
Dispersal Limitation Dominates Riverine Fish Communities in the Areas of the Water Diversion Project in the Western Sichuan Plateau, China



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
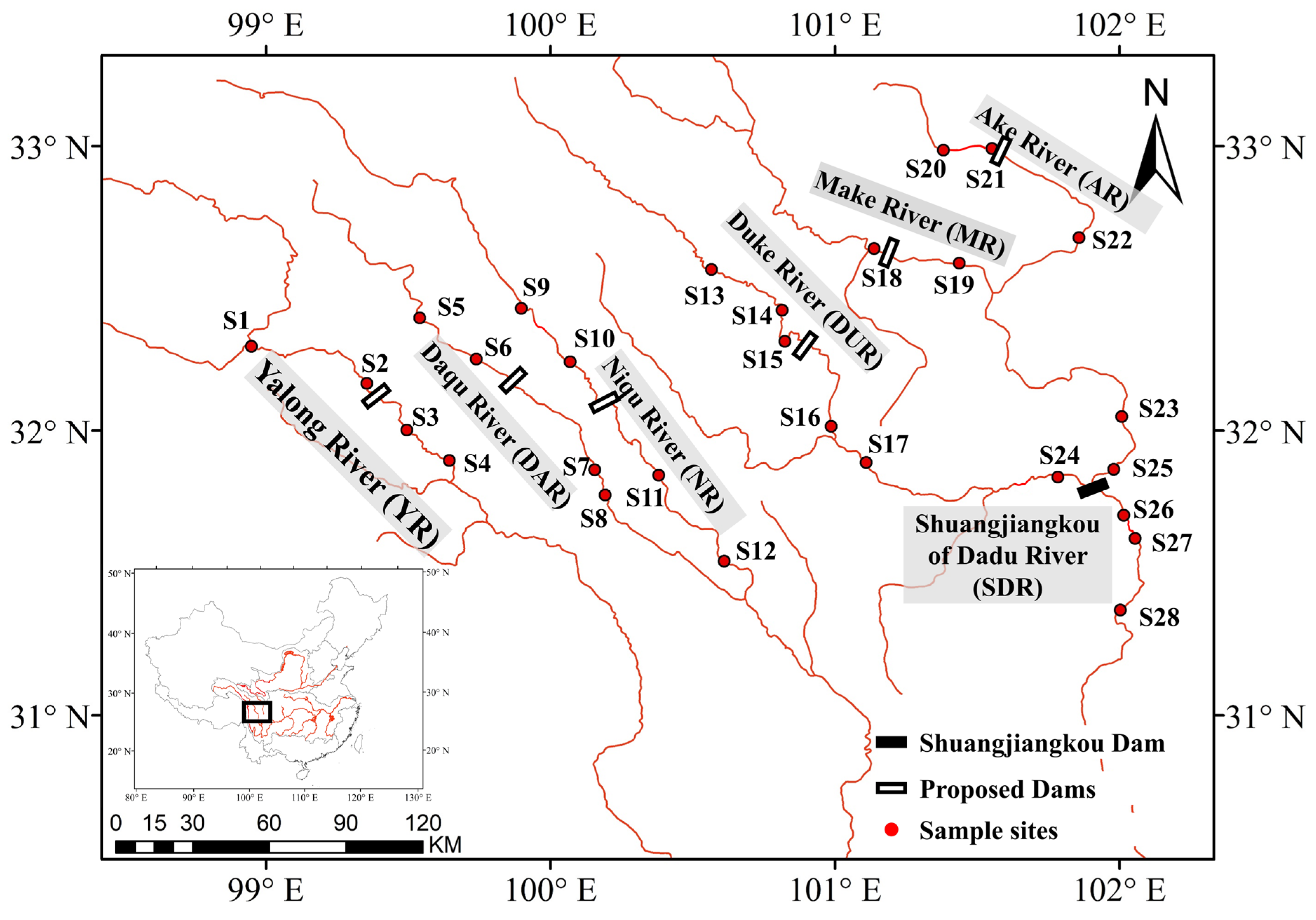
2.1. Study Area
2.2. Fish Sampling
2.3. Physicochemical Factors
2.4. Data Analysis
3. Results
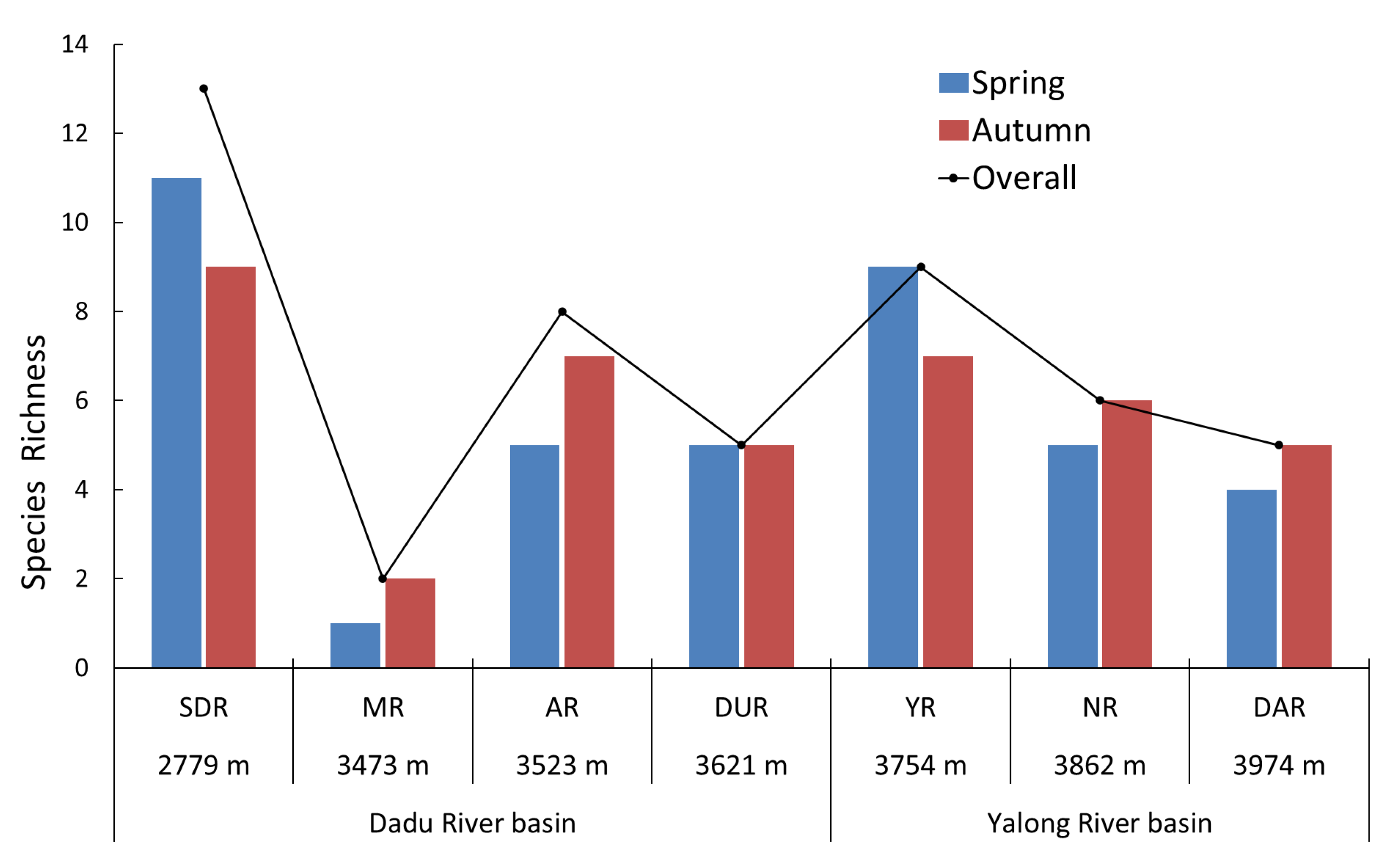
3.1. Environmental Characteristics
3.2. Species Composition and Dominant Species
3.3. Spatial–Temporal Changes in Fish Communities
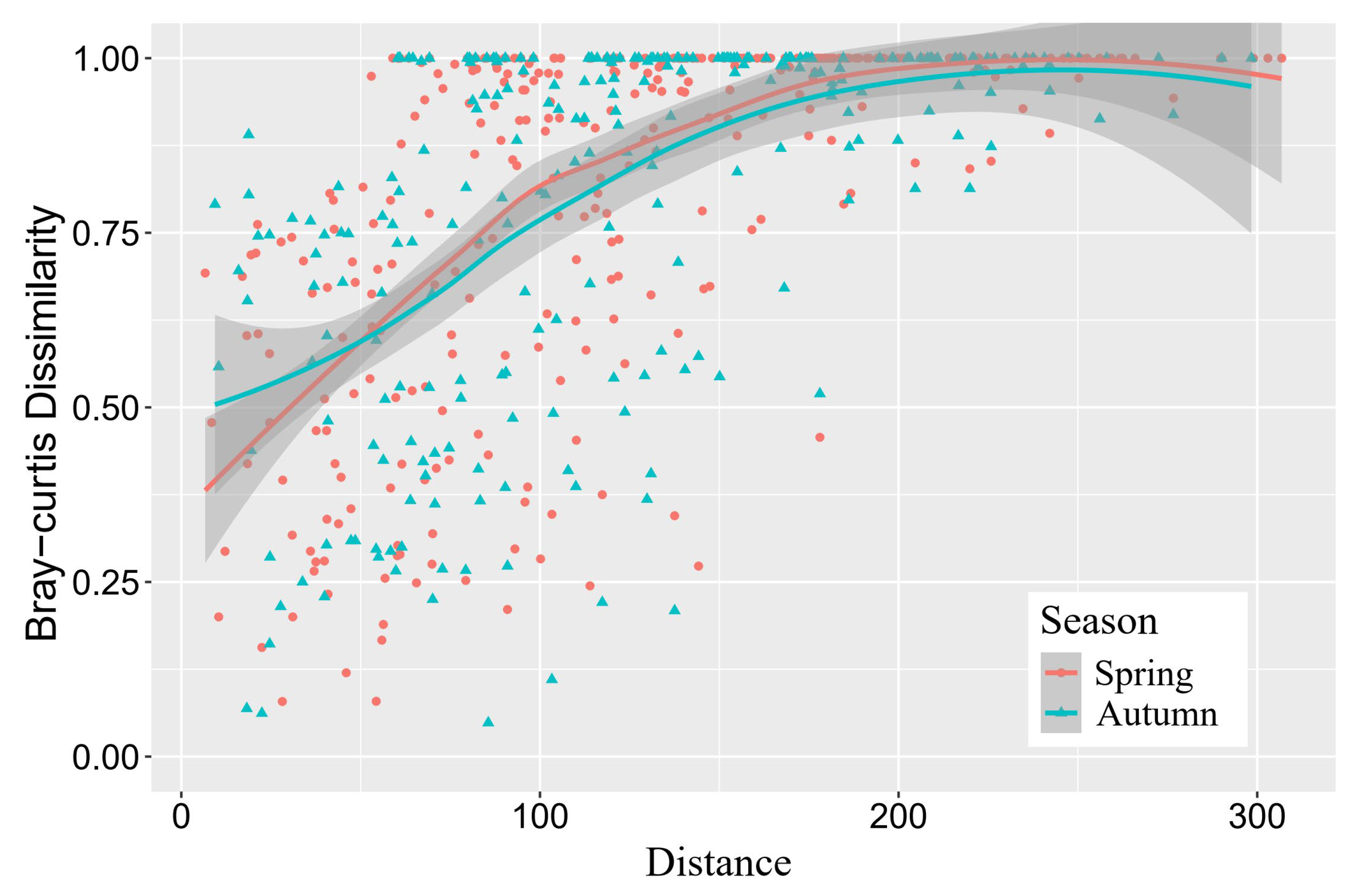
3.4. Influence of Geographical Distance
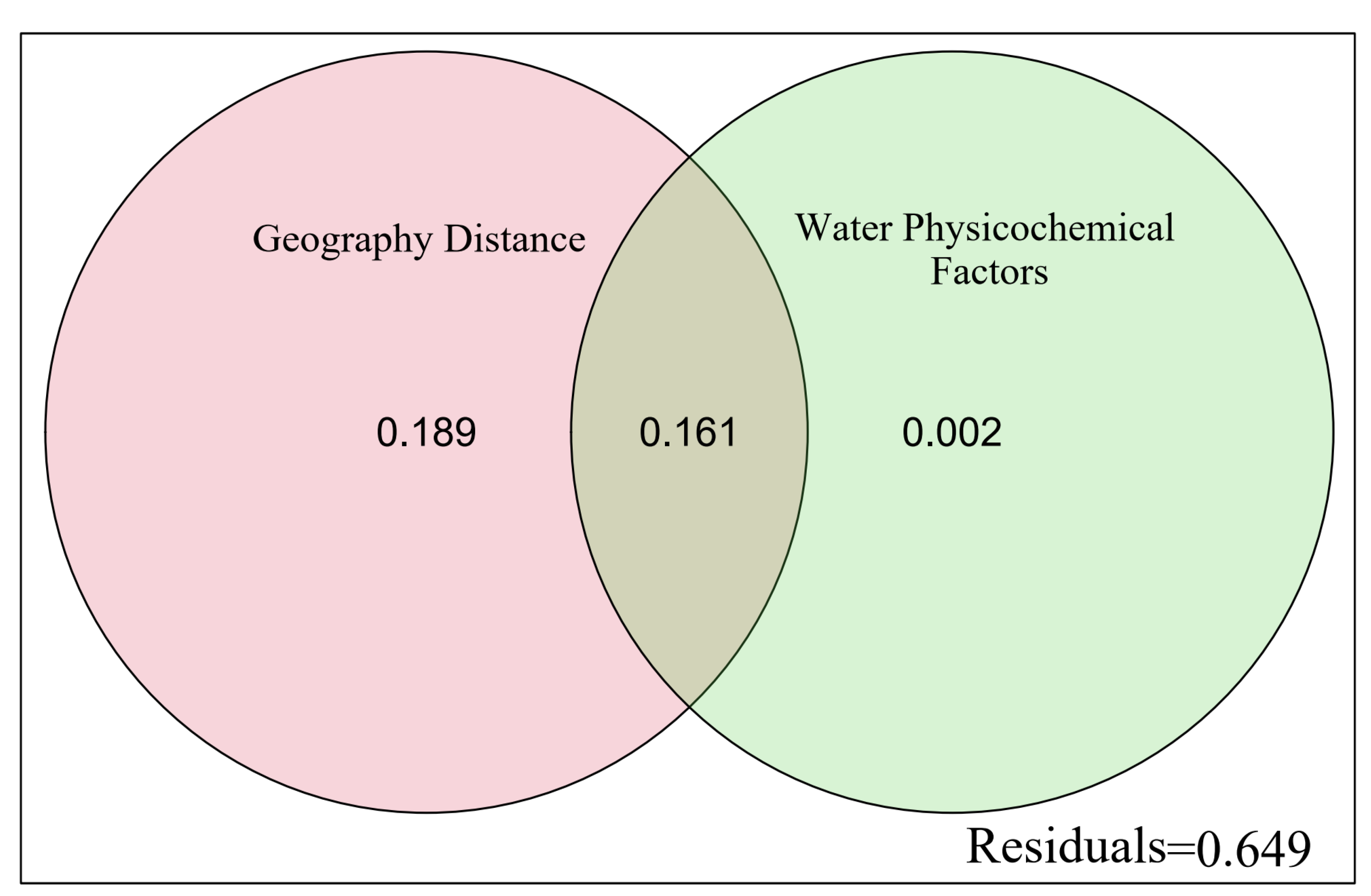
3.5. Influence of Physicochemical Factors
3.6. Contribution Difference in Different Environmental Factors
4. Discussion
4.1. Characteristics of Fish Diversity in Water-Transferred Reaches
4.2. Spatio-Temporal Pattern of Fish Communities
4.3. Dispersal Limitation Determines Spatial Heterogeneity of Fish Communities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wohl, E.E. Mountain Rivers; American Geophysical Union: Washington, DC, USA, 2013; Volume 14. [Google Scholar]
- Cuo, L.; Zhang, Y.; Zhu, F.; Liang, L. Characteristics and changes of streamflow on the Tibetan Plateau: A review. J. Hydrol. Reg. Stud. 2014, 2, 49–68. [Google Scholar] [CrossRef]
- Somers, L.D.; McKenzie, J.M. A review of groundwater in high mountain environments. Wires. Water 2020, 7, e1475. [Google Scholar] [CrossRef]
- Chen, Z.; Luo, L.; Wang, Z.; He, D.; Zhang, L. Diversity and distribution of fish in the Qilian Mountain Basin. Biodivers. Data J. 2022, 10, e85992. [Google Scholar] [CrossRef] [PubMed]
- Mao, K.S.; Wang, Y.; Liu, J.Q. Evolutionary origin of species diversity on the Qinghai-Tibet Plateau. J. Syst. Evol. 2021, 59, 1142–1158. [Google Scholar] [CrossRef]
- He, D.; Chen, Y. Phylogeography of Schizothorax o’connori (Cyprinidae: Schizothoracinae) in the Yarlung Tsangpo River, Tibet. Hydrobiologia 2009, 635, 251–262. [Google Scholar] [CrossRef]
- Wu, H.; Gu, Q.; Zhou, C.; Tang, Y.; Husemann, M.; Meng, X.; Zhang, J.; Nie, G.; Li, X. Molecular phylogeny and biogeography of Triplophysa stone loaches in the Central Chinese Mountains. Biol. J. Linn. Soc. 2020, 130, 563–577. [Google Scholar] [CrossRef]
- Huang, X.; Wang, X.; Zhang, X.; Zhou, C.; Ma, J.; Feng, X. Ecological risk assessment and identification of risk control priority areas based on degradation of ecosystem services: A case study in the Tibetan Plateau. Ecol. Indic. 2022, 141, 109078. [Google Scholar] [CrossRef]
- Wei, M.; Feng, T.; Lin, Y.; He, S.; Yan, H.; Qiao, R.; Chen, Q. Elevation-associated pathways mediate aquatic biodiversity at multi-trophic levels along a plateau inland river. Water Res. 2024, 258, 121779. [Google Scholar] [CrossRef]
- Zhang, Q.; Yuan, R.; Singh, V.P.; Xu, C.Y.; Fan, K.; Shen, Z.; Wang, G.; Zhao, J. Dynamic vulnerability of ecological systems to climate changes across the Qinghai-Tibet Plateau, China. Ecol. Indic. 2022, 134, 108483. [Google Scholar] [CrossRef]
