. Alzheimer’s disease, cellular prion protein, and dolphins. Alzheimer’s Dement. 2018, 14, 259–260. [Google Scholar] [CrossRef]
- Zimmer, W.M.X.; Tyack, P.L. Repetitive shallow dives pose decompression risk in deep-diving beaked whales. Mar. Mammal. Sci. 2007, 23, 888–925. [Google Scholar] [CrossRef]
- Fernández, A.; Sierra, E.; Díaz-Delgado, J.; Sacchini, S.; Sánchez-Paz, Y.; Suárez-Santana, C.; Arregui, M.; Arbelo, M.; Bernaldo de Quirós, Y. Deadly acute Decompression Sickness in Risso’s dolphins. Sci. Rep. 2017, 7, 13621. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, A.W. Frequency of decompression illness among recent and extinct mammals and “reptiles”: A review. Sci. Nat. 2017, 104, 56. [Google Scholar] [CrossRef] [PubMed]
- Fahlman, A.; Tyack, P.; Miller, P.; Kvadsheim, P. How man-made interference might cause gas bubble emboli in deep diving whales. Front. Physiol. 2014, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Delgado, J.; Fernández, A.; Sierra, E.; Sacchini, S.; Andrada, M.; Vela, A.I.; Quesada-Canales, Ó.; Paz, Y.; Zucca, D.; Groch, K. Pathologic findings and causes of death of stranded cetaceans in the Canary Islands (2006–2012). PLoS ONE 2018, 13, e0204444. [Google Scholar] [CrossRef]
- Sacchini, S.; Díaz-Delgado, J.; Espinosa de los Monteros, A.; Paz, Y.; Bernaldo de Quirós, Y.; Sierra, E.; Arbelo, M.; Herráez, P.; Fernández, A. Amyloid-beta peptide and phosphorylated tau in the frontopolar cerebral cortex and in the cerebellum of toothed whales: Aging versus hypoxia. Biol. Open 2020, 9, bio054734. [Google Scholar] [CrossRef]
- Ketten, D.; Rowles, T.; Cramer, S.; O’Malley, J.; Arruda, J.; Evans, P. Cranial trauma in beaked whales. In Proceedings of the Workshop on Active Sonar and Cetaceans Held at the European Cetacean Society 17th Annual Conference, Las Palmas de Gran Canaria, Spain, 10–13 March 2004; pp. 21–27. [Google Scholar]
- Jepson, P.D.; Arbelo, M.; Deaville, R.; Patterson, I.A.; Castro, P.; Baker, J.R.; Degollada, E.; Ross, H.M.; Herráez, P.; Pocknell, A.M.; et al. Gas-bubble lesions in stranded cetaceans. Nature 2003, 425, 575–576. [Google Scholar] [CrossRef]
- Fernández, A.; Edwards, J.; Rodriguez, F.; De Los Monteros, A.E.; Herraez, P.; Castro, P.; Jaber, J.; Martin, V.; Arbelo, M. “Gas and fat embolic syndrome” involving a mass stranding of beaked whales (family Ziphiidae) exposed to anthropogenic sonar signals. Vet. Pathol. 2005, 42, 446–457. [Google Scholar] [CrossRef]
- Davis, D.A.; Mondo, K.; Stern, E.; Annor, A.K.; Murch, S.J.; Coyne, T.M.; Brand, L.E.; Niemeyer, M.E.; Sharp, S.; Bradley, W.G. Cyanobacterial neurotoxin BMAA and brain pathology in stranded dolphins. PLoS ONE 2019, 14, e0213346. [Google Scholar] [CrossRef]
- Stylianaki, I.; Komnenou, A.T.; Posantzis, D.; Nikolaou, K.; Papaioannou, N. Alzheimer’s disease-like pathological lesions in an aged bottlenose dolphin (Tursiops truncatus). Vet. Rec. Case Rep. 2019, 7, e000700. [Google Scholar] [CrossRef]
- Byrd, B.L.; Hohn, A.A.; Lovewell, G.N.; Altman, K.M.; Barco, S.G.; Friedlaender, A.; Harms, C.A.; McLellan, W.A.; Moore, K.T.; Rosel, P.E. Strandings as indicators of marine mammal biodiversity and human interactions off the coast of North Carolina. Fish. Bull. 2014, 112, 1–23. [Google Scholar] [CrossRef]
- Geraci, J.; Lounsbury, V. Marine Mammals Ashore: A Field Guide for Strandings; National Aquarium: Baltimore, MD, USA, 2005. [Google Scholar]