- Zhao, G.; Mu, X.; Wen, Z.; Wang, F.; Gao, P. Soil erosion, conservation, and eco-environment changes in the Loess Plateau of China. Land Degrad. Dev. 2013, 24, 499–510. [Google Scholar] [CrossRef]
- Tian, K. Plateau Wetlands: Functions, Values, and Environmental Deterioration. In The China Environment Yearbook; Brill: Leiden, The Netherlands, 2010; Volume 4. [Google Scholar]
- Wang, H.; Zhou, X.; Wan, C.; Fu, H.; Zhang, F.; Ren, J. Eco-environmental degradation in the northeastern margin of the Qinghai-Tibetan Plateau and comprehensive ecological protection planning. Environ. Geol. 2008, 55, 1135–1147. [Google Scholar] [CrossRef]
- Giller, P. (Ed.) Community Structure and the Niche; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar]
- Tilman, D. Niche tradeoffs, neutrality, and community structure: A stochastic theory of resource competition, invasion, and community assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 10854–10861. [Google Scholar] [CrossRef]
- Chen, X.; Li, Z.; Boda, P.; Fernandes, I.M.; Xie, Z.; Zhang, E. Environmental filtering in the dry season and spatial structuring in the wet: Different fish community assembly rules revealed in a large subtropical floodplain lake. Environ. Sci. Pollut. Res. 2022, 29, 69875–69887. [Google Scholar] [CrossRef]
- Padial, A.A.; Ceschin, F.; Declerck, S.A.; De Meester, L.; Bonecker, C.C.; Lansac-Tôha, F.A.; Rodrigues, L.; Rodrigues, L.C.; Train, S.; Velho, L.F.M.; et al. Dispersal ability determines the role of environmental, spatial and temporal drivers of metacommunity structure. PLoS ONE 2014, 9, e111227. [Google Scholar] [CrossRef]
- Kirk, M.A.; Rahel, F.J.; Laughlin, D.C. Environmental filters of freshwater fish community assembly along elevation and latitudinal gradients. Global Ecol. Biogeogr. 2022, 31, 470–485. [Google Scholar] [CrossRef]
- Córdova-Tapia, F.; Hernández-Marroquín, V.; Zambrano, L. The role of environmental filtering in the functional structure of fish communities in tropical wetlands. Ecol. Freshw. Fish. 2018, 27, 522–532. [Google Scholar] [CrossRef]
- Le Bagousse-Pinguet, Y.; Gross, N.; Maestre, F.T.; Maire, V.; de Bello, F.; Fonseca, C.R.; Kattge, J.; Valencia, E.; Leps, J.; Liancourt, P. Testing the environmental filtering concept in global drylands. J. Ecol. 2017, 105, 1058–1069. [Google Scholar] [CrossRef]
- Li, Z.; Zhu, H.; García-Girón, J.; Gu, S.; Heino, J.; Xiong, X.; Yang, J.; Zhao, X.; Jia, Y.; Xie, Z.; et al. Historical and dispersal processes drive community assembly of multiple aquatic taxa in glacierized catchments in the Qinghai-Tibet plateau. Environ. Res. 2024, 251, 118746. [Google Scholar] [CrossRef]
- Lin, P.; Fujiwara, M.; Ma, B.; Xia, Z.; Wu, X.; Wang, C.; Chang, T.; Gao, X. Ecological drivers shaping mainstem and tributary fish communities in the upper Jinsha River, southeastern Qinghai-Tibet Plateau. Ecol. Process. 2025, 14, 8. [Google Scholar] [CrossRef]
- Kadye, W.T.; Magadza, C.H.; Moyo, N.A.; Kativu, S. Stream fish assemblages in relation to environmental factors on a montane plateau (Nyika Plateau, Malawi). Environ. Bio. Fish. 2008, 83, 417–428. [Google Scholar] [CrossRef]
- Harris, A.C.; Oyler-McCance, S.J.; Fike, J.A.; Fairchild, M.P.; Kennedy, C.M.; Crockett, H.J.; Winkelman, D.L.; Kanno, Y. Population genetics reveals bidirectional fish movement across the Continental Divide via an interbasin water transfer. Conserv. Genet. 2022, 23, 839–851. [Google Scholar] [CrossRef]
- Yevjevich, V. Water diversions and interbasin transfers. Water Int. 2001, 26, 342–348. [Google Scholar] [CrossRef]
- Sadeghi, S.H.R.; Kazemi Kia, S.; Kheirfam, H.; Hazbavi, Z. Experiences and consequences of inter-basin water transfer worldwide. Iran-Water Resour. Res. 2016, 12, 120–140. [Google Scholar]
- Zhuang, W. Eco-environmental impact of inter-basin water transfer projects: A review. Environ. Sci. Pollut. Res. 2016, 23, 12867–12879. [Google Scholar] [CrossRef]
- Snaddon, C.D.; Davies, B.R.; Wishart, M.J.; Meador, M.E.; Thoms, M. A Global Overview of Inter-Basin Water Transfer Schemes, with an Appraisal of Their Ecological, Socioeconomic and Socio-Political Implications, and Recommendations for Their Management; Water Research Commission Report No. TT120/00; Water Research Commission: Pretoria, South Africa, 1999. [Google Scholar]
- Yan, H.; Lin, Y.; Chen, Q.; Zhang, J.; He, S.; Feng, T.; Wang, Z.; Chen, C.; Ding, J. A review of the eco-environmental impacts of the South-to-North Water Diversion: Implications for interbasin water transfers. Engineering 2023, 30, 161–169. [Google Scholar] [CrossRef]
- Jing, L.; Li, F.; Cui, Q.; Cao, T.; Zhang, Q. Research on the upper-route water transfer scheme of the west route of south-to-north water Diversion project. Yellow River 2023, 45, 6–8. [Google Scholar]
- Liu, S.; Yang, Y.; Deng, B.; Zhong, Y.; Wen, L.; Sun, W.; Li, Z.; Jansa, L.; Song, J.; Zhang, X.; et al. Tectonic evolution of the Sichuan basin, southwest China. Earth Sci. Rev. 2021, 213, 103470. [Google Scholar] [CrossRef]
- Xiao, H.; Shao, H.; Long, J.; Zhang, S.; He, S.; Wang, D. Spatial-temporal pattern evolution and geological influence factors analysis of ecological vulnerability in Western Sichuan mountain region. Ecol. Indic. 2023, 155, 110980. [Google Scholar] [CrossRef]
- Ci, M.; Ye, L.; Liao, C.; Yao, L.; Tu, Z.; Xing, Q.; Tang, X.; Ding, Z. Long-term dynamics of ecosystem services and their influencing factors in ecologically fragile southwest China. Sustainability 2023, 15, 12331. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, H.; Lei, P.; Xin, X.; Zhang, A.; Yin, W. Evidence on the causes of the rising levels of CODMn along the middle route of the South-to-North Diversion Project in China: The role of algal dissolved organic matter. J. Environ. Sci. 2022, 113, 281–290. [Google Scholar] [CrossRef]
- Shen, L.; Dou, M.; Xia, R.; Li, G.; Yang, B. Effects of hydrological change on the risk of riverine algal blooms: Case study in the mid-downstream of the Han River in China. Environ. Sci. Pollut. Res. 2021, 28, 19851–19865. [Google Scholar] [CrossRef]
- Guo, C.; Chen, Y.; Liu, H.; Lu, Y.; Qu, X.; Yuan, H.; Lek, S.; Xie, S. Modelling fish communities in relation to water quality in the impounded lakes of China’s South-to-North Water Diversion Project. Ecol. Model. 2019, 397, 25–35. [Google Scholar] [CrossRef]