- Raverty, S.; Duignan, P.J.; Jepson, P.D.; Morell, M. Marine Mammal Gross Necropsy. In Marine Mammal Medicine, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2018; p. 18. [Google Scholar]
- Coles, J.A.; Myburgh, E.; Brewer, J.M.; McMenamin, P.G. Where are we? The anatomy of the murine cortical meninges revisited for intravital imaging, immunology, and clearance of waste from the brain. Prog. Neurobiol. 2017, 156, 107–148. [Google Scholar] [CrossRef] [PubMed]
- Schain, A.J.; Melo-Carrillo, A.; Borsook, D.; Grutzendler, J.; Strassman, A.M.; Burstein, R. Activation of pial and dural macrophages and dendritic cells by cortical spreading depression. Ann. Neurol. 2018, 83, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.F.; McLellan, W.A.; Rommel, S.A.; Costidis, A.M.; Harms, C.A.; Thewissen, H.J.; Rotstein, D.S.; Gay, M.D.; Potter, C.W.; Taylor, A.R. Gross and histological morphology of the cervical gill slit gland of the pygmy sperm whale (Kogia breviceps). Anat. Rec. 2022, 305, 688–703. [Google Scholar] [CrossRef]
- Lillie, R.D. Histopathologic Technic and Practical Histochemistry; The Blakiston: Philadelphia, PA, USA, 1954. [Google Scholar]
- Pawlak, J.B.; Caron, K.M. Lymphatic programing and specialization in hybrid vessels. Front. Physiol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Beck, S.; Gunawardena, P.; Horton, D.; Hicks, D.; Marston, D.; Ortiz-Pelaez, A.; Fooks, A.; Núñez, A. Pathobiological investigation of naturally infected canine rabies cases from Sri Lanka. BMC Vet. Res. 2017, 13, 99. [Google Scholar] [CrossRef]
- Chang, A.-M.; Chen, C.-C.; Chang, C.-D.; Huang, Y.-L.; Ke, G.-M.; Walther, B.A. Encephalitis induced by a newly discovered ruminant rhadinovirus in a free-living Formosan sambar deer (Rusa unicolor swinhoei). J. Vet. Med. Sci. 2018, 80, 810–813. [Google Scholar] [CrossRef]
- Gareau, P.J.; Wymore, A.C.; Cofer, G.P.; Johnson, G.A. Imaging inflammation: Direct visualization of perivascular cuffing in EAE by magnetic resonance microscopy. J. Magn. Reson. Imaging Off. J. Int. Soc. Magn. Reson. Med. 2002, 16, 28–36. [Google Scholar] [CrossRef]
- Braun, M.; Sevao, M.; Keil, S.A.; Gino, E.; Wang, M.X.; Lee, J.; Haveliwala, M.A.; Klein, E.; Agarwal, S.; Pedersen, T. Macroscopic changes in aquaporin-4 underlie blast traumatic brain injury-related impairment in glymphatic function. Brain 2024, 147, 2214–2229. [Google Scholar] [CrossRef] [PubMed]
- Zeppenfeld, D.M.; Simon, M.; Haswell, J.D.; D’Abreo, D.; Murchison, C.; Quinn, J.F.; Grafe, M.R.; Woltjer, R.L.; Kaye, J.; Iliff, J.J. Association of perivascular localization of aquaporin-4 with cognition and Alzheimer disease in aging brains. JAMA Neurol. 2017, 74, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Vacher, M.C.; Durrant, C.S.; Rose, J.; Hall, A.J.; Spires-Jones, T.L.; Gunn-Moore, F.; Dagleish, M.P. Alzheimer’s disease-like neuropathology in three species of oceanic dolphin. Eur. J. Neurosci. 2023, 57, 1161–1179. [Google Scholar] [CrossRef]
- Letchuman, V.; Donohoe, C. Neuroanatomy, Superior Sagittal Sinus; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Sindou, M.; Auque, J. The intracranial venous system as a neurosurgeon’s perspective. Adv. Tech. Stand. Neurosurg. 2000, 26, 131–216. [Google Scholar] [PubMed]
- Walsh, D.R.; Lynch, J.J.; O’Connor, D.T.; Newport, D.T.; Mulvihill, J.J. Mechanical and structural characterisation of the dural venous sinuses. Sci. Rep. 2020, 10, 21763. [Google Scholar] [CrossRef]