- Guo, C.; Chen, Y.; Gozlan, R.E.; Liu, H.; Lu, Y.; Qu, X.; Yuan, H.; Lek, S.; Wang, L. Patterns of fish communities and water quality in impounded lakes of China’s south-to-north water diversion project. Sci. Total Environ. 2020, 713, 136515. [Google Scholar] [CrossRef]
- Ning, Z.; Zhang, J.; Yuan, S.; Wang, G. Impact of climate change on water resources in the western route areas of the South-to-North water diversion project. Atmosphere 2022, 13, 799. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, D.; Wang, W.; Yu, S.; Wang, H. Quantifying future water and energy security in the source area of the western route of China’s South-to-North water diversion project within the context of climatic and societal changes. J. Hydrol. Reg. Stud. 2023, 47, 101443. [Google Scholar] [CrossRef]
- Fan, D.; Zeng, S.; Du, H.; Ren, Y.; Xia, J. Projected flow regimes and biodiversity changes under climate change in the planning western route source areas of the South-to-North Water Diversion Project. Ecol. Indic. 2023, 154, 110827. [Google Scholar] [CrossRef]
- Jargal, N.; Kim, J.E.; An, K.G. An extended novel approach of river health analysis using nonmetric scaling ordination of fish ecological entities and functional identity indices. J. Environ. Manag. 2024, 365, 121582. [Google Scholar] [CrossRef]
- Bacigalupi, J.; Staples, D.F.; Treml, M.T.; Bahr, D.L. Development of fish-based indices of biological integrity for Minnesota lakes. Ecol. Indic. 2021, 125, 107512. [Google Scholar] [CrossRef]
- Wu, J.; Wang, H.; Huo, L.; Du, H.; Wang, C.; Wei, Q. Resource and distribution of two fishes in the Make River. Freshw. Fish. 2015, 5, 46–49. [Google Scholar]
- Hart, R.K.; Calver, M.C.; Dickman, C.R. The index of relative importance: An alternative approach to reducing bias in descriptive studies of animal diets. Wildlife Res. 2002, 29, 415–421. [Google Scholar] [CrossRef]
- Anderson, M.J.; Walsh, D.C. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Hirst, C.N.; Jackson, D.A. Reconstructing community relationships: The impact of sampling error, ordination approach, and gradient length. Divers. Distrib. 2007, 13, 361–371. [Google Scholar] [CrossRef]
- Ter Braak, C.J.; Verdonschot, P.F. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca. hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Li, H.; Tan, L.; Li, X.; Cai, Q. Aquatic protected area system in the Qinghai-Tibet Plateau: Establishment, challenges and prospects. Front. Ecol. Evol. 2024, 12, 1204494. [Google Scholar] [CrossRef]
- Jia, Y.; Kennard, M.J.; Liu, Y.; Sui, X.; Li, K.; Wang, G.; Chen, Y. Human disturbance and long-term changes in fish taxonomic, functional and phylogenetic diversity in the Yellow River, China. Hydrobiologia 2020, 847, 3711–3725. [Google Scholar] [CrossRef]
- He, D.; Chen, J.; Ding, L.; Xu, Y.; Huang, J.; Sui, X. The status and distribution pattern of fish diversity in the Yarlung Tsangpo River. Biodivers. Sci. 2024, 32, 24143. [Google Scholar]
- Li, W.; Guo, W.; Liu, H.; Qiao, Q.; Zhao, K.; Ai, D.; Liu, C.; Zhao, W. Spawning grounds of Schizothoracinae fish on the Tibetan Plateau. Environ. Biol. Fish. 2024, 107, 1–14. [Google Scholar] [CrossRef]
- Zhang, Z.; Liang, W.; Zhang, D.; Zhou, W.; Yang, Y.; Cheng, R.; Xiong, W.; Chang, X.; Cheng, F. Fish resources and diversity in the mainstem and tributaries of Dadu River from Jinchuan to Danba. J. Hydroecol. 2023, 44, 54–61. [Google Scholar]
- Jia, Y.; Sui, X.; Chen, Y.; He, D. Climate change and spatial distribution shaped the life-history traits of schizothoracine fishes on the Tibetan Plateau and its adjacent areas. Glob. Ecol. Conserv. 2020, 22, e01041. [Google Scholar] [CrossRef]
- Gong, Z.; Zhai, D.; Chen, J.; Liu, B.; Zhu, T. Landscape determinants of genetic structure for Schizopygopsis younghusbandi in the Yarlung Tsangpo River drainage, Tibetan Plateau. Ecol. Indic. 2023, 151, 110309. [Google Scholar] [CrossRef]
- Liang, Y.; He, D.; Jia, Y.; Sun, H.; Chen, Y. Phylogeographic studies of schizothoracine fishes on the central Qinghai-Tibet Plateau reveal the highest known glacial microrefugia. Sci. Rep. 2017, 7, 10983. [Google Scholar] [CrossRef]
- He, Z.; Gao, K.; Chen, H.; Yang, D.; Pu, Y.; Zheng, L.; Jiao, Y.; Xiong, J.; Chen, Q.; Lai, B.; et al. Comparative population dynamics of Schizothorax wangchiachii (Cyprinidae: Schizothoracinae) in the middle reaches of the Yalong River and the upper reaches of the Jinsha River, China. Animals 2023, 13, 2209. [Google Scholar] [CrossRef]
- Rui, H.; Li, Y.; Sheng, Z.; Zhang, Y.; Liu, X.; Wu, X. Distribution and habitat character of Hucho bleekeri in the Dadu river basin. Resour. Environ. Yangtze Basin 2015, 24, 1779–1785. [Google Scholar]
- Chen, J.; Zhu, X.; Yang, X.; Hu, X.; Lin, P.; Xu, B.; Wei, K.; Ma, B. Age, growth, and reproductive biology of Euchiloglanis davidi in the middle and lower Yalong River, China. Fishes 2023, 8, 435. [Google Scholar] [CrossRef]
- Liu, B. Recent advances in altitudinal distribution patterns of biodiversity. Ecol. Environ. 2021, 30, 438. [Google Scholar]
- Jaramillo-Villa, U.; Maldonado-Ocampo, J.A.; Escobar, F. Altitudinal variation in fish assemblage diversity in streams of the central Andes of Colombia. J. Fish Biol. 2010, 76, 2401–2417. [Google Scholar] [CrossRef]
- Bhatt, J.P.; Manish, K.; Pandit, M.K. Elevational gradients in fish diversity in the Himalaya: Water discharge is the key driver of distribution patterns. PLoS ONE 2012, 8, e46237. [Google Scholar] [CrossRef]
- McPolin, M.C.; Kranabetter, J.M. Influence of endemic versus cosmopolitan species on the local assembly of ectomycorrhizal fungal communities. New Phytol. 2021, 229, p2395. [Google Scholar] [CrossRef]
- Burlakova, L.E.; Karatayev, A.Y.; Karatayev, V.A.; May, M.E.; Bennett, D.L.; Cook, M.J. Endemic species: Contribution to community uniqueness, effect of habitat alteration, and conservation priorities. Biol. Conserv. 2011, 144, 155–165. [Google Scholar] [CrossRef]
- Li, X.; Xu, Q.; Xia, R.; Zhang, N.; Wang, S.; Ding, S.; Gao, X.; Jia, X.; Shang, G.; Chen, X. Stochastic process is main factor to affect plateau river fish community assembly. Environ. Res. 2024, 254, 119083. [Google Scholar] [CrossRef]