- Ahn, J.H.; Cho, H.; Kim, J.-H.; Kim, S.H.; Ham, J.-S.; Park, I.; Suh, S.H.; Hong, S.P.; Song, J.-H.; Hong, Y.-K. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature 2019, 572, 62–66. [Google Scholar] [CrossRef]
- Aspelund, A.; Antila, S.; Proulx, S.T.; Karlsen, T.V.; Karaman, S.; Detmar, M.; Wiig, H.; Alitalo, K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 2015, 212, 991–999. [Google Scholar] [CrossRef]
- Izen, R.M.; Yamazaki, T.; Nishinaka-Arai, Y.; Hong, Y.K.; Mukouyama, Y.S. Postnatal development of lymphatic vasculature in the brain meninges. Dev. Dyn. 2018, 247, 741–753. [Google Scholar] [CrossRef]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S. Structural and functional features of central nervous system lymphatic vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef]
- Louveau, A.; Herz, J.; Alme, M.N.; Salvador, A.F.; Dong, M.Q.; Viar, K.E.; Herod, S.G.; Knopp, J.; Setliff, J.C.; Lupi, A.L. CNS lymphatic drainage and neuroinflammation are regulated by meningeal lymphatic vasculature. Nat. Neurosci. 2018, 21, 1380–1391. [Google Scholar] [CrossRef]
- Iliff, J.J.; Wang, M.; Zeppenfeld, D.M.; Venkataraman, A.; Plog, B.A.; Liao, Y.; Deane, R.; Nedergaard, M. Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain. J. Neurosci. 2013, 33, 18190–18199. [Google Scholar] [CrossRef] [PubMed]
- Louveau, A.; Plog, B.A.; Antila, S.; Alitalo, K.; Nedergaard, M.; Kipnis, J. Understanding the functions and relationships of the glymphatic system and meningeal lymphatics. J. Clin. Investig. 2017, 127, 3210–3219. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Delgado, J.; Sacchini, S.; Suárez-Bonnet, A.; Sierra, E.; Arbelo, M.; Espinosa, A.; Bassas, E.R.-G.; Mompeo, B.; Pérez, L.; Fernández, A. High-grade astrocytoma (glioblastoma multiforme) in an Atlantic spotted dolphin (Stenella frontalis). J. Comp. Pathol. 2015, 152, 278–282. [Google Scholar] [CrossRef]
- Ackermans, N.L.; Varghese, M.; Wicinski, B.; Torres, J.; De Gasperi, R.; Pryor, D.; Elder, G.A.; Gama Sosa, M.A.; Reidenberg, J.S.; Williams, T.M. Unconventional animal models for traumatic brain injury and chronic traumatic encephalopathy. J. Neurosci. Res. 2021, 99, 2463–2477. [Google Scholar] [CrossRef]
- Ridgway, S.H. Physiological observations on dolphin brains. In Dolphin Cognition and Behavior; Psychology Press: East Sussex, UK, 2013; pp. 31–59. [Google Scholar]
- Liu, Y.; Li, K.; Huang, Y.; Sun, J.; Gao, X. Treatment of the superior sagittal sinus and transverse sinus thrombosis associated with intracranial hemorrhage with the mechanical thrombectomy and thrombolytics: Case report. Medicine 2017, 96, e9038. [Google Scholar] [CrossRef]
- Patchana, T.; Zampella, B.; Berry, J.A.; Lawandy, S.; Sweiss, R.B.; Sweiss, R. Superior sagittal sinus: A review of the history, surgical considerations, and pathology. Cureus 2019, 11, e4597. [Google Scholar] [CrossRef] [PubMed]
- Siegert, C.; Smelt, A.; De Bruin, T. Superior sagittal sinus thrombosis and thyrotoxicosis: Possible association in two cases. Stroke 1995, 26, 496–497. [Google Scholar] [CrossRef]
- Visanji, N.P.; Lang, A.E.; Munoz, D.G. Lymphatic vasculature in human dural superior sagittal sinus: Implications for neurodegenerative proteinopathies. Neurosci. Lett. 2018, 665, 18–21. [Google Scholar] [CrossRef]