- Du, T.; Ding, C.; Yang, K.; Chen, J.; Liu, X.; Lv, W.; Ding, L.; He, D.; Tao, J. A global dataset on species occurrences and functional traits of Schizothoracinae fish. Sci. Data 2024, 11, 272. [Google Scholar] [CrossRef]
- Chi, W.; Ma, X.; Niu, J.; Zou, M. Genome-wide identification of genes probably relevant to the adaptation of schizothoracins (Teleostei: Cypriniformes) to the uplift of the Qinghai-Tibet Plateau. BMC Genomics 2017, 18, 1–8. [Google Scholar] [CrossRef]
- Wang, X.; Gan, X.; Li, J.; Chen, Y.; He, S. Cyprininae phylogeny revealed independent origins of the Tibetan Plateau endemic polyploid cyprinids and their diversifications related to the Neogene uplift of the plateau. Sci. China Life Sci. 2016, 59, 1149–1165. [Google Scholar] [CrossRef]
- Guan, L.; Chi, W.; Xiao, W.; Chen, L.; He, S. Analysis of hypoxia-inducible factor alpha polyploidization reveals adaptation to Tibetan plateau in the evolution of schizothoracine fish. BMC Evol. Biol. 2014, 14, 1–13. [Google Scholar] [CrossRef]
- Smith, G.R.; Badgley, C.; Eiting, T.P.; Larson, P.S. Species diversity gradients in relation to geological history in North American freshwater fishes. Evol. Ecol. Res. 2010, 12, 693–726. [Google Scholar]
- Chakona, A.; Swartz, E.R.; Gouws, G. Evolutionary drivers of diversification and distribution of a southern temperate stream fish assemblage: Testing the role of historical isolation and spatial range expansion. PLoS ONE 2013, 8, e70953. [Google Scholar] [CrossRef]
- Albert, J.S.; Carvalho, T.P.; Petry, P.; Holder, M.A.; Maxime, E.L.; Espino, J.; Corahua, I.; Quispe, R.; Rengifo, B.; Ortega, H.; et al. Aquatic biodiversity in the Amazon: Habitat specialization and geographic isolation promote species richness. Animals 2011, 1, 205–241. [Google Scholar] [CrossRef]
- Liu, F.; Li, M.; Wang, J.; Gong, Z.; Liu, M.; Liu, H.; Lin, P. Species composition and longitudinal patterns of fish assemblages in the middle and lower Yarlung Zangbo River, Tibetan Plateau, China. Ecol. Indic. 2021, 125, 107542. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Kühn, I.; Wild, J.; Arianoutsou, M.; Bacher, S.; Chiron, F.; Didžiulis, V.; Essl, F.; et al. Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc. Natl. Acad. Sci. USA 2010, 107, 12157–12162. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Season | Dadu River Basin | Yalong River Basin | |||||
|---|---|---|---|---|---|---|---|---|
| SDR | MR | AR | DUR | YR | NR | DAR | ||
| Channel width (m) | Spring | 98.33 | 48.44 | 32.09 | 49.10 | 120.79 | 53.12 | 50.58 |
| Autumn | 112.54 | 34.50 | 43.31 | 53.84 | 117.05 | 56.21 | 48.48 | |
| Water temperature (°C) | Spring | 11.08 | 10.58 | 8.49 | 8.04 | 10.92 | 10.08 | 10.03 |
| Autumn | 15.39 | 10.86 | 9.49 | 11.13 | 11.70 | 12.17 | 11.74 | |
| pH | Spring | 8.22 | 8.24 | 7.88 | 8.22 | 8.13 | 8.15 | 8.17 |
| Autumn | 8.17 | 5.45 | 6.78 | 7.27 | 8.09 | 8.55 | 8.21 | |
| Flow velocity (m/s) | Spring | 0.53 | 0.20 | 0.33 | 0.47 | 0.54 | 0.49 | 0.39 |
| Autumn | 0.52 | 0.68 | 0.52 | 0.86 | 0.85 | 0.75 | 0.91 | |
| Conductivity (μS/cm) | Spring | 164.86 | 173.66 | 100.49 | 204.20 | 230.82 | 256.20 | 193.31 |
| Autumn | 223.93 | 202.90 | 103.98 | 223.22 | 330.95 | 325.05 | 214.71 | |
| Average altitude (m) | 2779 | 3473 | 3523 | 3621 | 3754 | 3862 | 3974 | |
| Family | Genus | Latin Name | SDR | MR | AR | DUR | YR | NR | DAR |
|---|---|---|---|---|---|---|---|---|---|
| Cyprinidae | Aristichthys | A. nobilis * | + | ||||||
| Carassius | C. auratus * | + | |||||||
| Cyprinus | C. carpio * | + | |||||||
| C. carpio var. * | + | ||||||||
| Gymnocypris | G. eckloni * | + | + | ||||||
| Gymnodiptychus | G. pachycheilus | + | + | + | |||||
| Ptychobarbus | P. leptosomus | + | + | + | + | ||||
| Schizopygopsis | S. chengi | + | + | + | + | ||||
| S. malacanthus | + | + | + | ||||||
| Schizothorax | S. davidi | + | + | + | |||||
| S. dolichonema | + | ||||||||
| S. kozlovi | + | ||||||||
| S. prenanti | + | + | |||||||
| S. wangchiachii | + | + | |||||||
| Nemacheilidae | Triplophysa | T. markehenensis | + | + | |||||
| T. orientalis | + | + | |||||||
| T. pseudoscleroptera | + | + | + | + | + | ||||
| T. pseudostenura | + | + | + | ||||||
| T. stoliczkae | + | + | + | ||||||
| Cobitidae | Paramisgurnus | P. dabryanus * | + | + | |||||
| Misgurnus | M. anguillicaudatus * | + | |||||||
| Siluridae | Silurus | S. asotus * | + |
| Species | N% | W% | F% | IRI |
|---|---|---|---|---|
| S. chengi | 46.8% | 18.2% | 59.4% | 3858 |
| S. malacanthus | 30.3% | 44.1% | 40.6% | 3025 |
| G. pachycheilus | 4.3% | 12.6% | 40.6% | 683 |
| T. pseudoscleroptera | 5.8% | 1.8% | 53.1% | 404 |
| T. pseudostenura | 2.9% | 0.5% | 37.5% | 127 |
| S. prenanti | 2.8% | 0.6% | 28.1% | 95 |
| G. eckloni * | 1.6% | 1.6% | 18.8% | 60 |
| T. stoliczkae | 1.3% | 0.3% | 34.4% | 56 |
| P. leptosomus | 0.6% | 1.6% | 21.9% | 50 |
| C. carpio * | 0.1% | 5.7% | 6.3% | 36 |
| S. dolichonema | 0.1% | 5.0% | 6.3% | 32 |
| S. davidi | 0.7% | 0.3% | 21.9% | 22 |
| P. dabryanus * | 0.5% | 0.4% | 15.6% | 14 |
| A. nobilis * | 0.1% | 3.6% | 3.1% | 11 |
| S. asotus * | 0.3% | 1.5% | 6.3% | 11 |
| T. orientalis | 0.7% | 0.2% | 9.4% | 8 |
| S. wangchiachii | 0.3% | 0.6% | 6.3% | 5 |
| C. carpio var. * | 0.0% | 1.4% | 3.1% | 4 |
| S. kozlovi | 0.2% | 0.0% | 6.3% | 1 |
| C. auratus * | 0.2% | 0.1% | 3.1% | 1 |
| M. anguillicaudatus * | 0.2% | 0.0% | 3.1% | 1 |
| T. markehenensis | 0.1% | 0.0% | 6.3% | 1 |
| Region | Sum of Squares | R2 | F | Pr (>F) | |
|---|---|---|---|---|---|
| Overall | Season | 0.459 | 0.026 | 1.509 | 0.123 |
| Reaches | 10.793 | 0.621 | 11.821 | 0.001 * | |
| Season and Reaches | 0.651 | 0.037 | 0.856 | 0.679 | |
| Residual | 5.478 | 0.315 | |||
| Sum | 17.382 | 1.000 | |||
| Reaches | AR vs. DUR | 1.499 | 0.473 | 11.677 | 0.002 * |
| AR vs. MR | 0.121 | 0.094 | 0.624 | 0.590 | |
| AR vs. DAR | 2.145 | 0.527 | 13.396 | 0.001 * | |
| AR vs. NR | 2.130 | 0.495 | 11.765 | 0.001 * | |
| AR vs. YR | 2.333 | 0.589 | 17.201 | 0.001 * | |
| AR vs. SDR | 0.983 | 0.254 | 4.437 | 0.001 * | |
| DUR vs. MR | 0.522 | 0.369 | 5.258 | 0.021 * | |