- Schaller, B.; Graf, R.; Sanada, Y.; Tolnay, M.; Rosner, G.; Wienhard, K.; Heiss, W.-D. Hemodynamic changes after occlusion of the posterior superior sagittal sinus: An experimental PET study in cats. Am. J. Neuroradiol. 2003, 24, 1876–1880. [Google Scholar]
- Gotoh, M.; Fukuhara, T.; Shirakawa, T.; Nishino, S.; Saijyo, T.; Kawauchi, M.; Ohmoto, T.; Kuyama, H. Experimental Study in Cerebral Venous Circulatory Disturbance: With Special Reference to Venous Hemorrhage. Recent. Adv. Neurotraumatol. 1993, 275–278. [Google Scholar]
- Guan, S.; Brookens, T. An overview of research efforts to understand the effects of underwater sound on cetaceans. Water Biol. Secur. 2023, 2, 100141. [Google Scholar] [CrossRef]
- Jepson, P.; Deaville, R.; Patterson, I.; Pocknell, A.; Ross, H.; Baker, J.; Howie, F.; Reid, R.; Colloff, A.; Cunningham, A. Acute and chronic gas bubble lesions in cetaceans stranded in the United Kingdom. Vet. Pathol. 2005, 42, 291–305. [Google Scholar] [CrossRef]
- Jepson, P.D.; Deaville, R.; Acevedo-Whitehouse, K.; Barnett, J.; Brownlow, A.; Brownell Jr, R.L.; Clare, F.C.; Davison, N.; Law, R.J.; Loveridge, J. What caused the UK’s largest common dolphin (Delphinus delphis) mass stranding event? PLoS ONE 2013, 8, e60953. [Google Scholar] [CrossRef]
- Li, G.; Cao, Y.; Tang, X.; Huang, J.; Cai, L.; Zhou, L. The meningeal lymphatic vessels and the glymphatic system: Potential therapeutic targets in neurological disorders. J. Cereb. Blood Flow. Metab. 2022, 42, 1364–1382. [Google Scholar] [CrossRef] [PubMed]
- Hershenhouse, K.S.; Shauly, O.; Gould, D.J.; Patel, K.M. Meningeal lymphatics: A review and future directions from a clinical perspective. Neurosci. Insights 2019, 14, 1179069519889027. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Fernández, A.; Felipe-Jiménez, I.; Zucca, D.; Di Francesco, G.; Díaz-Delgado, J.; Sacchini, S.; Rivero, M.A.; Arbelo, M. Neurobrucellosis in a common bottlenose dolphin (Tursiops truncatus) stranded in the Canary Islands. BMC Vet. Res. 2019, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Fernández, A.; Felipe-Jiménez, I.; Zucca, D.; Díaz-Delgado, J.; Puig-Lozano, R.; Câmara, N.; Consoli, F.; Díaz-Santana, P.; Suárez-Santana, C. Histopathological differential diagnosis of meningoencephalitis in cetaceans: Morbillivirus, Herpesvirus, Toxoplasma gondii, Brucella sp., and Nasitrema sp. Front. Vet. Sci. 2020, 7, 650. [Google Scholar] [CrossRef]
- Sierra, E.; Fernández, A.; Fernández-Maldonado, C.; Sacchini, S.; Felipe-Jiménez, I.; Segura-Göthlin, S.; Colom-Rivero, A.; Câmara, N.; Puig-Lozano, R.; Rambaldi, A.M. Molecular characterization of herpesviral encephalitis in cetaceans: Correlation with histopathological and immunohistochemical findings. Animals 2022, 12, 1149. [Google Scholar] [CrossRef]
- Wells, R.S.; Rhinehart, H.L.; Hansen, L.J.; Sweeney, J.C.; Townsend, F.I.; Stone, R.; Casper, D.R.; Scott, M.D.; Hohn, A.A.; Rowles, T.K. Bottlenose dolphins as marine ecosystem sentinels: Developing a health monitoring system. EcoHealth 2004, 1, 246–254. [Google Scholar] [CrossRef]
- Grattarola, C.; Minoia, L.; Giorda, F.; Consales, G.; Capanni, F.; Ceciarini, I.; Franchi, E.; Ascheri, D.; Garibaldi, F.; Dondo, A. Health Status of Stranded Common Bottlenose Dolphins (Tursiops truncatus) and Contamination by Immunotoxic Pollutants: A Threat to the Pelagos Sanctuary—Western Mediterranean Sea. Diversity 2023, 15, 569. [Google Scholar] [CrossRef]