| DUR vs. DAR | 3.300 | 0.666 | 29.948 | 0.001 * | |
| DUR vs. NR | 3.155 | 0.624 | 24.852 | 0.001 * | |
| DUR vs. YR | 3.472 | 0.719 | 38.303 | 0.001 * | |
| DUR vs. SDR | 2.017 | 0.436 | 12.355 | 0.001 * | |
| MR vs. DAR | 1.255 | 0.523 | 8.767 | 0.025 * | |
| MR vs. NR | 1.205 | 0.463 | 6.902 | 0.022 * | |
| MR vs. YR | 1.314 | 0.607 | 12.338 | 0.018 * | |
| MR vs. SDR | 0.566 | 0.212 | 2.421 | 0.055 | |
| DAR vs. NR | 0.293 | 0.120 | 1.905 | 0.132 | |
| DAR vs. YR | 0.302 | 0.158 | 2.619 | 0.054 | |
| DAR vs. SDR | 2.829 | 0.497 | 14.814 | 0.001 * | |
| NR vs. YR | 0.116 | 0.059 | 0.874 | 0.413 | |
| NR vs. SDR | 2.609 | 0.456 | 12.563 | 0.001 * | |
| YR vs. SDR | 2.962 | 0.535 | 17.281 | 0.002 * | |
| Reaches | Bray–Curtis Dissimilarity Index | Geographical Distance (Km) | ||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| DAR vs. MR | 1.00 | 0.00 | 148.11 | 16.80 |
| NR vs. MR | 1.00 | 0.00 | 127.96 | 12.07 |
| YR vs. MR | 1.00 | 0.00 | 191.93 | 22.98 |
| SDR vs. DAR | 1.00 | 0.00 | 198.49 | 33.39 |
| SDR vs. YR | 1.00 | 0.01 | 248.52 | 29.75 |
| YR vs. DUR | 0.99 | 0.02 | 147.31 | 26.45 |
| SDR vs. NR | 0.99 | 0.02 | 169.47 | 33.68 |
| DAR vs. DUR | 0.99 | 0.03 | 102.82 | 23.24 |
| NR vs. DUR | 0.99 | 0.02 | 83.30 | 20.75 |
| NR vs. AR | 0.96 | 0.06 | 168.91 | 10.39 |
| YR vs. AR | 0.96 | 0.05 | 231.37 | 19.95 |
| DAR vs. AR | 0.95 | 0.07 | 188.89 | 13.00 |
| SDR vs. DUR | 0.84 | 0.17 | 117.46 | 32.31 |
| DUR vs. AR | 0.80 | 0.15 | 109.00 | 12.58 |
| SDR vs. AR | 0.79 | 0.16 | 130.67 | 28.05 |
| SDR vs. MR | 0.79 | 0.19 | 115.08 | 22.16 |
| MR vs. DUR | 0.66 | 0.23 | 67.98 | 14.74 |
| NR vs. DAR | 0.55 | 0.20 | 59.77 | 33.58 |
| MR vs. AR | 0.53 | 0.24 | 50.12 | 9.39 |
| YR vs. DAR | 0.49 | 0.18 | 65.09 | 28.98 |
| YR vs. NR | 0.48 | 0.18 | 93.42 | 35.20 |
| Variables | Relative Importance (%) | Significance |
|---|---|---|
| Altitude | 50.1 | 0.001 * |
| Conductivity | 19.2 | 0.004 * |
| Channel width | 14.1 | 0.008 * |
| Water temperature | 9.5 | 0.025 * |
| Flow velocity | 5.3 | 0.1249 |
| pH | 1.8 | 0.3506 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, T.; Gong, Z.; Shang, K.; Hu, P. Dispersal Limitation Dominates Riverine Fish Communities in the Areas of the Water Diversion Project in the Western Sichuan Plateau, China. Animals 2025, 15, 730. https://doi.org/10.3390/ani15050730
Chang T, Gong Z, Shang K, Hu P. Dispersal Limitation Dominates Riverine Fish Communities in the Areas of the Water Diversion Project in the Western Sichuan Plateau, China. Animals. 2025; 15(5):730. https://doi.org/10.3390/ani15050730
Chicago/Turabian StyleChang, Tao, Zheng Gong, Kunyu Shang, and Piao Hu. 2025. "Dispersal Limitation Dominates Riverine Fish Communities in the Areas of the Water Diversion Project in the Western Sichuan Plateau, China" Animals 15, no. 5: 730. https://doi.org/10.3390/ani15050730
APA StyleChang, T., Gong, Z., Shang, K., & Hu, P. (2025). Dispersal Limitation Dominates Riverine Fish Communities in the Areas of the Water Diversion Project in the Western Sichuan Plateau, China. Animals, 15(5), 730. https://doi.org/10.3390/ani15050730