- López-Berenguer, G.; Peñalver, J.; Martínez-López, E. A critical review about neurotoxic effects in marine mammals of mercury and other trace elements. Chemosphere 2020, 246, 125688. [Google Scholar] [CrossRef] [PubMed]
- Starnes, H.M.; Rock, K.D.; Jackson, T.W.; Belcher, S.M. A critical review and meta-analysis of impacts of per-and polyfluorinated substances on the brain and behavior. Front. Toxicol. 2022, 4, 881584. [Google Scholar] [CrossRef] [PubMed]
- Christensen, G.M.; Li, Z.; Liang, D.; Ebelt, S.; Gearing, M.; Levey, A.I.; Lah, J.J.; Wingo, A.; Wingo, T.; Hüls, A. Association of PM2. 5 Exposure and Alzheimer Disease Pathology in Brain Bank Donors—Effect Modification by APOE Genotype. Neurology 2024, 102, e209162. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal ID | Species | Strand Date | SI Code | Sex | Life History Category | Total Length (cm) |
|---|---|---|---|---|---|---|
| IFAW-12-364 * | Delphinus delphis | 25-Nov-2012 | 1 | Female | Subadult | 210 |
| CAHA559 | Delphinus delphis | 15-Feb-2022 | 1 | Male | Subadult | 191 |
| CAHA560 | Delphinus delphis | 15-Feb-2022 | 1 | Male | Subadult | 179 |
| JPIER040 | Delphinus delphis | 26-Feb-2023 | 2 | Male | Subadult | 202.5 |
| Antigen | Antibody Species/ Isotype | Target Species | Class | Suppliers | Concentration |
|---|---|---|---|---|---|
| VEGFR3 | Goat IgG | Human | Polyclonal | R&D Inc., Minneapolis, MN, USA (PN:AF349) | 0.2 µg/mL |
| Prox-1 | Rabbit IgG | Human, mouse, rat, zebrafish | Polyclonal | AngioBio Inc., San Diego, CA, USA (PN:11-002P) | 10 µg/mL |
| Secondary Antibody | |||||
| Cy-5-conjugated donkey anti-goat IgG | Jackson Immunoresearch (705-175-147) | 2.5 µg/mL | |||
| Cy-3-conjugated donkey anti-rabbit IgG | Jackson Immunoresearch (711-165-152) | 2.5 µg/mL | |||
| Protein | Percentage of Shared Identity to Human Protein |
|---|---|
| LYVE-1 | 74.84% |
| PROX-1 | 99.32% |
| VEGFR3 | 88.08% |
| Podoplanin | 50.20% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson, O.N.; Keenan, T.F.; Nelson-Maney, N.P.; Rommel, S.A.; McLellan, W.A.; Pabst, D.A.; Costidis, A.M.; Caron, K.M.; Kernagis, D.N.; Rotstein, D.S.; et al. Meningeal Lymphatic and Glymphatic Structures in a Pelagic Delphinid (Delphinus delphis). Animals 2025, 15, 729. https://doi.org/10.3390/ani15050729
Jackson ON, Keenan TF, Nelson-Maney NP, Rommel SA, McLellan WA, Pabst DA, Costidis AM, Caron KM, Kernagis DN, Rotstein DS, et al. Meningeal Lymphatic and Glymphatic Structures in a Pelagic Delphinid (Delphinus delphis). Animals. 2025; 15(5):729. https://doi.org/10.3390/ani15050729
Chicago/Turabian StyleJackson, Olivia N., Tiffany F. Keenan, Nathan P. Nelson-Maney, Sentiel A. Rommel, William A. McLellan, D. Ann Pabst, Alexander M. Costidis, Kathleen M. Caron, Dawn N. Kernagis, David S. Rotstein, and et al. 2025. "Meningeal Lymphatic and Glymphatic Structures in a Pelagic Delphinid (Delphinus delphis)" Animals 15, no. 5: 729. https://doi.org/10.3390/ani15050729
APA StyleJackson, O. N., Keenan, T. F., Nelson-Maney, N. P., Rommel, S. A., McLellan, W. A., Pabst, D. A., Costidis, A. M., Caron, K. M., Kernagis, D. N., Rotstein, D. S., Thayer, V. G., Harms, C. A., Piscitelli-Doshkov, M. A., Doshkov, P., Schweikert, L. E., Yopak, K. E., Braun, M., & Tift, M. S. (2025). Meningeal Lymphatic and Glymphatic Structures in a Pelagic Delphinid (Delphinus delphis). Animals, 15(5), 729. https://doi.org/10.3390/ani15050729